
Abstract
The color of normal skin and of oral mucosa is not determined by the number of melanocytes in the epithelium but rather by their melanogenic activity. Pigmented biopolymers or melanins are synthesized in melanosomes. Tyrosinase is the critical enzyme in the biosynthesis of both brown/black eumelanin and yellow/red pheomelanin. The number of the melanosomes within the melanocytes, the type of melanin within the melanosomes, and the efficacy of the transfer of melanosomes from the melanocytes to the neighboring keratinocytes all play an important role in tissue pigmentation. Melanin production is regulated by locally produced factors including proopiomelanocortin and its derivative peptides, particularly alpha-melanocyte-stimulating hormone (α-MSH), melanocortin 1 receptor (MC1R), adrenergic and cholinergic agents, growth factors, cytokines, and nitric oxide. Both eumelanin and pheomelanin can be produced by the same melanocytes, and the proportion of the two melanin types is influenced by the degree of functional activity of the α-MSH/MC1R intracellular pathway. The cause of HIV oral melanosis is not fully understood but may be associated with HIV-induced cytokine dysregulation, with the medications commonly prescribed to HIV-seropositive persons, and with adrenocortical dysfunction, which is not uncommon in HIV-seropositive subjects with AIDS. The purpose of this article is to discuss some aspects of melanocyte biology and HIV-associated oral melanin hyperpigmentation.
Introduction
O
The main functions of melanocytes are to produce melanin, which determines the color of skin, hair, and eyes, to confer photoprotection, to combat oxidative stress by scavenging reactive oxygen species, 10,11 and to neutralize microorganisms and their harmful products. 12 Melanocytes contribute to the homeostasis of the tissues in their immediate vicinity by subverting some functions of neuroendocrine cells by secreting neuropeptides, and as immunocompetent cells capable of phagocytosis and of antigen presentation. 12,13
Histologically and ultrastructurally, oral melanocytes resemble epidermal melanocytes. 14,15 However, the baseline metabolic activity of oral melanocytes is generally lower than that of epidermal melanocytes, but oral melanocytes can become more active in melanin biosynthesis in response to genetic factors (physiological pigmentation), noxious habits (smoking), drugs, injury, a range of pathological conditions (infections, inflammation, hormonal dysfunctions, neoplasms), and perhaps to other environmental stimuli. 16 There are no numerical or structural differences in oral melanocytes between dark-skinned and light-skinned persons except that in the former the melanosomes, the structural organelles in which the melanin is synthesized, are larger and more numerous. 16 –18 Physiological differences in color of skin, and probably of oral mucosa, are largely determined by type and amount of melanin produced, 19 by the number and size of the melanosomes, and by the efficacy of distribution of melanosomes to neighboring keratinocytes. 20 Variation in the color of the oral mucosa will be affected by the degree of vascularization of the tissues, by the level of hemoglobin in the blood, by melanin pigmentation, and by the masking effect of keratin if the epithelium is heavily keratinized. 13
The Biology of Melanocytes
During embryogenesis, melanocyte precursor cells migrate from the neural crest to reach their final destination in the basal cell layer of the epithelium of the skin and of mucous membranes including oral mucosa, as well as in the choroid and iris of the eye, in the leptomeninges, and in the cochlea of the inner ear. 11,21,22
It has been shown that in the skin, melanocyte precursors may originate not only from the neural crest, but also from neural crest-derived Schwann cell precursors. 23 Mature melanocytes in the basal cell layer of the epithelium are in contact with about 35 keratinocytes via their dendritic processes through which they transfer melanosomes to the keratinocytes in the suprabasal layers of the epithelium. This complex is commonly referred to as an epidermal melanin unit, 24 but might perhaps better be called a “melanocyte-keratinocyte unit.” 13 Whether these observations can be extrapolated to oral mucosa needs further research.
Under physiological conditions melanocytes have a low mitotic rate, and it is uncertain whether the increase in the number of melanocytes in response to ultraviolet radiation (UVR) or to neuroendocrine agents is the result of an increase in the rate of mitosis of existing mature melanocytes or of stimulation and subsequent proliferation and maturation of undifferentiated melanocyte precursors. 25 In skin, melanocyte stem/progenitor cells reside in the epithelial bulge of the hair follicle, 13,26,27 and replacement of lost melanocytes owing to injury or apoptosis occurs by migration of melanocyte stem/progenitor cells from these niche reservoirs. However, the mechanism whereby stem/progenitor melanocytes are able to maintain the population of melanocytes of the oral mucosa is unknown, 27 and indeed niche reservoirs of melanocyte stem cells in oral mucosa have not been found.
PAX3, TCF/LEF (T cell factor/lymphoid enhancer factors), and SOX10 are transcription factors expressed during early embryogenesis, and are required for survival and migration of neural crest-derived melanocyte precursors. These factors also have the capacity to mediate the expression of microphthalmia-associated transcription factor (MITF), which not only is the master regulator of genes involved in melanogenesis, but is also important for the development of melanocytes. 20 In addition, the survival and migration of neural crest-derived melanocyte precursors during development also require interactions between the melanocyte tyrosine kinase receptor C-Kit and its extracellular ligand, stem cell factor (SCF), as well as between endothelin 3 and its endothelin receptor B (EDNRB). 20,28
The Wnt/β-catenin signaling pathway is essential for melanocyte development. 20 Under physiological conditions, β-catenin, a key protein in the canonical Wnt signaling pathway, plays an important role in MITF-mediated melanocyte differentiation, proliferation, and melanogenesis. 29 Upon stimulation of the Wnt receptor, Frizzled, there is an accumulation of β-catenin in the cytoplasm, which then translocates to the nucleus where it binds to TCF/LEF1, which together induce the transcription of MITF. On the other hand, Dickkopf 1, an inhibitor of the Wnt/β-catenin pathway, upon interaction with Wnt receptor lipoprotein receptor-related protein 6 (LRP6), 30 suppresses the differentiation and functions of melanocytes. 30,31
Activation of the Wnt signaling pathway in keratinocytes brings about production and secretion of endothelins. In turn, activation of the endothelin/endothelin receptor pathway in melanocytes promotes their proliferation, differentiation, and melanin production. 31 However, mesenchymal–epithelial interactions via fibroblast-derived Dickkopf 1 expressed in the microenvironment of the melanocytes suppress the expression of β-catenin with downregulation of MITF activities, so that melanocyte progenitors remain undifferentiated. 29 –32 It appears that fibroblast-derived Dickkopf 1 also has the capacity to inhibit the transfer of melanosomes from melanocytes to keratinocytes by downregulating the expression of the protease-activated receptor-2 (PAR-2) on keratinocytes. This results in decreased phagocytic uptake of melanin by keratinocytes. 32
The process of melanocyte precursor migration from the dermis into the epidermis is facilitated by the expression of E-cadherin molecules and by the interaction between SCF, endothelin 1 and 3, hepatocyte growth factor (HGF), and basic fibroblast growth factor (bFGF) secreted in the microenvironment, with their respective receptors on melanocytes. 11,20
Later, during functional activity, signals from neighboring keratinocytes, from the microenvironment, and also from autocrine signals regulate melanocyte survival, proliferation, dendritic development, and melanogenesis. 33,34 There is a long and varied list of agents that regulate the functional activity of melanocytes including proopiomelanocortin (POMC)-derived peptides, endothelin 1, SCF, inflammatory mediators [i.e., interleukin (IL)-1, tumor necrosis factor (TNF)-α, and tumor growth factor (TGF)-α], neurotrophins, nerve growth factor (NGF), keratinocyte growth factor (KGF), bFGF, and nitrous oxide (NO). 11
Some melanocyte precursors fail to reach their destination in the basal cell layer of the epithelium because, for reasons unknown, they become arrested in the dermis/lamina propria. 13 These melanocyte precursors may mature, become active, and give rise to intradermal or intralamina proprial pigmented nevi. HGF, bFGF, and possibly other biological mediators within the microenvironment of these arrested melanocyte precursors may be important for their survival, proliferation, and maturation. 25
The number of melanocytes in the basal cell layer of the epithelium varies in different skin or oral mucosal sites, but is similar in corresponding skin or oral mucosal sites of different persons, regardless of their racial/ethnic origin. 13,35 The most important factor governing the quantum of melanin of any particular epithelial site is therefore not the number of melanocytes but rather their degree of melanogenic activity, which is genetically determined. Melanin pigmentation in any individual depends on the inherent baseline level of activity of the melanogenic enzymes and on how these enzymes respond to intrinsic and extrinsic agents as mentioned above; on the functional activity of the extracellular ligand/receptor interactions mediating pathways of melanin biosynthesis; and on the degree of maturation of the melanosomes and on the extent of arborization of the melanocytic dendritic processes. 13,27
Mesenchymal–epithelial interactions are important for melanocyte development and function. 29,36 In vitro extracellular matrix proteins of cultured fibroblast and other biological mediators promote melanocyte survival and differentiation. 36,37
Intraepidermal free nerve endings containing calcitonin gene-related peptide, substance P, or neuropeptide Y are in intimate contact with epidermal melanocytes, and it has been suggested that these neuropeptides have the capacity to modulate the functional activity of epidermal melanocytes. 38
Melanin Biosynthesis and Distribution
Synthesis of melanin occurs in specialized membrane-bound organelles termed melanosomes. Melanosomes contain both the structural matrix proteins that form a scaffold for melanin deposition as well as all the enzymes and proteins necessary for melanin biosynthesis. The development and maturation of melanosomes progress through a series of well-characterized steps from endosomal precursors through proteinaceous fibrillar structures to fully melanized organelles in which the structural proteins become masked by the deposition of melanin. 39
There are two chemically distinct types of melanin, brown/black eumelanin and red/yellow pheomelanin. 20 Both of these are derivatives of the amino acid tyrosine via the intermediate step of conversion of dopa to dopamine. Tyrosinase as well as tyrosinase-related protein 1 (TYRP1) and TYRP2 are important regulatory enzymes in the pathway of melanin biosynthesis. 40 The shapes of pheomelanosomes and eumelanosomes are said to differ, with eumelanosomes having an elliptical shape and longitudinal deposition of melanin 41 –43 and pheomelanosomes having a spherical shape with granular deposition of pigment. 42 Eumelanin and pheomelanin are both produced by the same melanocytes. 40 Melanogenesis should thus be regarded as “mixed melanogenesis,” with the proportions of pheomelanin and eumelanin being genetically determined by tyrosinase activity but also by the availability of tyrosine and cysteine in the melanosomes. 44
In general, melanogenesis is regulated by several local factors including POMC and its derivative peptides, particularly alpha-melanocyte-stimulating hormone (α-MSH); by the activity of melanocortin-1 receptor (MC1R) on melanocytes; and by cytokines and growth factors in the microenvironment. 35,45,46 These factors have the capacity to activate MITF, which in turn regulates genes encoding enzymes driving the process of melanogenesis, including tyrosinase, TYRP-1, and TYRP-2. 19 The functional activity of tyrosinase in the complex conversion of tyrosine to melanin is supported by p protein and membrane-associated transporter protein (MATP), which provide an appropriate ionic environment. 47 Control of MITF, which plays an important role in regulating signaling pathways in melanogenesis, is itself regulated by factors produced by both keratinocytes and fibroblasts. 44
The transport of melanosomes within the melanocyte is governed by the motor proteins kinesin and dynein. These proteins convey the melanosomes from the region of the nucleus of the melanocyte into and along the dendritic microtubules, and by means of a tripartite complex 48 along actin filaments 39 toward the tip of the dendrites. 49 Thus, the route of melanosomal distribution is from a perinuclear position proceeding peripherally, with the mature melanosomes accumulating at the extremity of the dendrites. 39 When the melanosomes reach the tips of the dendrites, they become bound to the actin-rich dendritic membrane by myosin V filaments 49,50 and are externalized by a process that as yet is unclear.
The transfer of melanosomes externalized from the arborizing dendritic processes to the neighboring keratinocytes in the keratinocyte melanin unit is influenced by the functional activity of PAR-2 and by keratinocyte growth factor receptors on the keratinocytes, followed by phagocytosis. 25,51,52 In addition, unidentified surface lectins and glycoproteins are said to facilitate the transfer. 53
Epidermal melanocytes express POMC-derived peptides including adrenocorticotropin (ACTH), α-MSH and β-MSH, and β endorphins. In particular, the αMSH/MC1R and the β endorphin/μ opiate receptor signaling pathways are important in the regulation of proliferation and differentiation of melanocytes and of melanin biosynthesis. 45
Melanogenesis depends on external stimuli such as UVR or on internal factors such as paracrine and endocrine agents as outlined above. Melanogenesis occurs through the stimulation of the MC1R by α-MSH, which then induces the production of cAMP with activation of the transcription factor CREB (cAMP responsive element-binding protein). This cascade of molecular events will activate MITF resulting in eumelanin production. On the other hand, agouti signaling protein (ASIP) can also bind to MC1R, blocking MC1R-induced activation of cAMP. 20,47 However, melanogenesis may also be controlled by α-MSH independently of MC1R. 54
In white persons, the MC1R is highly polymorphic and its germline variants are associated with different degrees of loss of functional activity of its intracellular signaling pathways. This leads to variations in melanogenic capacity and in the spectrum of distinct phenotypic traits such as red hair, freckling, and fair skin. 55 Activation of the common MC1R promotes eumelanin synthesis while MC1R polymorphism is associated with increased pheomelanin and decreased eumelanin production, and consequently with a light pigment phenotype. 56,57
Both adrenergic and cholinergic nonneural pathways are expressed in melanocytes, and when activated may promote melanogenesis. α1- and β2-adrenoceptors on melanocytes when activated by noradrenalin and adrenalin, respectively, have the capacity to trigger both melanogenesis and arborization of dendrites; and when nicotine receptors on oral melanocytes are activated by acetylcholine or by nicotine they can initiate induction of hyperpigmentation. 58
It appears that NO also plays a role in the regulation of melanogenesis. In response to UVR or inflammatory agents, melanocytes express NO synthetase (NOS1), which brings about an increased production of NO. 11,35 In turn, NO promotes the maturation and transport of melanosomes, upregulates tyrosinase gene expression, and enhances the activity of the tyrosinase. Overall, NO promotes melanin production by melanocytes. 59
The importance of pH in melanogenesis was indicated by Fuller et al. (2001). 60 The melanosomes derived from light human skin are more acidic than those derived from dark human skin. While the mechanisms and molecules engaged in the regulation of melanosomal pH are not known, vacuolar ATPase 44 and p protein appear to be important. 25
The Functions of Melanocytes and Melanins
Epidermal melanocytes are both a source of, and a target for, neurotransmitters, hormones, and other biological mediators. These play a role in the local innate immune system as well as in protection against both biochemical and physical insults as well as microbial infections. 40 Furthermore, they have the capacity to phagocytose microorganisms, to process foreign antigens, to function as antigen-presenting cells, and to produce cytokines such as IL-1 and IL-6, thus contributing to the maintenance of tissue homeostasis. 12
Melanins are complex polymers that are photoprotective, can inactivate reactive oxygen species (ROS) and free radicals, and can sequestrate redox-active metals and organic toxic compounds. Melanin through its strong binding capacity acts as a physical barrier to microorganisms penetrating the epidermis, and it neutralizes microorganismal toxins. The process of melanin biosynthesis, however, generates free radicals, 12 but the potential toxicity of these free radicals is outweighed by their antimicrobial properties.
Inflammation-Associated Melanin Pigmentation
Histamine, arachidonic acid metabolites, and some proinflammatory cytokines have the capacity to stimulate melanogenesis and/or proliferation of melanocytes. Melanin pigmentation is thus sometimes observed where the oral mucosa has been traumatized or has been chronically inflamed owing to conditions such as lichen planus. Inflammation-associated melanin pigmentation usually appears as single or multiple light-brown patches, occurring most commonly in darker skinned persons. 61 UVR and inflammatory mediators can induce PGE2 production by keratinocytes and melanocytes, which in turn stimulates proliferation and arborization of the melanocytic dendrites. PGE2 also promotes tyrosinase activity and proliferation of melanocytes independently of the α-MSH/MC1R pathway. 62
In response to inflammatory mediators, keratinocytes and melanocytes produce α-MSH, which promotes melanin production, 9,63 and by blocking NF-κB in inflammatory cells, downregulates the expression of proinflammatory cytokines such as IL-α, IL-β, IL-2, IL-6, and TNF-α, 64 upregulates the expression of the antiinflammatory cytokine IL-10, and modulates the function of monocytes, macrophages, and dermal microvascular endothelial cells. Thus, α-MSH may elicit antiinflammatory and immunosuppressive responses. 64,65
Melanin Hyperpigmentation in HIV-Seropositive Subjects
In some HIV-seropositive subjects, brown/black melanotic maculae may appear on the oral mucosa (HIV-OMP) (Fig. 1) as a result of an increase in melanin not only in the basal cell layers of the epithelium, but also in melanophages in the upper lamina propria. 2 HIV-OMP may be induced by drugs such as zidovudine, Clofazimine, and ketoconazole, taken for the treatment of the HIV disease or HIV-associated conditions, may occur because of adrenocortical insufficiency that is not uncommon in AIDS, or may be idiopathic. 1,2,66,67
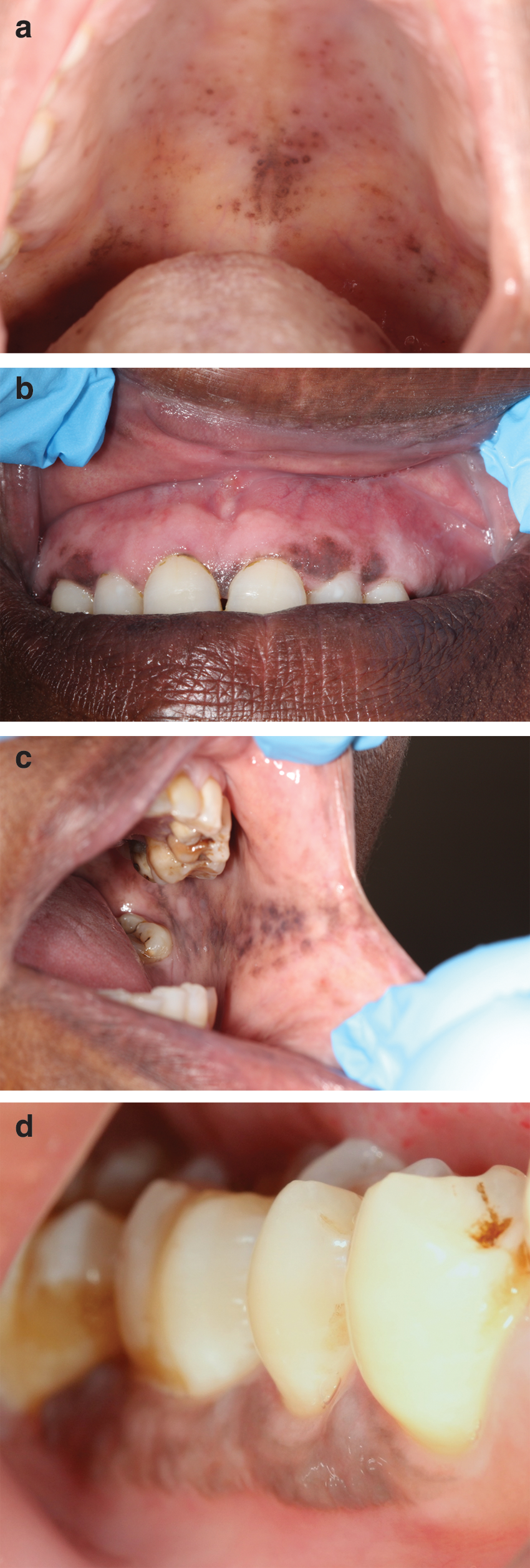
HIV-associated oral melanin hyperpigmentation (HIV-OMP).
The onset of HIV-OMP if it arises occurs within a period of 2 years of the diagnosis of HIV infection. 3,66 It is possible that HIV-induced upregulation of IL-1, IL-6, and TNF-α promotes the production of α-MSH by oral keratinocytes and melanocytes, and induces upregulation of the expression of MC1R of melanocytes 9,63 with an increased production of melanin resulting in the development of HIV-OMP. HIV-induced cytokine dysregulation roughly parallels a decreasing CD4+ T cell count, 68 so that oral melanin hyperpigmentation is observed more frequently in HIV-seropositive subjects with a CD4+ T cell count of 200 cells/mm3 or less. 66
Mucosal inflammation, as mentioned above, can promote melanin pigmentation. As there are more immunoinflammatory cells in the subepithelial connective tissue of clinically healthy oral mucosa of HIV-seropositive subjects, probably as a subclinical response to putative exogenous antigens, it is likely that in some cases HIV-OMP is brought about by inflammatory or postinflammatory states. 2 Although it is clear that HIV does not infect melanocytes, because the receptors for HIV are lacking, it is not known whether HIV proteins have the capacity to activate melanocytes directly.
While in idiopathic HIV-OMP the pigmentation is the result of increased production of melanin without an increase in the number of melanocytes, it appears that the mucocutaneous pigmentation associated with the antiretroviral drug zidovudine is caused either by an increase in the number of melanocytes in the basal and suprabasal cell layers of the epithelium 69 or by increased melanogenesis of the normal resident melanocytes. 70 In general, the extent and intensity of zidovudine-induced mucocutaneous pigmentation are greater in darker skinned persons. It has been reported that if it occurs, zidovudine-induced OMP appears about a month after the medication is started. 70,71 When zidovudine is discontinued, usually there will be a gradual diminution of the hyperpigmentation.
Thus, both HIV-induced cytokine dysregulation and zidovudine have the capacity to activate melanocytes with increased production of melanosomes containing melanin. 63
Clinically and histologically, HIV-OMP and melanotic maculae or physiological pigmentation are very similar; the distinction can be made only if the pigmentation appears after the diagnosis of HIV infection or after HIV-related medication has been started. HIV-OMP is common, affecting up to 30% of HIV-seropositive subjects. Typically it appears as single, or as multifocal macules on the buccal mucosa, 8,9,72 affecting males and females equally. 7
HIV-OMP must also be differentiated from the pigmentation associated with systemic disorders such as Addison's disease, acromegaly, Albright's disease, or neurofibromatosis, or from local lesions including inflammation-associated pigmentation, drug-induced melanosis, smoker's melanosis, atypical melanocytic proliferation, or melanomas. 2,61
Whether oral melanin hyperpigmentation has any clinical and pathological significance in HIV-seropositive subjects is unknown and it is uncertain whether it is caused by an increase in the number of melanocytes, by an increase in the number of melanosomes, by hyperarborization of melanocytic dendrites accelerating the intercellular distribution of melanin, by locally increased adrenergic or cholinergic stimulation of melanin production by resident melanocytes, or by increased activity of melanocortins. Further research is needed to elucidate these matters. 2
Further research is also needed to investigate whether HIV-OMP is a side-effect of any adventitious subclinical infection or inflammatory process, or whether the melanin is a protective immune response against the noxious effects of oral pathogens. 73
Summary
Melanosome biogenesis, the rate of melanin biosynthesis, and the efficacy of the transfer of melanosomes from melanocytes via melanocytic dendrites to the neighboring keratinocytes in the melanin keratinocyte unit are genetically determined, but can be modified by multiple factors including UVR, infection, noninfectious inflammatory processes, and keratinocyte- and fibroblast-derived biological mediators.
The causes of idiopathic HIV-associated melanin pigmentation are unknown, but it is likely that HIV-associated cytokine dysregulation plays an important role. It is possible that idiopathic HIV-associated melanin hyperpigmentation represents either a protective immune response against oral pathogens in which the melanin inactivates noxious products of the pathogens, or it may simply be a side-effect of some subclinical infection and its concomitant inflammatory process.
Footnotes
Author Disclosure Statement
No competing financial interests exist.