
Abstract
Investigations into apoptotic pathways, intrinsic and extrinsic, and the effects of highly active antiretroviral therapy (HAART) on T cell death via those pathways may provide insight into the mechanisms of and barriers to immune recovery. HIV-1-infected patients were enrolled into a randomized, controlled study of the immune effects of a lopinavir/ritonavir (LPV/r)-based versus an efavirenz (EFV)-based HAART regimen in antiretroviral-naive subjects with CD4+ counts <350 cells/mm3. Patients were randomized to receive TDF/FTC/EFZ or TDF/FTC plus LPV/r. Fourteen patients were enrolled and 10 patients completed 6 months of therapy as per the protocol. CD4+ counts were measured before and during HAART therapy. We isolated T cell subsets to measure ex vivo apoptosis by propidium iodide staining. We also assessed caspase activation for the intrinsic and extrinsic pathways of apoptosis, as well as effector caspase activation. We also measured mitochondrial membrane potential. Cells were analyzed by flow cytometry. All patients had increased activation of caspase 8 (extrinsic pathway), caspase 9 (intrinsic pathway), effector caspases 3/7, and low mitochondrial membrane potential at baseline compared to controls. By 4 weeks, there was a decrease in activation of all caspases, but little further decrease by week 24. T cell mitochondrial membrane potential did not increase until week 12, but continued to increase until week 24. The only predictor of CD4+ count increase was the increase in mitochondrial membrane potential of naive cells at 6 months (r=0.66, p=0.038). This suggests that positive selection of naive CD4+ T cells in the thymus is the major determinant of CD4+ recovery.
T
Apoptosis is a major mechanism of T cell depletion in HIV infection, and this is reduced with effective antiretroviral therapy. 13,14 Accelerated CD4+ T cell apoptosis persists in immune nonresponders on HAART, even in patients with complete viral suppression. 15 Apoptosis occurs via “extrinsic” and “intrinsic pathways. 16 Extrinsic or receptor-mediated apoptosis results in activation of caspase 8. Intrinsic apoptosis results from the loss of mitochondrial membrane potential with subsequent cytochrome c release, and formation of an apoptosome that activates caspase 9.
Mitochondrial membrane potential is determined by the expression of antiapoptotic Bcl-2 proteins, and in the case of T cells, this is part of the process of positive selection in the thymus. Both the intrinsic and extrinsic pathways result in activation of effector caspases (caspases 3 and 7) that are involved in the cleavage of substrates involved in the process of apoptosis downstream in the cascade. Little is known about the pathways of apoptosis in patients with persistently accelerated CD4+ T cell death and poor immune recovery despite effective viral suppression with HAART. Investigation into the pathways of CD4+ T cell apoptosis, intrinsic and extrinsic, and the effects of HAART on these pathways may provide insight into the mechanisms of immune recovery.
There is substantial evidence that protease inhibitors may have a beneficial immune effect independent of their effect on viral replication. 17 –21 Our group has reported that a switch to a lopinavir/ritonavir (LPV/r)-containing regimen for patients with poor immune reconstitution despite complete viral suppression with HAART resulted in a greater increase in absolute CD4+ T cell counts compared to continuation of the current regimen, and this was associated with a reduction in ex vivo apoptosis of naive CD4+ T cells. 22 We conducted a clinical trial to determine if a boosted protease inhibitor (PI; LPV/r)-containing regimen had beneficial immune effects compared to a nonnucleoside reverse transcriptase inhibitor [NNRTI; efavirenz (EFV)]-based regimen in antiretroviral-naive patients with absolute CD4+ T cells counts <350/mm3 (NCT00775606). The primary endpoint was the reduction in ex vivo CD4+ T cell apoptosis, with evaluation of the intrinsic and extrinsic pathways of apoptosis. Secondary endpoints included changes in absolute CD4+ T cell counts, T cell subsets, T cell activation, T cell turnover, and pathways of apoptosis.
Peripheral blood mononuclear cells (PBMCs) were obtained at baseline and at weeks 4, 12, and 24 after study entry and initiation of HAART. Ex vivo apoptosis of naive (CD4+CD45RA+) and memory (CD4+CD45RO+) CD4+ T cells and CD8+ T cells was examined by propidium iodide staining after magnetic-activated cell sorting and 72 h of incubation at 37°C in 5% CO2. 23
Pathways of apoptosis were examined by assaying activation of caspase 8, caspase 9, and effector caspases 3 and 7 by staining with carboxyfluorescein-labeled active caspase detection kits from APO LOGIX and analysis by flow cytometry. 24 Mitochondrial membrane potential was measured after staining with a fluorescent cationic dye JC-1 (APO LOGIX). 25 Changes in CD4+ and CD8+ T cell (naive and memory) subsets, cell activation, and cell turnover were characterized by polychromatic flow cytometry on cryopreserved PBMCs that were removed from liquid nitrogen storage, thawed rapidly in a 37°C water bath, washed, and then rested overnight in RPMI 1640 media containing 10% heat-inactivated fetal bovine serum (FBS) and 10 U/ml DNase at 37°C in 5% CO2. The following day, cells were washed and stained for viability with Aqua Live/Dead cell stain kit (Invitrogen) prior to cell surface staining with fluorochrome-conjugated monoclonal antibodies. The following day, cells were washed and stained with the Aqua Live/Dead cell stain kit (Invitrogen) to assess cell viability. PBMCs were then pretreated with human Fc block (Miltenyi Biotec) and stained with fluorochrome-conjugated antihuman monoclonal antibodies (Becton Dickinson, Pharmingen, eBioscience, Invitrogen) for cell surface or intracellular markers.
The following panels were used to assess T cell naive and memory subsets (CD3\CD4\CD8\CD45RA\CCR7), T regulatory cells (CD3\CD4\CD25\FoxP3), cell activation (CD3\CD4\CD8\HLA-DR\CD38), and cell proliferation (CD3\CD4\CD8\Ki67). After staining cells were washed, resuspended in 2% formaldehyde, and kept at 4°C until analyzed within 24 h on an LSR2 flow cytometer (BD). Analysis of flow cytometry data was performed using FACS Diva 6.1.1 software (BD) and FlowJo software (Tree Star, Inc.).
Fourteen patients entered the study before it was closed early for slow enrollment; 10 patients completed the treatment protocol, six in the LPV/r arm and four in the EFV arm. Nine of 10 patients achieved complete viral suppression, while one patient in the LPV/r arm had a decrease in viral load (VL) from 208,356 to 7,695 copies/ml by 24 weeks. The purpose of the study was to compare differences between LPV/r and EFV on immune recovery and the effects on apoptosis, but due to the small number of participant we were unable to show any differences between treatment group with respect to any of the endpoints for the study, i.e., ex vivo T cell apoptosis and CD4+ T cell recovery (see Table 1).
Statistically significant, p<0.05 by Student's t-test.
LPV-r, lopinavir/ritonavir; EFV, efavirenz.
Although patients in the LPV-r arm had a significantly higher CD4+ T cell count at 48 weeks compared to the EFV arm, they had a higher baseline count and the difference in the increase in CD4 count, 177 versus 127, was not statistically significant. Three patients had a <50 cell/mm3 increase despite complete viral suppression, including one patient with a CD4+ T cell count <200 cells/mm3 and two with counts between 200 and 350 cells/mm3 at baseline. One immune nonresponder was in the LPV-r arm and two were in the EFV arm. Our study was unable to show a difference in the PI-based versus NNRTI-based regimen with respect to effects on immune recovery and ex vivo T cell apoptosis, but it would be unlikely to demonstrate a difference given the small numbers in each arm. We had initially estimated that we needed 22 patients to complete both arms in order to demonstrate a significant difference in these parameters.
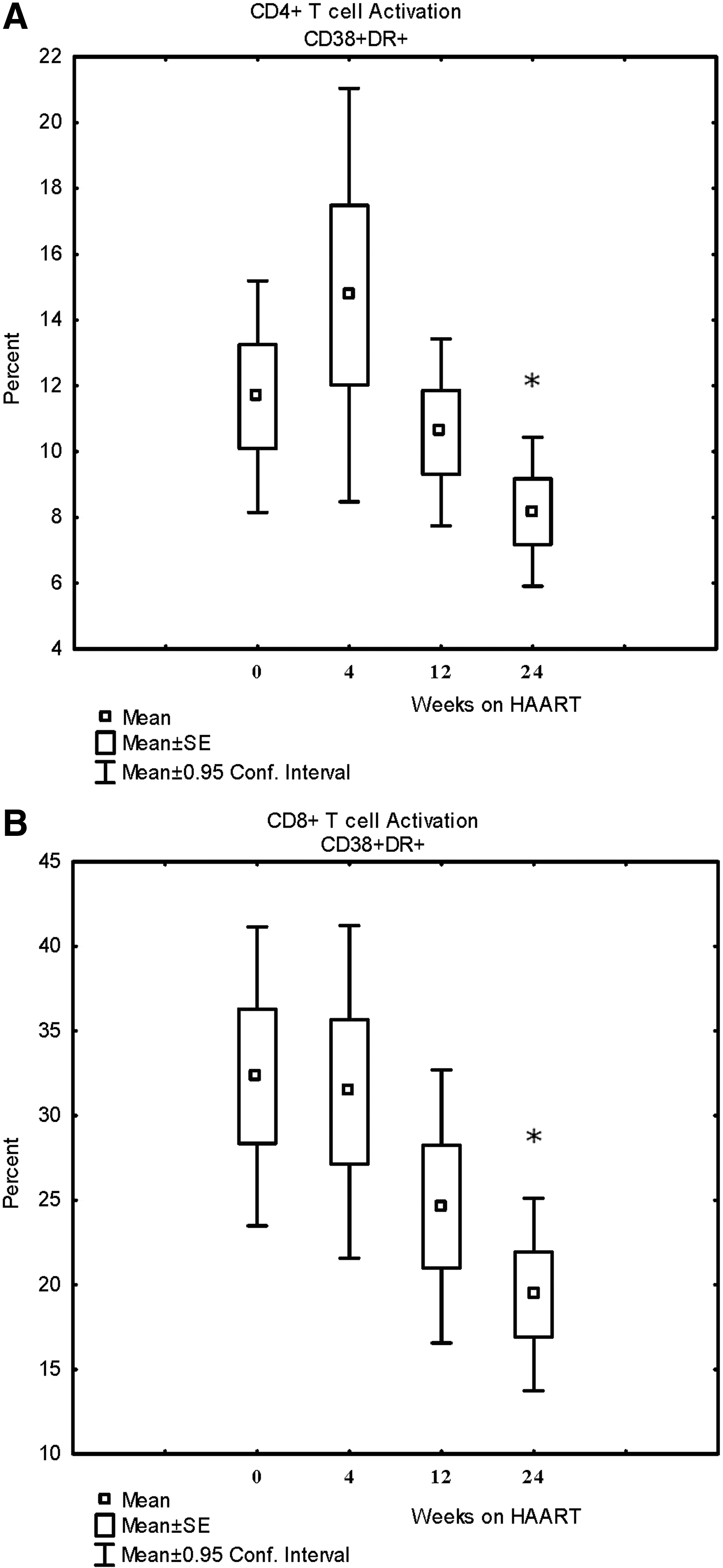
To determine the effects of HAART on T cell activation, proliferation, pathways of apoptosis, and immune recovery, regardless of the specific regimen, we analyzed these measurements for the entire cohort over 24 weeks of therapy. There were no significant changes in the percentages for the various CD4+ T cells subsets (naive, central memory, effector memory, and Treg T cells) at 24 weeks, although there was a trend toward a decrease in Tregs (see Fig. 1). All patients showed activation of CD4+ and CD8+ T cells at baseline as indicated by an increased percent of CD38+HLA-DR+ T cells (no control data, just comparisons over time; see Fig. 2). There was also increased proliferation and T cell turnover at baseline as indicated by the percent of Ki-67+ CD4+ and CD8+ T cells (no control data, just comparisons over time; Fig. 3). Activation of CD4+ and CD8+ T cells was significantly decreased after 24 weeks of HAART, and proliferation CD4+ and CD8+ T cells at both the 12-week and 24-week time points, but the reductions in activation and proliferation did not correlate with increases in absolute CD4+ T cell counts.

Changes in lymphocyte subsets before, during, and after 24 weeks of highly active antiretroviral therapy (HAART).
Changes in caspase 9 activation for naive (CD45RA+)

Proliferation of CD4+ T cells
All patients had an increased proportion of CD4+ and CD8+ T cells undergoing ex vivo apoptosis as indicated by DNA fragmentation by PI staining compared to HIV-negative controls at baseline, but this was significantly decreased after 24 weeks of HAART. Figure 4 shows the significant decrease in ex vivo apoptosis for naive (CD4+CD45RA+) T cells at 24 weeks and memory (CD4+CD45RO+) T cells at 12 and 24 weeks. CD8+ T cells also had a significant decrease in ex vivo apoptosis at 24 weeks (difference of −14±11%, p=0.01). All patients had increased activation of caspase 8, caspase 9, and caspases 3 and 7 at baseline compared to HIV-negative controls. The activation of all caspases was decreased by 24 weeks, but only the decrease in caspase 9 activation (intrinsic pathway) for naive CD4+ T cells was significant and there was a trend toward decreased caspase 9 activation at 24 weeks for memory T cells as well (see Table 2 and Fig. 5). This was associated with an increase in the mitochondrial membrane potential at 24 weeks for both naive and memory CD4+ T cells (see Fig. 6).
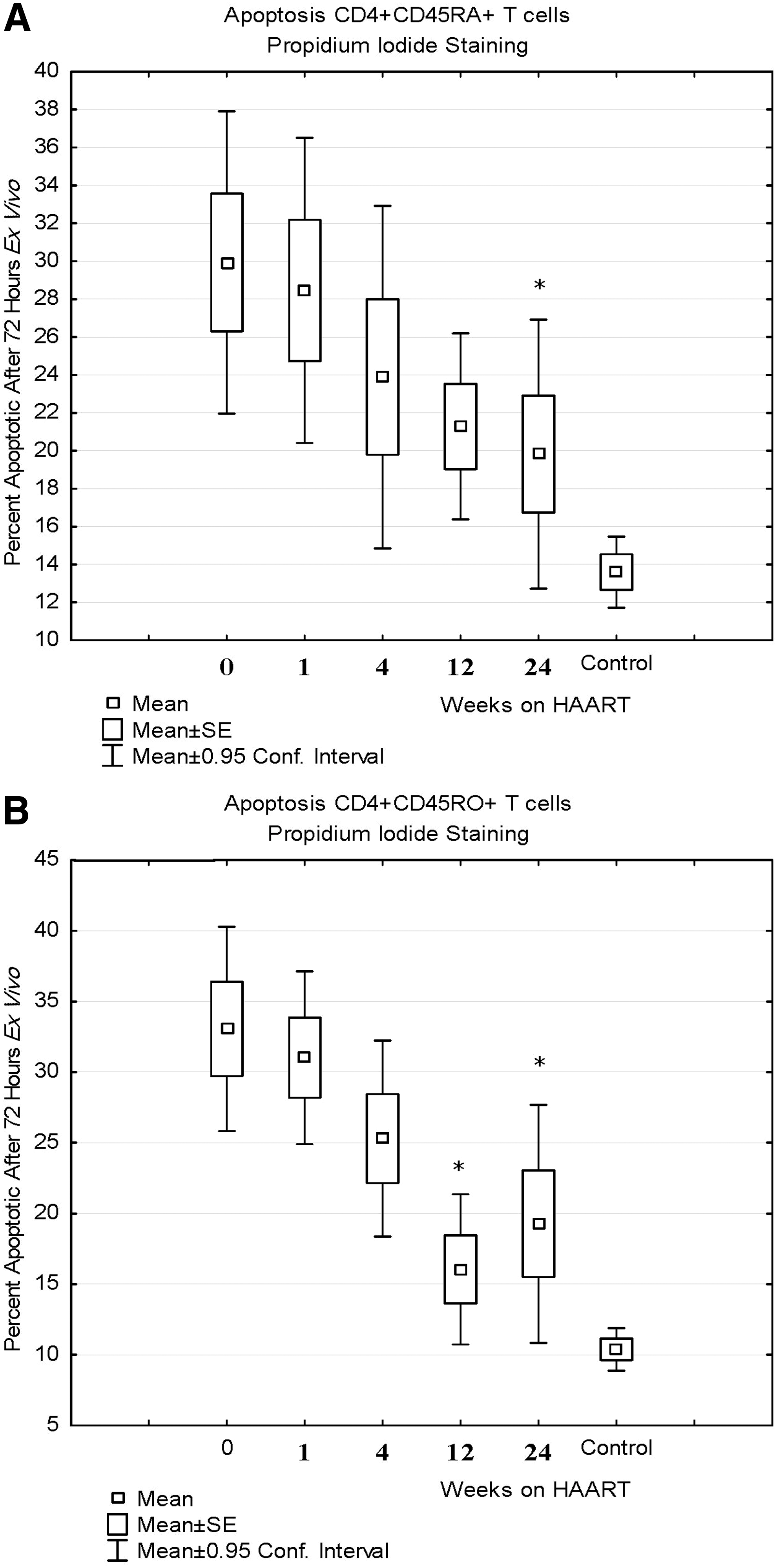
Ex vivo apoptosis as measured by propidium iodide staining for naive CD4+ T cells (CD4+CD45RA+)
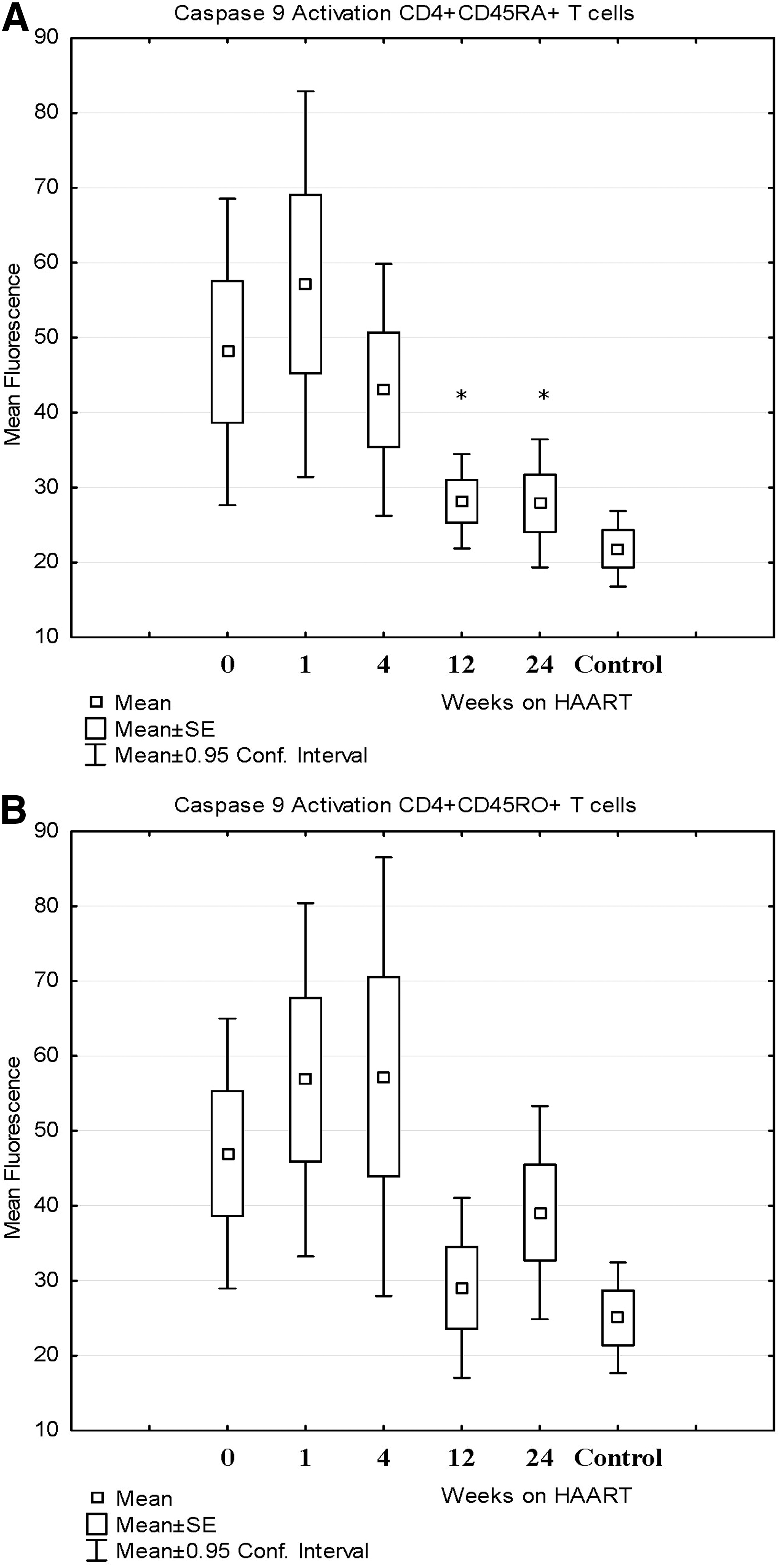
Caspase 9 activation expressed as mean fluorescence for CD4+ T cells compared to HIV-negative controls before and after HAART. Caspase 9 activation for naive CD4+ T cells (CD4+CD45RA+)

Mitochondrial membrane potential CD4+ T cells compared to controls before and after HAART. Membrane potential for naive CD4+ T cells (CD4+CD45RA+)
p<0.05, paired t test.
The increase in mitochondrial membrane potential for naive CD4+ T cells after 24 weeks of HAART was the only parameter that correlated with the increase in absolute CD4+ T cell count (r=0.66, p=0.038; see Fig. 7). Also, the three immune nonresponders with CD4+ T cell increase <50 cell/mm3 at 24 weeks had significantly lower increases in mitochondrial membrane potential at 24 weeks compared to the seven immune responders (–16.8±39.1 versus 166.2±89.2, p=0.017 by the Wilcoxon rank-sum test). Baseline CD4+ T cell count, change in activation, change in proliferation, and changes in the activation of individual caspases did not correlate with immune response.
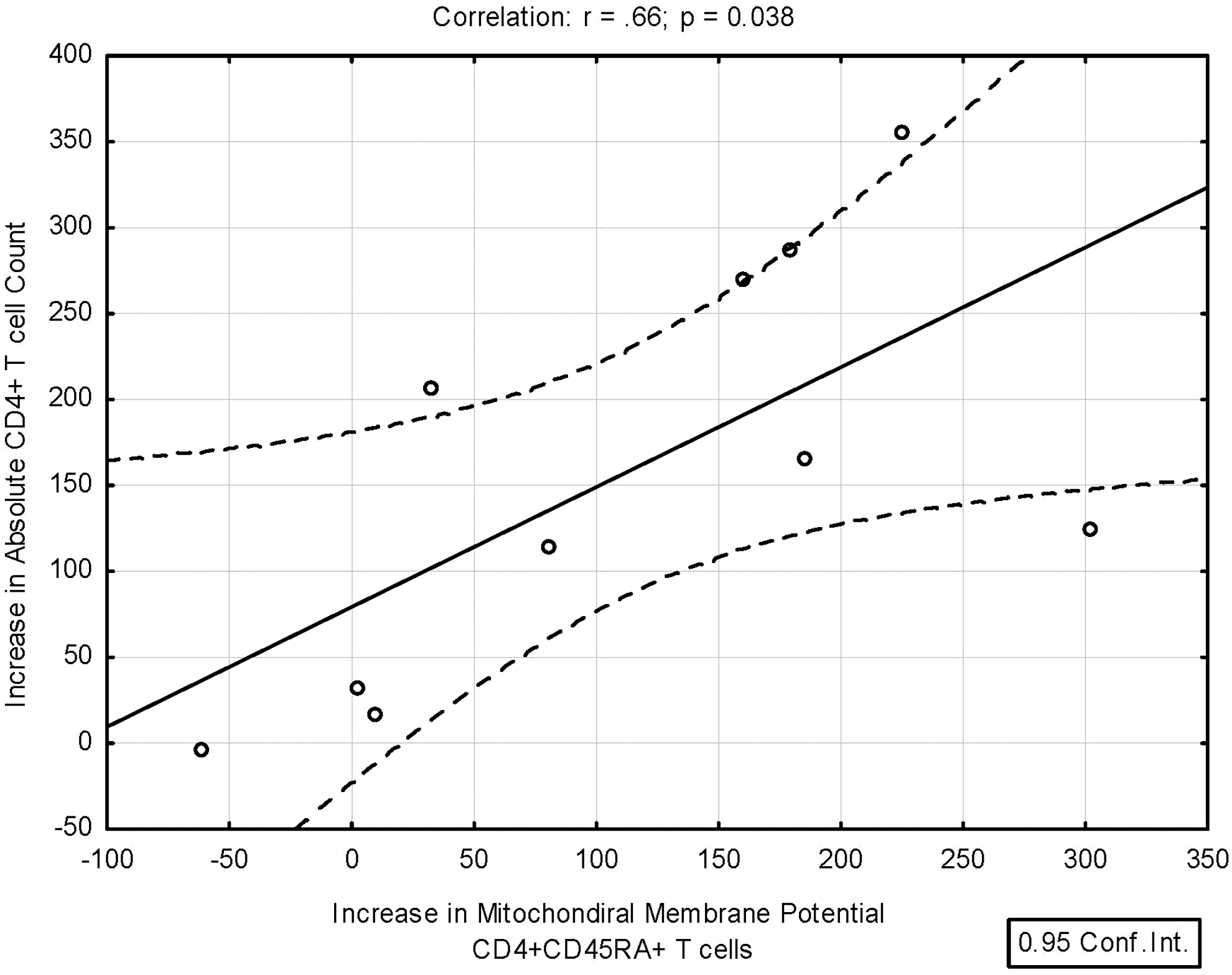
This graph shows the correlation between the increase in mitochondrial membrane potential for naive CD4+ T cells and the increase in absolute CD4+ T cell count after 24 weeks of HAART, r=0.66, p=0.038. The increase in the mitochondrial membrane potential for naive CD4+ T cells at 24 weeks was the only immune parameter measured that significantly correlated with immune reconstitution as measured by the increase in the absolute CD4+ T cell count. The three patients with <50 cell/mm3 increase in CD4+ T cells also had the lowest increase in mitochondrial membrane permeability after 24 weeks of HAART (p=0.017 by Wilcoxon rank-sum test).
HAART results in immune recovery and increases in absolute CD4+ T cell counts, but the degree of recovery is variable. Increase in mean CD4+ T cell counts by approximately 100 to 150 cells/mm3 over the first year with HAART and responses between 3 and 6 months may ultimately predict the degree of immune recovery. 26 A number of factors impact the degree of immune recovery patients enjoy on HAART, including viral factors, immune factors, and other factors. 7 Some patients are immunologic failures, usually defined as unable to increase their CD4 cell count by 25 to 50 cells/mm3 above baseline over the first year of therapy, and 15% to 20% of patients achieve a <50 cells/mm3 CD4+ T cell increase in the first 6 months of HAART despite achieving a virologic response. 3 –6 These discordant CD4+ T cell responses are associated with a higher risk of mortality. In the British Columbia Center for Excellence in HIV/AIDS cohort study (n=1,527), the relative risk of death was 1.87 (95% CI 1.15–3.04) and in the French Hospital Database on HIV (n=2,236), the relative risk was 1.98 (95% CI 1.26–3.10). 6
Immune activation or inflammation is important in HIV immune pathogenesis. There are heightened levels of proinflammatory cytokines in plasma, including interferon (IFN)-α, tumor necrosis factor (TNF)-α, and interleukin (IL)-6. 8 –11,26,27 There is also increased expression of the activation markers HLA-DR antigen and CD38 on CD4+ and CD8+ T cells. HAART results in a decrease in T cell activation and T cell proliferation/turnover, and other investigators have reported that expression of CD38, an activation marker, is associated with a less robust CD4+ T cell recovery. Our study also shows that immune activation is increased in HIV patients and this is reduced with effective HAART. HAART also reduces extrinsic and intrinsic apoptosis and increases mitochondrial membrane potential of naive and memory CD4+ T cells. The most important determinant of CD4+ T cell recovery after 24 weeks of HAART in our study, however, was the increase in mitochondrial membrane potential of naive CD4+ T cells at the 24 week time point. This is a small study, however, and further studies to confirm this observation are warranted, but other studies support this finding.
Negredo and co-workers have also conducted a study of CD4+ T cell death pathways and how this limits immune recovery in discordant immune responders, defined as a CD4+ T cell count persistently <350/mm3 despite suppressive HAART for >2 years. 28 This group found there was a greater correlation between immune response and intrinsic apoptosis of CD4+ T cells compared to extrinsic apoptosis due to activation. Nadir CD4+ count clearly correlated with immune response in both a univariate and multivariate analysis. Interestingly, in their univariate analysis, PI use was associated with a poor immune response, but PI use was more likely in patients with a low nadir CD4+ count and multivariate analysis did not show any effect of PI use on immune response. In our analysis, baseline CD4+ count did not predict poor immune recovery, but our study was not powered to detect such a difference. Nadir CD4+ counts for patients with optimal versus suboptimal immune recovery overlap significantly.
Other studies have shown that thymic dysfunction is a barrier to CD4+ T cell recovery, and our findings suggest that positive selection of naive CD4+ T cells in the thymus, rather than persistent immune activation, is the most important determinant of CD4+ T cell recovery. 29 –32 It is very likely that thymic recovery is less likely in patients with advanced HIV infection. Normal CD4+ T cell counts are achieved more frequently in patients who started HAART with a baseline CD4 >350 cells/mm3 and lower plateau CD4+ T cell counts are observed in patients initiating therapy at lower baseline counts even when durable viral suppression is achieved, but responses are variable. The increase in mitochondrial membrane potential observed in our study would best be explained by increased expression of antiapoptotic Bcl-2 gene family members in T cells being positively selected in the thymus, resulting in higher mitochondrial membrane potential and a reduction in the rate of apoptosis.
There are very few data on the relative contributions of immune activation or inflammation versus thymic dysfunction as barriers to immune recovery. Future studies of adjunctive therapies to aid immune recovery in HIV infection should include agents that enhance thymic function as well as agents aimed at decreasing immune activation, although previous studies of immune-based therapies have been disappointing. 33
The finding that the change in mitochondrial membrane potential for naive CD4+ T cells accurately predicts CD4+ T cell recovery suggests that this measure is a potentially useful surrogate marker for thymic function in HIV infection and perhaps in other conditions where thymic dysfunction is important. Other measures of thymic function, such as computed tomography (CT) of the thymus and 3D reconstruction to estimate thymic mass or measurement of T cell receptor excision circles (TRECs), are either expensive or technically difficult. 34,35 Mitochondrial membrane potential is easily performed in any laboratory that has flow cytometry capability. Further studies are needed to determine if mitochondrial membrane potential of naive CD4+ T cells is a reliable surrogate marker of thymic function that may help guide future studies of immune-based therapies.
Footnotes
Acknowledgments
We would like to thank the patients who participated in this study and the study staff at the clinical trial sites. Data were presented at the Annual IDWeek 2012, Session 81, HIV Pathogenesis and Transmission, October 18, 2012, San Diego, CA, Abstract 592.
Author Disclosure Statement
Funding support for this trial was provided by Abbott Laboratories. Medications were supplied by Abbott Laboratories and Gilead Sciences. Trial Registration Number: NCT00775606.