
Abstract
Persons living with HIV/AIDS (PLWHA) frequently use cannabinoids, either recreationally by smoking marijuana or therapeutically (delta-9-tetrahydrocannabinol; Δ9-THC dronabinol). Previously, we demonstrated that chronic Δ9-THC administration decreases early mortality in male simian immunodeficiency virus (SIV)-infected macaques. In this study, we sought to examine whether similar protective effects resulted from chronic cannabinoid administration in SIV-infected female rhesus macaques. Clinical and viral parameters were evaluated in eight female rhesus macaques that received either Δ9-THC (0.18–0.32 mg/kg, intramuscularly, twice daily) or vehicle (VEH) starting 28 days prior to intravenous inoculation with SIVmac251. SIV disease progression was assessed by changes in body weight, mortality, viral levels in plasma and mucosal sites, and lymphocyte subsets. In contrast to our results in male animals, chronic Δ9-THC did not protect SIV-infected female rhesus macaques from early mortality. Markers of SIV disease, including viral load and CD4+/CD8+ ratio, were not altered by Δ9-THC compared to control females; however, females that received chronic Δ9-THC did not gain as much weight as control animals. In addition, Δ9-THC administration increased total CXCR4 expression in both peripheral and duodenal CD4+ and CD8+ T lymphocytes prior to SIV inoculation. Although protection from early mortality was not evident, chronic Δ9-THC did not affect clinical markers of SIV disease progression. The contrasting effects of chronic Δ9-THC in males versus females remain to be explained, but highlight the need for further studies to explore the sex-dependent effects of Δ9-THC and other cannabinoids on the HIV disease course and their implications for virus transmission.
Introduction
S
Cannabinoids exert their actions largely through two cannabinoid receptors, CB1 and CB2, which are expressed in the central nervous system and in peripheral tissues. 13 –15 CB1 receptors mediate the psychotropic effects of cannabinoids, while CB2 receptors have been shown to produce immunomodulatory effects. 16,17 CB2 receptors are expressed on both innate and adaptive immune cells in which the level of expression is dependent upon a cell's activation state. 18,19 In vitro and in vivo studies have shown that cannabinoids exert diverse immunomodulatory effects, including reducing cellular proliferation, shifting the Th1/Th2 balance, altering immune cell function, and affecting cytokine production. 17,19,20 Animal studies have also shown that immunomodulation by cannabinoids can decrease resistance to bacterial, parasitic, and viral infections. 21 –25 The immunosuppressive effects of Δ9-THC alter host–pathogen interactions and thereby impact disease pathogenesis. 26,27 Thus, the potential interaction of cannabinoids with HIV disease progression has been the focus of scientific attention. 9
Limited clinical investigations have evaluated the impact of cannabinoids on HIV disease progression. For example, an early study noted that marijuana use was associated with progression of HIV-seropositive subjects to end-stage AIDS. 28 In contrast, Abrams et al. evaluated the effects of both smoked and oral Δ9-THC on HIV infection in a randomized, placebo-controlled study and found that short-term (21 days) administration had no harmful effects on plasma viral levels or CD4+ T lymphocyte counts. 29 These data were also consistent with our previous studies utilizing a well-established nonhuman primate model, which examined the effects of chronic cannabinoid use on HIV disease progression. Chronic daily administration of Δ9-THC (0.32 mg/kg twice a day) to simian immunodeficiency virus (SIV)-infected male rhesus macaques provided protection from early mortality, along with attenuated plasma viral levels and retention of body mass. 30 More specifically, reduced viral loads were seen in tissue from the brain, spleen, and lymph nodes of THC/SIV+ animals relative to controls, and this was associated with decreased expression of tissue proinflammatory cytokines and decreased intestinal cell death. 15,31,32 Together, these findings demonstrated that Δ9-THC could attenuate SIV disease progression in male rhesus macaques. A similar investigation had not yet been conducted to determine if chronic Δ9-THC would produce the same effects in SIV-infected female rhesus macaques.
Cannabinoids have clearly been shown to exert sex-dependent biological and behavioral effects. In humans, marijuana abuse is more prevalent in males than females, which suggests potential differences in both the reinforcing and subjective effects of Δ9-THC. 33 –35 In animals, both behavioral and pharmacodynamic data support a direct interaction with gonadal hormones. 36 For example, Daniel et al. used a within-subject design to show that estradiol administration attenuated the disruptive effects of Δ9-THC in ovariectomized (OVX) female rats responding in a complex learning task. 37 These data are also supported by pharmacodynamic data indicating that ovarian hormones can either inhibit or antagonize cannabinoid signaling in areas of the brain important for cognitive functioning, such as the hippocampus and striatum. 38 Moreover, estrogens appear to produce the attenuation of cannabinoid signaling in multiple ways, including reducing guanosine-5′-O-(3-thio)triphosphate (GTPγS) binding or coupling of CB1 receptors to signaling pathways, modification of CB1 receptor mRNA and protein expression, altering the relative affinity of agonists for CB1 receptors, and affecting cochaperones such as activator of heat-shock 90-kDa protein ATPase homolog 1 (AHA-1) that move CB1 receptors to the cell surface from subcellular locations. 39 –43
Taken together, reports in the literature suggest a potentially antagonistic interaction between the estrogens and cannabinoids. The aim of this study was to determine if chronic Δ9-THC administration would confer protection from SIV disease in female rhesus macaques similar to that seen in SIV-infected males. 30 To accomplish this, female rhesus macaques were administered the same chronic regimen of Δ9-THC as used previously with male rhesus macaques and then inoculated with SIV. Both clinical and viral parameters were assessed throughout a 12-month period of SIV infection.
Materials and Methods
Cohort and sample collection
Eight 4-year-old female rhesus macaques of Indian origin (Macaca mulatta) were obtained from the California National Primate Center and served as subjects. These animals were housed individually at the LSU Health Sciences Center Animal Care Facility in New Orleans, LA (LSUHSC-NO) and maintained at approximately 90% of their free-feeding body weights on a diet consisting of banana-flavored food pellets (Purina Mills TestDiet, Richmond, IN), standard primate chow (Formula 2050, Harlan Teklad, Madison, WI), vitamins, and fresh fruit. This modest food restriction was used to maintain consistent levels of responding in food-motivated behavioral tests during 5 days of the week (Monday through Friday). All experiments were conducted in accordance with the LSUHSC-NO Institutional Animal Care and Use Committee and followed the guidelines provided by the National Institutes of Health (NIH) for use of experimental animals.
Animals were assigned to receive vehicle (VEH, n=4) or Δ9-THC (THC, n=4), provided by the National Institute on Drug Abuse (Research Technical Branch, Rockville, MD). The Δ9-THC was prepared as an injectable emulsion using alcohol, emulphor, and saline (1:1:18) as the vehicle, and both Δ9-THC and its vehicle were administered chronically via intramuscular injection twice daily throughout the duration of the study as previously described. 15,44 The initial dose of Δ9-THC was 0.18 mg/kg and this dose was increased during the first 2-week period of the study to a final dose of 0.32 mg/kg. After 28 days of chronic administration (T=0), all animals were intravenously inoculated with 100 TCID50 (50% tissue culture infective dose) of SIVmac251. An in vitro SIV infection assay that predicts viral replication kinetics was used to stratify the animals across the two treatment groups to optimize the small cohort size and address the variability associated with SIV disease progression among rhesus macaques. 45
Animals in both experimental groups, VEH/SIV and THC/SIV, were monitored daily by trained technicians and received weekly physical examinations by a veterinarian that included assessment of body weight and physical condition. Blood was collected in EDTA-containing tubes at these times and an aliquot of whole blood was removed for flow cytometry prior to centrifuge separation for plasma specimens. Duodenal pinch biopsies were acquired using a flexible endoscope (GIF-Q180, Olympus America Inc., Melville, NY) at discrete time points as previously described. 32 Vaginal fluids were serially sampled at the time of blood collection by rotating a polyester-tipped swab along the vaginal wall and then placing the swab in 1 ml RNALater (Life Technologies, Grand Island, NY) for elution. After 24 h at 4°C, the supernatant fraction of the vaginal fluid was frozen at −70°C for subsequent viral load analysis. During menses, genital samples were not collected. Menstrual cycle characteristics were based on observational menses logs and reverse cycle calculations. SIV inoculation was conducted during the secretory phase of the menstrual cycle in early spring. Plasma progesterone levels were evaluated at the time of genital sampling using a radioimmunoassay kit, Coat-A-Count Progesterone (Seimens Healthcare Diagnostics Inc., Tarrytown, NY), according to the manufacturer's protocol. The animals were humanely euthanized when they reached predetermined end-stage criteria as previously defined. 31
SIV quantification
Viral loads in plasma, vaginal fluid, and gut tissue were determined by real-time quantitative polymerase chain reaction assay (RT-qPCR) that targets a highly conserved region of SIV gag with a sensitivity of 50 copies SIV/ml of plasma or vaginal fluid and 50 copies SIV/μg mRNA as previously described. 32 Briefly, RNA isolation was performed using Trizol reagent (Life Technologies, Grand Island, NY) according to the manufacturer's protocol and assayed in duplicate RT-qPCR reactions. For statistical comparisons, viral loads were log10-transformed and samples below the limit of quantification were assigned the midpoint value between zero and the lower limit (25 copies).
Lymphocyte quantification and assessment
Complete and differential blood counts were performed at the Tulane National Primate Research Center (TNPRC) using a Siemens Advia 120 Hematology System for total leukocyte counts and Wright–Giemsa staining of blood smears for leukocyte differentials. Lymphocyte subpopulations from whole blood and duodenal tissue were enumerated using flow cytometric analysis as detailed previously. 32,44 The fluorochrome-conjugated antibodies included Alexa Fluor 700 anti-CD3, AmCyan anti-CD8, APC-Cy7 anti-CD20, PerCP-Cy 5.5 anti-CD28, PE-Cy 5 anti-CD95, PE-Cy 7 anti-CD184, and APC anti-CD195 (BD Biosciences, San Jose, CA) and Pacific blue anti-CD4 (eBioscience, San Diego, CA). Analysis was conducted on a BD LSRII flow cytometer (BD Biosciences, San Jose, CA) at the LSUHSC-NO Comprehensive Alcohol Research Center Core Laboratory.
Statistical analysis
All data are presented as mean±standard error of the mean (SEM) for both the VEH/SIV+ and THC/SIV+ groups, unless otherwise indicated. Survival analysis was performed using log-rank statistics in Prism 5 (GraphPad Software, Inc., La Jolla, CA). Two-way repeated measures ANOVA tests were used to analyze viral levels, CBC, and changes in body weight, with chronic treatment (THC vs. VEH) and time serving as factors. Bonferroni multiple comparison tests were performed when significant (p<0.05) interactions were identified. Lymphocyte subsets were analyzed as percent change from baseline with PROC MIXED in SAS (version 9.3) in addition to the Kenward–Roger approximation for denominator degrees of freedom in test statistics. Mann–Whitney t-tests were performed when assessing treatment effects alone with significance set at p≤0.05.
Results
Viral load
Over the course of SIV infection, viral levels did not significantly differ between groups (Fig. 1A). Mean viral load at 7 days post-infection (0.25 months) averaged 6.09±0.22 log copies/ml in the VEH/SIV+ group and 5.79±0.17 log copies/ml in the THC/SIV+ group. At viral set point (2 months post-SIV infection), mean viral loads were slightly higher in the THC/SIV+ animals as compared to the VEH/SIV+ group (5.82±0.37 vs. 5.65±0.25 log copies/ml, respectively). Throughout the remainder of the course of infection, mean plasma viral loads were generally higher in the THC/SIV+ animals as compared to the VEH/SIV+ animals (p=NS).
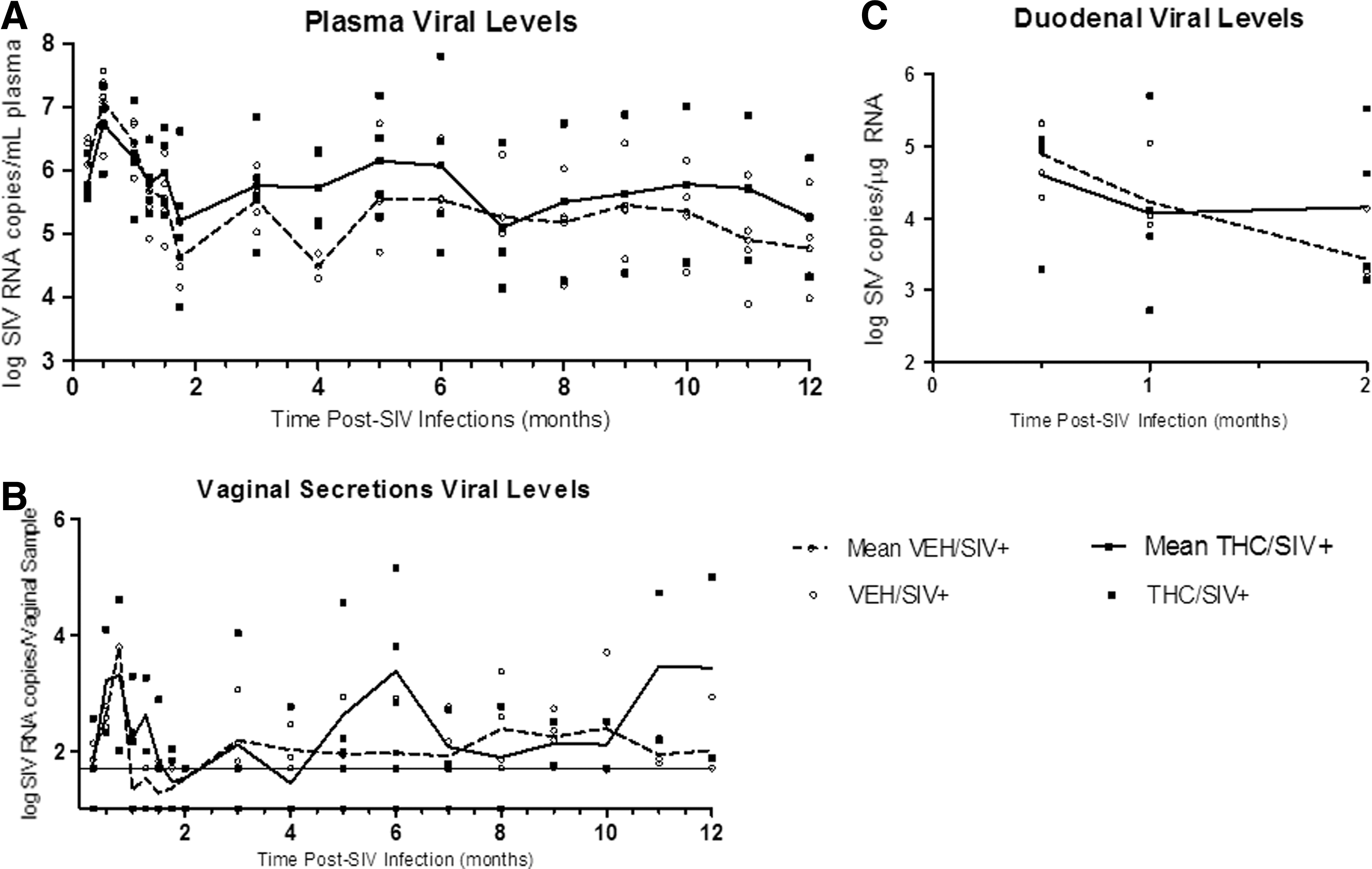
Viral loads detected in
Similarly, vaginal viral loads did not significantly differ between groups over the course of the study, with an average viral level in the VEH/SIV+ group of 2.08±0.12 log copies/vaginal sample and 2.43±0.15 log copies/vaginal sample in the THC/SIV+ group (Fig. 1B). Overall, vaginal viral loads were highly variable over the course of infection, irrespective of treatment. This variability was also reflected in the fact that virus was detectable in only approximately 60% of the samples, and the frequency of viral shedding was not affected by chronic Δ9-THC administration. To examine the levels of vaginal SIV with respect to the menstrual cycle, the relationship between plasma progesterone levels and vaginal viral loads was determined at the time of genital sampling. There was no direct correlation between these two variables (data not shown).
In the gut, an early site of viral replication and potential viral reservoir, viral levels in duodenal biopsies did not differ significantly during the acute phase of the infection (4.34±0.26 and 4.56±0.33 log SIV copies/μg RNA for VEH/SIV+ and THC/SIV+ groups, respectively) (Fig. 1C). The similarity of the viral levels for these two groups over time was also verified statistically in that the two-way ANOVA revealed only a main effect of time (i.e., the effect of treatment was not significant and the interaction of treatment and time was not significant).
Menstrual cycle
Menstrual cycle characteristics were evaluated throughout the study, beginning with the initiation of THC or VEH treatment (Fig. 2). The mean cycle lengths, which were 33.84±3.4 and 34.72±3.2 days for the VEH/SIV+ and THC/SIV+ groups, respectively, were not significantly different. In contrast to the effects on length of the menstrual cycle, the Δ9-THC group had significantly longer menses (5.08±0.4 days) compared to the vehicle group (3.05±0.2 days) (p<0.001). The limited study period prior to SIV inoculation prevented analysis of the effects of THC alone.
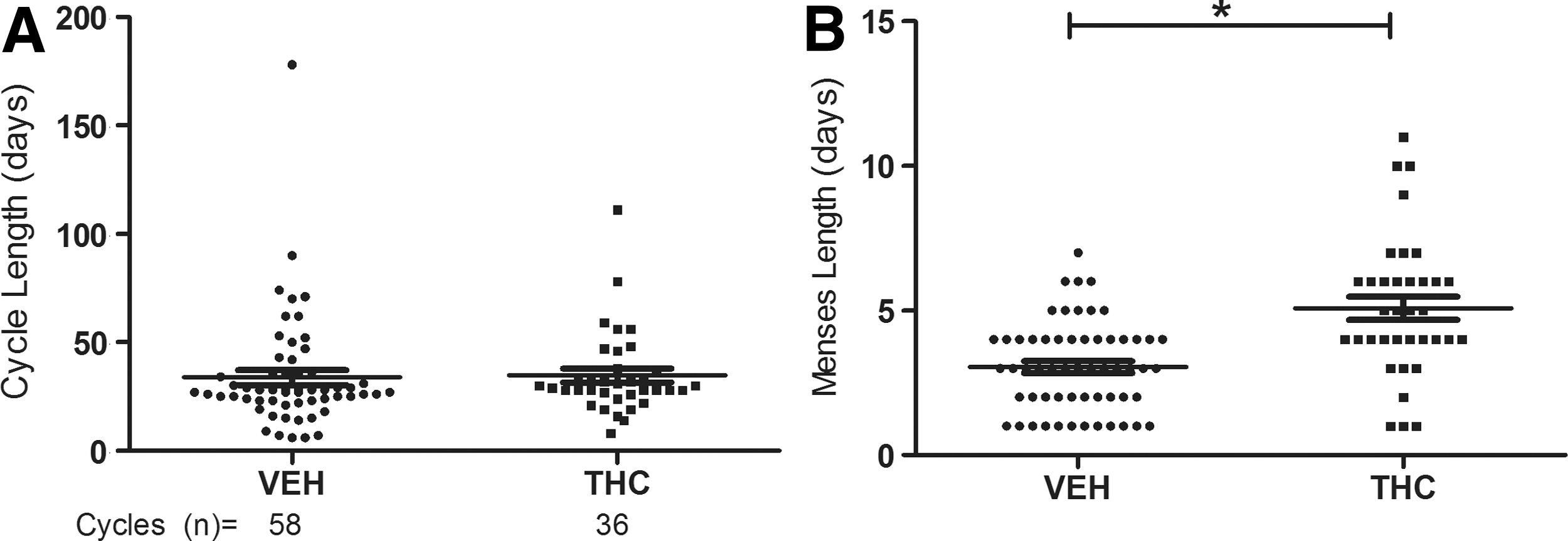
Observations of menstrual cycle in VEH (n=4) and THC (n=4) animals for 58 and 36 cycles, respectively. Observations began at the initiation of VEH or THC treatment and continued throughout the SIV infection period. Daily observation of vaginal bleeding and reverse cycling calculations were used to determine
Body weight change
After 28 days of treatment, a mean loss in body weight was observed in the Δ9-THC animals, as compared to the VEH animals. Over the first 6 months of SIV infection, this trend continued and significant differences in body weight between groups were noted (Fig. 3). During this time period, VEH/SIV+ animals gained weight; in contrast, the THC/SIV+ animals had little or no weight gain. This difference in weight gain was statistically significant after 4 months of SIV infection (p=0.0019). As shown by the plot for the cumulative weight change in Fig. 3 (see inset), the VEH/SIV+ group gained an average of 0.3475±0.12 kg while the THC/SIV+ group lost an average of 0.1041±0.06 kg over the same 6 months of SIV infection (p=0.0052).

Body weight measurements were recorded from individual animals during weekly physical examinations pre-SIV and post-SIV inoculation. The mean weight changes (kg) from basal to 6 months post-SIV infection are shown for individual animals (symbol) and treatment group (lines). Significant changes in body weight were observed between experimental groups after 4 months of SIV infection; *p<0.05, # p<0.001, two-way repeated measures ANOVA was used to analyze the change in body weight over time and chronic treatment. Inset represents the cumulative mean weight change±SEM over the initial 6 months post-SIV infection (VEH, white bar; THC, black bar). Treatment effect was assessed using the Mann–Whitney t-test, p=0.0052.
Lymphocyte population
Prior to SIV inoculation, the THC-treated animals had lower absolute counts of both CD4+ and CD8+ T lymphocytes, but a comparable CD4+ to CD8+ ratio to that of VEH-treated animals (1.87 vs. 1.93) (Fig. 4). Further assessment of the lymphocyte subsets in peripheral blood indicated no significant effect of chronic Δ9-THC administration prior to SIV inoculation on either central (CD28+CD95+) or effector (CD28−CD95+) memory T cell populations. However, total CXCR4 expression in peripheral blood was significantly different between the experimental groups, as the THC-treated animals had a higher percentage of CD4+ and CD8+ cells expressing this receptor (Table 1). The increased frequency of CXCR4 was also seen in the duodenal CD4+ and CD8+ lymphocytes where the THC-treated animals had a lower incidence of CD8+ cells compared to controls, as well as significant differences in CD8+ memory subsets (Table 1).

Analysis of peripheral blood T cell populations in VEH/SIV+ and THC/SIV+ groups before and after SIV inoculation.
Peripheral blood and duodenal lymphocyte subpopulations, memory subsets, and chemokine receptor expression within cells isolated from the VEH/SIV+ and THC/SIV+ groups, both pre-SIV and at necropsy. Values (percent of the parent population) are presented as mean±SEM for VEH (N=4) and THC (N=4 pre-SIV, N=3 at necropsy) animals.
SIV, simian immunodeficiency virus; VEH, vehicle; THC, Δ9-tetrahydrocannabinol.
Following SIV inoculation, a similar decline in the CD4+ to CD8+ ratio was observed in both experimental groups. With respect to the CD4+ and CD8+ lymphocyte memory subsets and chemokine receptor expression, there was also no difference between treatment groups over the course of SIV infection (data not shown). At necropsy, animals in the THC/SIV+ group had reduced percentages of peripheral CD8+ central memory cells and higher duodenal CD4+ memory subpopulations. No significant alterations in the frequency of CXCR4 and CCR5 receptors were noted at necropsy in either group (Table 1).
Survival and end-stage criteria
The first animal to reach end-stage criteria was from the THC/SIV+ group at approximately 6 months post-SIV inoculation (Fig. 5). Two additional THC/SIV+ animals met end-stage criteria before the first subject in the VEH/SIV+ group at 13 months postinoculation. The median survival time of the THC/SIV+ group was approximately 9.6 months. Differences in survival rates were not statistically significant based on the Kaplan–Meier survival analysis (log rank, p=0.1005). Although 75% of the THC/SIV+ group succumbed by study end, SIV levels at that time suggest the death of one animal (ID# 194) was unrelated to SIV infection. Markers of SIV disease at necropsy and clinical indications for euthanasia are listed in Table 2 for individual animals.

Percent survival of VEH/SIV+ (n=4) and THC/SIV+ (n=4) female rhesus macaques over the months following SIV inoculation. Log rank (Mantel–Cox) statistics for the survival curve indicate no significant difference in mortality between experimental groups (p=0.1005).
Discussion
This study evaluated the impact of chronic Δ9-THC administration on markers of SIV disease progression before and after SIV infection in female rhesus macaques. The purpose was to compare the response with the protective effects previously shown in male rhesus macaques using a similar analysis of clinical parameters. In contrast to the protective effects observed in our previous study with males, chronic Δ9-THC administration did not protect female macaques from early mortality induced by SIV infection. 31 Three of four females administered chronic Δ9-THC in this study met end-stage SIV disease criteria, while none of the four males treated similarly with THC progressed to end stage. Moreover, chronic Δ9-THC did not attenuate viral loads in the plasma, gut mucosa, or vaginal secretions of SIV-infected female macaques compared to those of VEH/SIV+ females. With respect to body weight, female macaques that received chronic Δ9-THC showed attenuated weight gain compared to that of vehicle-treated animals. 15,30,31 In fact, the THC/SIV+ females had a net loss in body weight over the study period, relative to VEH/SIV+ animals.
Marijuana use is highly prevalent within the HIV-infected population and is commonly used by HIV-infected women as observed in the Women's Interagency HIV Study (WIHS). 2,3,46,47 However, neither clinical nor preclinical studies have directly examined whether cannabinoids can affect disease progression in women. This question is even more pertinent given the mounting evidence indicating that cannabinoids exert both sex-dependent behavioral and biological effects. 36,48,49 For example, men have frequently reported greater subjective effects (e.g., feeling high) from Δ9-THC than women, whereas women are more likely to report dizziness than men at similar ratings of intoxication from THC and marijuana. 50 –53 These reports are consistent with studies showing that the cannabinoids produce sexually dimorphic effects on antinociception, motor activity, catalepsy, and anxiety levels. 49,54 –56
Sex hormones, particularly estrogens, have been shown to antagonize cannabinoid-mediated effects and potentially underlie their sexually dimorphic effects. For example, estrogens have been shown to decrease CB1 receptor density, cochaperones, ligand affinity, signal transduction, and transcription in distinct regions of the rat brain. 38,39,41 –43 These effects, in combination with the capacity of ovarian hormones to regulate the immune response, could help explain the lack of protection seen in females with SIV disease and our sexually dimorphic findings between male and female macaques. However, if sex hormones can antagonize cannabinoid receptor-mediated effects, it is possible that higher THC doses could produce the same “protective” effects observed in SIV-infected male macaques. In other words, if estrogen only shifts the dose-effect curves for agonists of the cannabinoid receptor system rightward in a parallel manner, higher doses should provide the same protection in females as observed in males. This needs to be determined with further study, as does the bidirectional interaction between the endocannabinoids and gonadal hormones. 57 If both endogenous and exogenous cannabinoids can downregulate the hypothalamic–pituitary–gonadal activity, then higher cannabinoid doses may not be an option due to their adverse effects on sexual function and behavior. 58,59
Cannabinoids are used therapeutically to treat HIV wasting syndrome, as Δ9-THC influences orexigenic activity through interactions with appetite-related neuropeptides in the hypothalamus. 60,61 Previous studies have shown weight gain or maintenance with cannabinoid use in HIV-infected individuals, as well as improvement in appetite. 10,11,62 However, women represented only 7–10% of the study participants and an analysis based on sex was not performed. Changes in body composition due to HIV wasting have already been noted to be sex specific; women lose more body fat relative to lean body mass with the opposite occurring in men. 7,63,64 Furthermore, males have been shown to be more sensitive to CB1 receptor agonist/antagonist effects concerning food intake and body weight relative to females. 16,65 Taken together with estrogens' effects on appetite, the difference in weight gain between SIV-infected male and female macaques chronically administered Δ9-THC is not particularly surprising. 66,67 Furthermore, the female macaques were at a stage of their growth curve where weight gain was expected despite SIV infection, as observed in the VEH/SIV+ group. The absence of change in body weight suggests a potential detrimental effect of chronic Δ9-THC use on growth and development during late adolescence and early adulthood in females.
The observed increase of CXCR4 receptor expression on peripheral T lymphocytes prior to SIV inoculation was consistent with that seen in male macaques chronically administered Δ9-THC. 32,44 The major ligand for CXCR4 receptors, stromal-derived factor-1 (SDF-1), has been shown to play a role in the migration of T cells and an increase in expression could interact with SIV infection in this manner. However, the importance of this increase is not related to the capacity of the CXCR4 receptor to serve as a coreceptor for HIV infection of CD4+ T lymphocytes, because SIVmac251 does not use this site for cell entry. 68,69 Interpreting the contribution of these increases is also complicated by the fact that the increased CXCR4 expression occurred in both sexes, but was associated with the attenuation of disease only in males. Finally, despite the changes in receptor expression prior to SIV inoculation, higher levels of peripheral or duodenal lymphocytes in the Δ9-THC female animals were not observed as compared to vehicle animals. 70
The results from this study involving female rhesus macaques infected with SIV suggest that chronic Δ9-THC produces sex-specific differences on protection from early mortality, which may be relevant to HIV-infected women. Further studies are needed to explore the sex-dependent effects of cannabinoids on HIV disease progression as well as viral transmission, as our studies show that specific clinical markers of SIV disease progression were not affected by chronic Δ9-THC administration. Moreover, this study had several limitations, including the small number of animals, the evaluation of only a single dose of Δ9-THC, the absence of a Δ9-THC SIV-negative control group, and incomplete examination of gonadal hormone patterns throughout the study. Additional studies are warranted to further explore these factors and to evaluate both protective and permissive factors associated with HIV acquisition and transmission.
Footnotes
Acknowledgments
The authors would like to thank Constance Porretta for her outstanding analytical contributions. We would also like to thank Jamie A. Hubbell and Peter B. Lewis for their technical assistance, and Dr. Lynn LaMotte for his statistical contributions. Supported by NIH/NIDA R01 DA020419-05, NIDA R01 DA020419-04S3, NIDA R01 DA030053-04, and NIAAA T32 AA007577-15.
Parts of the data were presented at the Symposium on Nonhuman Primate Models for AIDS (Atlanta, October 2013) and the Society of Neuroimmune Pharmacology 20th Scientific Conference (New Orleans, March 2014).
Author Disclosure Statement
No competing financial interests exist.