
Abstract
The immune-correlates analysis of the RV144 trial suggested that epitopes targeted by protective antibodies (Abs) reside in the V1V2 domain of gp120. We mapped V1V2 positional sequence variation onto the conserved V1V2 structural fold and showed that while most of the solvent-accessible V1V2 amino acids vary between strains, there are two accessible molecular surface regions that are conserved and also naturally antigenic. These sites may contain epitopes targeted by broadly cross-reactive anti-V1V2 antibodies.
T
In contrast to the V3 loop, V1V2 demonstrates substantial length variability. Analysis of the distribution of insertions and deletions in V1V2 (Supplementary Methods; Supplementary Data are available online at
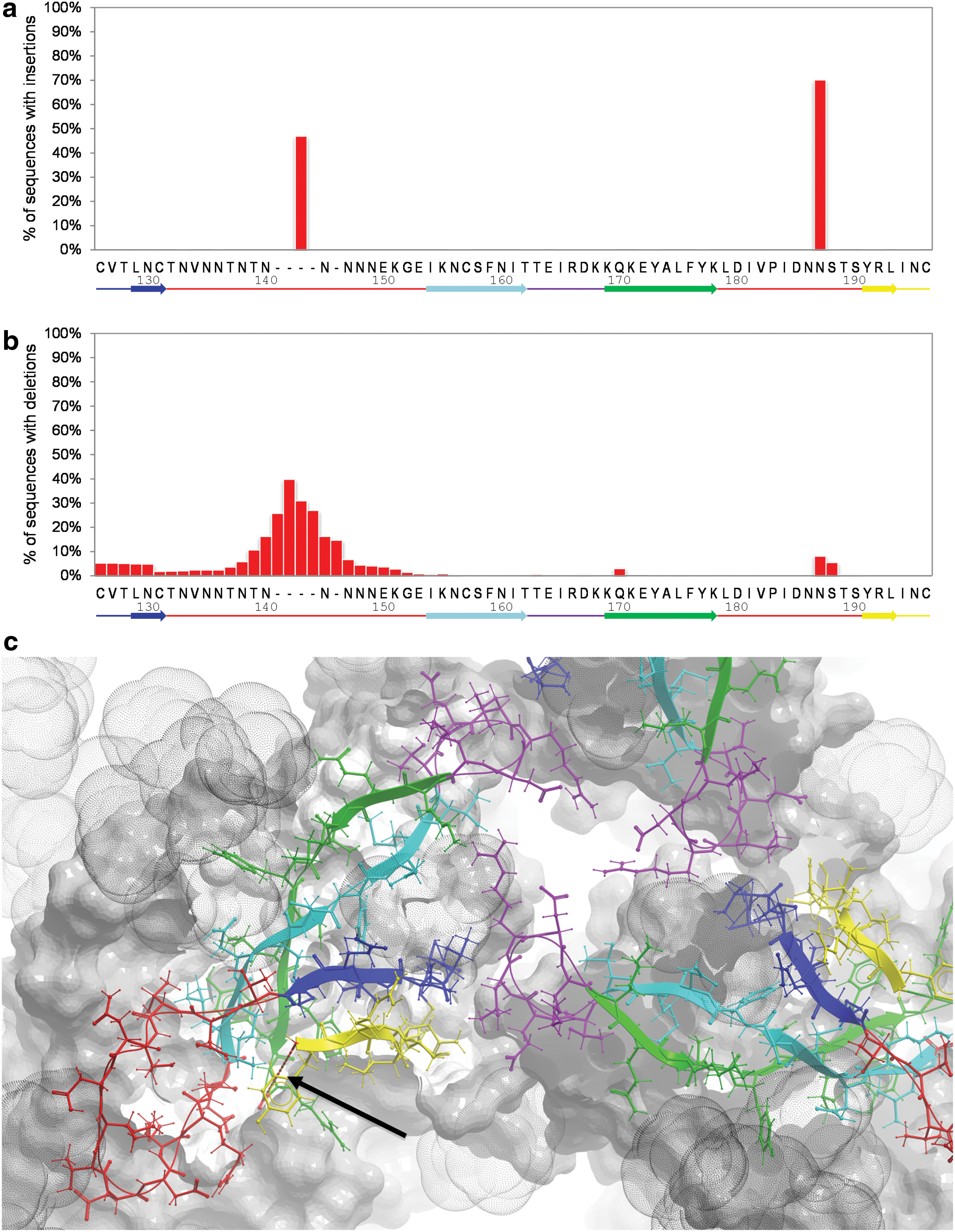
Occurrence of insertions and deletions in the V1V2 domain. Distributions of insertions
Accordingly, we calculated the side chain variability at each position of the V1V2 (Fig. 2; Supplementary Methods). The majority (29 out of 31) of V1V2 positions with variability scores higher than 50% are clustered to three linear segments: V1133–152, V2169–172, and V2185–190, which are likely subject to immune pressure. Indeed, RV144-associated mAbs CH58/CH596 as well as the BNAbs PG9/PG168 target the V2169–172 variable site (“C” ß-strand of V1V2). Narrowly cross-reactive CH58/CH59 mAbs engage variable amino acid side chains in this segment. In contrast, PG9/PG16 make sequence-independent contacts with the peptide backbone and a glycan, demonstrating, therefore, much broader cross-reactivity. CAP256 antibodies, while being broadly neutralizing for subtypes A and C, surprisingly target the highly variable lysine at position 169. 11,17 This could be explained by the fact that K169 is conserved in subtype C and K/R169 are conserved in subtype A (lysine and arginine have similar structural properties), but not in other subtypes.
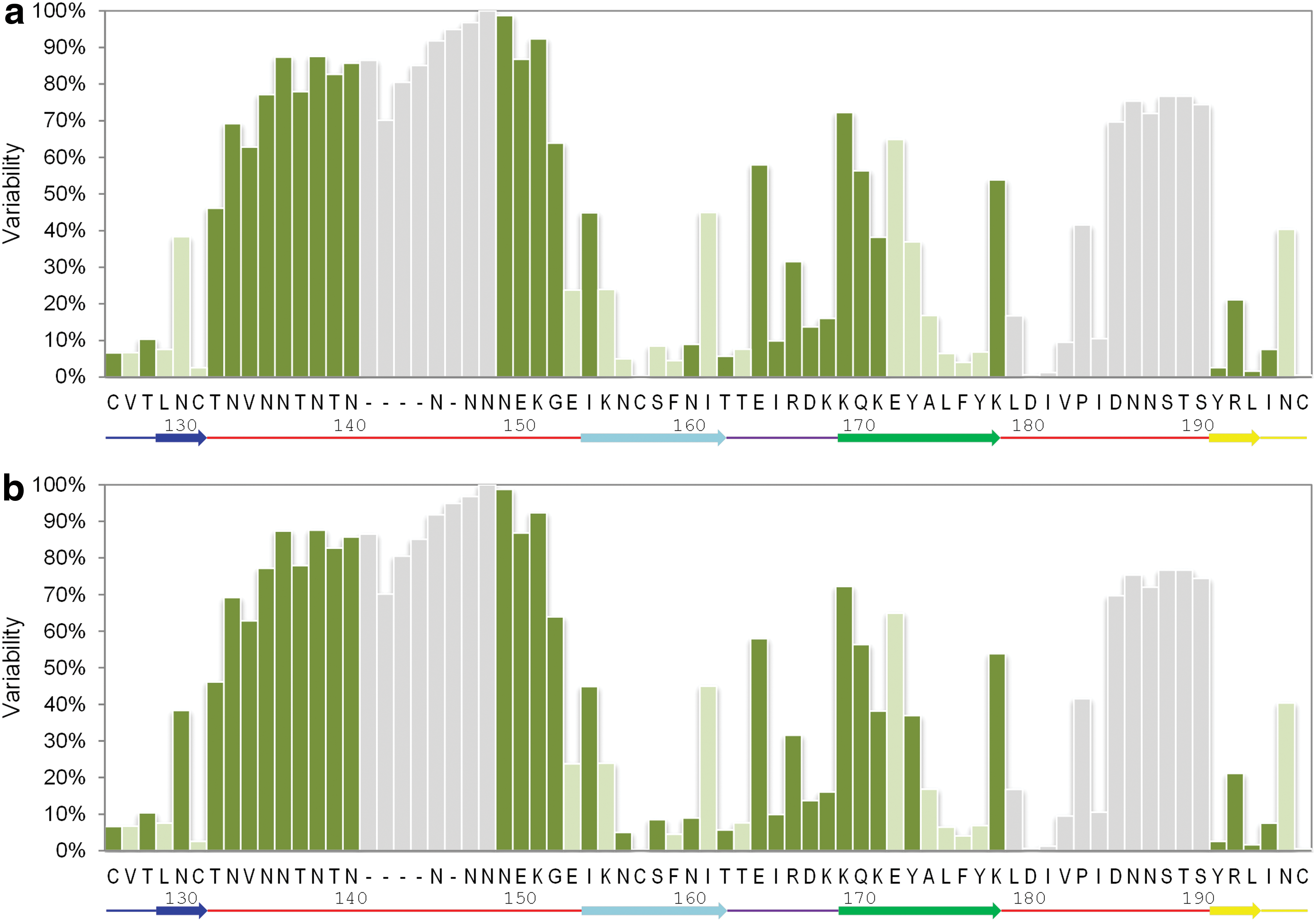
Position-specific variability and accessibility in the V1V2 domain. HIV-1 reference strain HxB2 positions from 126 to 196 are shown. The most frequently occurring side chains at each position corresponding to the HxB2 numbering are labeled on the x-axis. Each bar is colored according to the mean relative accessibility of that specific side chain in the context of glycosylated
Since these sequence variable regions are targeted by known Abs, they are expected to be solvent exposed. To test this, we calculated the solvent accessible area of each V1V2 amino acid in the context of the gp140 trimer (see Supplementary Methods). Since glycosylation could differ substantially between strains, we studied the accessibility of V1V2 both in a fully glycosylated trimer and in a trimer with removed glycans. As expected, the sequence-variable segments are exposed, while many of the sequence-conserved segments are poorly accessible (Fig. 2). The sequence-variable C-strand is the most accessible area of V1V2.
Interestingly, there are two relatively conserved and yet solvent-exposed V1V2 regions (Fig. 3). These regions are primarily composed of solvent-exposed amino acids (relative solvent accessibility of at least 20%; see Supplementary Methods) with variability scores of less than 50%, which are clustered together in a three-dimensional space. Region “I” is an intermonomer quaternary site formed by the tip of the V2 loop between its B and C strands (V2162–168) and stems of V1V2 on the A and B strands (V1127–128, V2192–194) of an adjacent monomer. This region is located at the apex of the trimer relatively far (∼15 Å) from the length-polymorphic V1V2 segments, and thus is likely accessible in many different strains. Region “II” is formed by positions located in three-dimensional proximity to the integrin binding site (V2176,179–184,191–194). This region can potentially be occluded by the two sequence-variable and structurally polymorphic loops in V1V2.
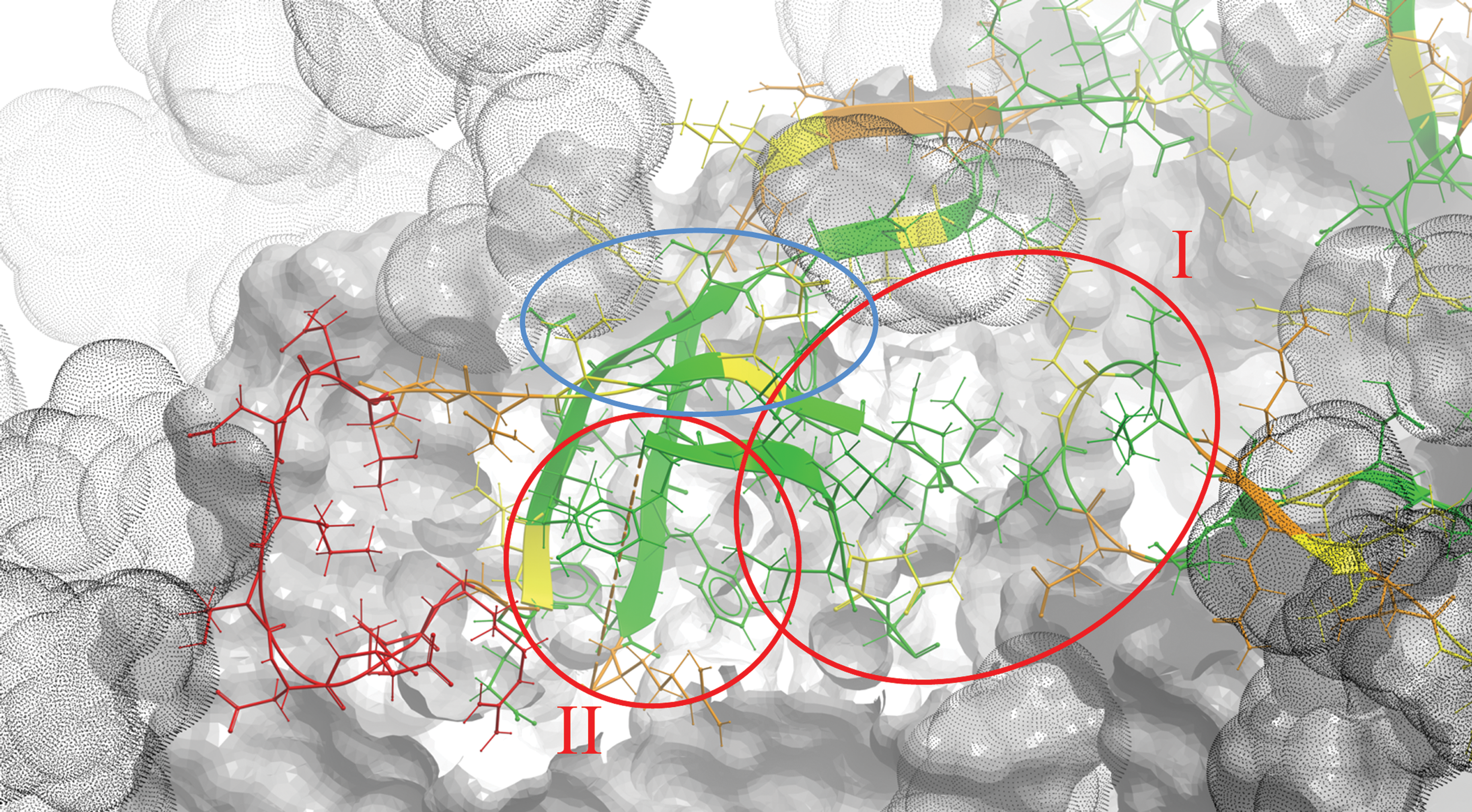
Positional variability in V1V2 mapped onto the three-dimensional structure of the gp140 trimer. The structure of V1V2 (positions 126–196) is shown in ribbon and stick representation, while the rest of the trimer is shown in skin representation. Each V1V2 side chain is colored from green for sequence-conserved to red for sequence-variable positions. The space occupied by glycans is shown by gray dot-envelopes. The two epitope regions discussed in the text are highlighted with red circles and labeled as “I” and “II.” Conserved side chains that can contribute to the formation of region “I” in less glycosylated strains are highlighted with a blue circle. Color images available online at
To test this, we performed computational sampling of one of these loops—V1132–153 (see Supplementary Methods). Indeed, the conformational flexibility of V1 is high enough to partly mask epitopes in region “II” (Fig. 4). The length-polymorphic loop of V2 is expected to be at the same three-dimensional location (although not resolved in crystal structures), and likely contributes further to this masking. Interestingly, if all glycans are removed some additional conserved side chains become accessible (Supplementary Fig. S1). Some of them (V1V2130,156,158,160) may further contribute to the formation of region “I” (Fig. 3). It is also important to mention that both regions “I” and “II” are conserved throughout subtypes A, B, C, and CRFs 01_AE and 02_AG (Supplementary Fig. S2; other clades were not studied due to a small sample size). This observation further emphasizes the potentially broadly cross-reactive nature of Abs targeting these two regions.

Conformational variability of the V1 loop in the context of the gp140 trimer. The structure of V1V2 (positions 126–196) is shown in ribbon and stick representation, while the rest of the trimer is shown in skin representation. Each V1V2 side chain is colored from green for sequence-conserved to red for sequence-variable positions. The space occupied by glycans is shown by gray dot-envelopes. The space that can reasonably be occupied by energetically favorable conformational flexibility of the crown segment of the V1 loop and the attached N137-linked glycan is shown as an orange cloud. Color images available online at
A linear fragment of region “I” (V2165–168) has been previously confirmed as antigenic in a murine model. 7 CAP256 antibodies also depend on conserved positions at that same region. 11,17 On the other hand, seven human conformational anti-V2 mAb have been recently mapped to V1V2 positions located within region “II.” 18 Normally, it would be reasonable to expect high sequence diversity in exposed and naturally antigenic viral regions due to the constant immune pressure. However, both regions “I” and “II” are conserved. Therefore, these V1V2 regions could be sites of vulnerability in HIV.
If immune pressure so evidently leads to sequence and length variation in most of the Ab-exposed regions of V1V2, then why might regions “I” and “II” be conserved despite their exposure? The most obvious reason would be that alterations in these regions decrease viral fitness. Region “I” offers a ready explanation for this: sequence variation may perturb an intermonomer contact important for trimer integrity. Reasons for the conservation of region “II” are not obvious, but predicted partial masking of “II” by the length-variable loops may be one of them.
In summary, our map of sequence variability outlines two conserved, unmasked, and apparently antigenic V1V2 segments that could be sites of vulnerability to human host Abs. Region “I” is an especially promising site in being quaternary, unmasked, and sequence conserved. The map also shows that the C-strand of V1V2 is exposed, highly variable, and perhaps the most immunogenic region of V1V2. However, to be broadly cross-reactive, Abs targeting this region have to bind the structurally conserved peptide backbone atoms, some conserved side chains, and/or glycans.
If the protective anti-V1V2 Abs detected in the RV144 trial that cross-react with multiple subtypes 3 are clonal species, they may target one or both sites outlined here. Our delineation of the three-dimensional structures of these sites can be useful for identifying the Abs targeting these regions and for the rational design of antigens capable of eliciting broadly cross-reactive anti-V1V2 Abs.
Footnotes
Acknowledgments
This work was supported by a grant from the National Institutes of Health, R01AI084119, to T.C.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.