
Abstract
Previously, we had shown that although only 8% of patients with large granular lymphocytic leukemia (LGLL) were infected with human T cell lymphoma/leukemia virus (HTLV)-2, almost half had antibodies to HTLV Gag and Env peptides. Herein, we investigated whether this could be due to cross-reactive antibodies to two homologous peptides in the Gag protein of the endogenous retrovirus HTLV-related endogenous sequence-1 (HRES-1). In addition, we had previously shown that patients with HTLV neurodegenerative diseases had increased seroreactivity to homologous HERV-K10 endogenous retrovirus peptides. Hence, in this study we also examined whether these patients had increased seroreactivity to the aforementioned HRES-1 Gag peptides. Sera from 100 volunteer blood donors (VBD), 53 patients with LGLL, 74 subjects with HTLV-1 or 2 infection (58 nonmyelopathy and 16 myelopathy), and 83 patients with multiple sclerosis (MS) were evaluated. The HTLV-positive myelopathy (HAM) patients had a statistically increased prevalence of antibodies to both HRES-1 Gag peptides (81%) vs. the VBD (0%), LGLL patients (13%), and MS patients (1%), and the HTLV-positive nonmyelopathy subjects (21%). The data suggest that cross-reactivity to HRES-1 peptides could be involved in the pathogenesis of HAM. The difference between the VBD and LGLL patients was also statistically significant, also suggesting a possible association in a minority of patients.
Introduction
H
In a subsequent study using the same VBD and LGLL patient cohorts but a different HTLV-1-infected or HTLV-2-infected population, we tested for seroreactivity to this same HERV-K10 Pol peptide and an HERV-K10 Gag peptide that was homologous to HTLV p24 Gag. 9 Although there was a slight, statistically significant increased prevalence of anti-HERV-K10 Pol peptide antibodies in the LGLL group vs. the VBD, this was not the case for anti-HERV-K10 Gag reactivity. The more interesting observations were that the HTLV-infected myelopathy patients had statistically significant increased seroreactivities to both HERV-K10 peptides vs. the VBD or HTLV nonmyelopathy patients. Given that the HTLV Tax protein can transactivate the expression of HERV-K10 in human tissues, the above data suggested that immuno-cross-reactivity to HERV-K10 peptides may be involved in the pathogenesis of HTLV myelopathy. 10
To investigate other reasons for the observed anti-HTLV p24 Gag reactivity in LGLL patients, and whether cross-reactive immunity against other endogenous retroviral sequences could be involved in the pathogenesis of HTLV-myelopathy (HAM), we tested the above subject populations for antibodies to HTLV Gag homologous peptides present in HTLV-related endogenous sequence-1 (HRES-1).
HRES-1 is an incomplete endogenous retroviral sequence that entered the primate genome many years ago. 11 It is located on chromosome 1q42. 12 It exists as either of three distinct alleles and is polymorphic among humans. 12,13 There are three different sequences published for HRES-1, each derived from different cultured and uncultured human lymphocytes. 11,14 –16 One of these sequences encodes for overlapping putative p25 and p15 Gag proteins, while the other two, because of a stop codon, do not; but they do encode for three putative products of 8 (p8), 15 (p15), and 5 (p5) kDa. The p25 contains two peptides, which are also in p8 and p15, respectively, that have homology to HTLV-I and II p24 Gag peptides (Fig. 1). Previous studies have shown immuno-cross-reactivity between antibodies against HTLV p24 Gag proteins and HRES-1 Gag peptides, and their possible association with autoimmune diseases. 16,17

Alignment of the amino acids from human T cell lymphoma/leukemia virus-related endogenous sequence-1 (HRES-1) Gag peptide 14–24 (top) and 117–127 (bottom) against homologous sequences in human T cell lymphoma/leukemia virus (HTLV)-II and HTLV-II p24 Gag.
Materials and Methods
Subjects
In an IRB-approved archival study, serum and peripheral blood mononuclear cells (PBMCs) were obtained from 100 VBD, 53 LGLL patients, 74 HTLV-1 or HTLV-2-infected individuals (HTLVt), 16 of whom had myelopathy (HTLVm) (eight HTLV-1 and eight HTLV-2), and 58 who did not (HTLVn) (22 HTLV-1 and 36 HTLV-2), and 83 HTLV-negative multiple sclerosis (MS) patients. All LGLL patients had clonal T cell lymphocytoses, as confirmed by polymerase chain reaction (PCR) for clonal rearrangements of the T cell receptor γ-gene and by positivity for CD3, CD8, and CD57 proteins. 18 The HTLV myelopathy patients were all positive for either HTLV-1 or HTLV-2 antibodies and DNA in both their peripheral blood and cerebrospinal fluid. They all manifested signs and symptoms of upper motor neuron disease, and met the updated WHO criteria for HTLV myelopathy. 19,20 The MS patients were negative for HTLV-1 and HTLV-2 by serologic and nucleic acid studies and fulfilled the McDonald diagnostic criteria, which utilize both clinical and magnetic resonance imaging (MRI) findings. 21
Serologic studies
All subjects were screened for anti-HTLV antibodies using a commercial HTLV1/2 EIA assay, as previously described. 22 Seropositive individuals were further evaluated using an HTLV-1/2 Western blot, as previously described. 23 Sera were also tested for antibodies against the HRES-1 peptide PTRAPSGPRPP Gag (14–24) and RREGPDRSPR Gag (117–127), which are homologous to HTLV-I and HTLV-II Gag p24 (Fig. 1). Briefly, the peptides were synthesized (New England Peptide LLC, Gardner, MA), and the ELISA was performed as follows. The HRES-1 peptides were diluted in carbonate buffer, pH 9.6 (Sigma, St. Louis, MO) and 50 μl containing 0.5 μg was added to each well and incubated overnight at 4°C. The wells were then washed five times with wash buffer [1×phosphate-buffered saline (PBS) plus 0.2% Tween 20], and 100 μl of superblock blocking buffer (Thermo Scientific, Hudson, NH) was added to each well, incubated for 1 h at 37°C, and washed five times with wash buffer. Then, 50 μl of the primary antibody sample at the desired dilution in diluent buffer was added to the well. The samples were incubated for 1 h at 37°C and removed, then washed five times with wash buffer. Goat antihuman IgG-HRP (1:2,000 in diluent buffer) (Sigma, St. Louis, MO) was added and incubated for 1 h at 37°C. Plates were then washed five times, and 50 μl of hydrogen peroxide and tetramethylbenzidine was added to each well and incubated for 10 min at room temperature in the dark. The reaction was stopped by the addition of 50 μl 1N H2SO4 to each well and the optical densities of the samples were read at 450 nm. A sample was deemed positive if, at a dilution of 1:200, its optical density was ≥2.5×background. Positive samples were then serially diluted and retested to determine their end point titer.
PCR studies
DNA was extracted from serum or PBMCs, and DNA integrity was confirmed by β-globin PCR, as previously described. 24 Only samples that tested positive for human β-globin at 0.1 μg and 1 μg of DNA input were deemed suitable for subsequent retroviral DNA analysis. DNA was analyzed for HTLV-1 and HTLV-2 DNA using the pol primers SK110/SK111, the HTLV-1 pol probe SK-112, and the HTLV-2 pol probe SK188, as previously described. 24
Sequencing analyses
DNA from the PBMCs of four VBD and four HTLVm patients was amplified with the HRES-1 gag primers H2, L3, HS, and HAS, as previously described, 16 except that the following cycling conditions were used: 94°C, 1 min; 55°C, 45 s; 68°C, 45 s, for 40 cycles, with a final elongation step at 68°C for 5 min. Amplified DNA was directly sequenced, as previously described. 25 The new sequences were aligned with the previously published sequences using NCBI-BLAST. 26
Statistics
Seroprevalence rates between various subject populations were analyzed using Fisher's t-test. 27 Comparisons of serologic optical density scores and antibody titers were analyzed using the contingency table method. 27
Results
The HRES-1 gag DNA sequences from four VBD and four HTLVm patients were identical (GenBank accession #s KJ596646–KJ596653). They do not encode for a p25 peptide, but rather p8 and p15 peptides (Fig. 2). Hence, they are in agreement with three out of four previous publications describing the HRES-1 sequence. The HRES-1 Gag (14–24) and Gag (117–127) peptide sequences were conserved in all eight sequences and there were no differences in the deduced amino acid sequence among the VBD or HTLVm patients.

Alignment of deduced amino acid sequences from amplified HRES-1 gag DNA from various sources including four volunteer blood donors (VBD) and four patients with HTLVm and three previously published sequences (GenBank accession #s KJ596646–KJ596653, XI6660, and AF216972) and those published by Rasmussen et al.
13
Those for HRES-1 p8 Gag

Table 1 shows the prevalence rates for anti-HRES-1 Gag (14–24) and HRES-1 Gag (117–127) peptides antibodies among the various subject populations. Antibodies to HRES-1 Gag (117–127) tended to be more prevalent. Compared to the VBD (8%), there was a statistically higher prevalence for antibodies to HRES-1 Gag (14–24) in the HTLVt (54%), HTLVm (81%), and HTLVn (47%) populations. The HTLVm patients had a statistically higher prevalence rate than the HTLVn subjects. For the HRES-1 peptide (117–127), the LGLL (81%), HTLVt (59%), HTLVm (100%), HTLVn (48%), and MS (61%) all had statistically significant higher prevalence rates than the VBD (8%) subjects. Again, the seroprevalence rate in the HTLVm patients was statistically significantly higher than the HTLVn subjects. These relationships held true when either optical density (data not shown) or antibody titer was examined (Figs. 3 and 4).
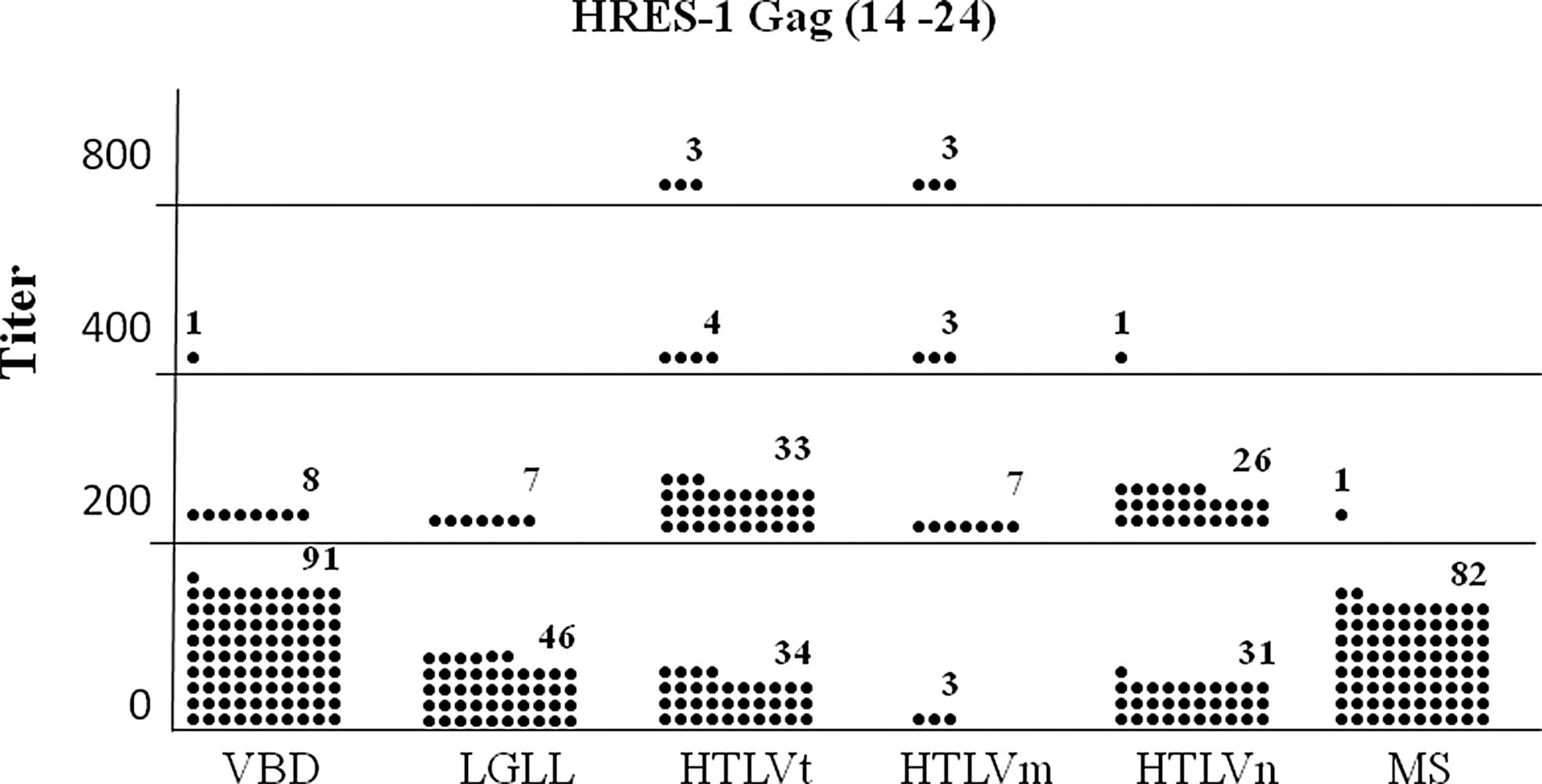
Distribution of seropositive titers for the anti-HRES-1 Gag (14–24) peptide ELISA antibody assay among the various subject populations. Each dot represents one subject. The means for VBD, large granular lymphocytic leukemia (LGLL), HTLVt, HTLVm, HTLVn, and MS subjects were 20, 26, 181, 488, 97, and 2, respectively. VBD vs. LGLL, p=0.459; VBD vs. HTLVt, p≤0.001; VBD vs. HTLVm, p≤0.001; VBD vs. HTLVn, p≤0.001; VBD vs. MS, p=0.068; HTLVm vs. HTLVn, p≤0.001.
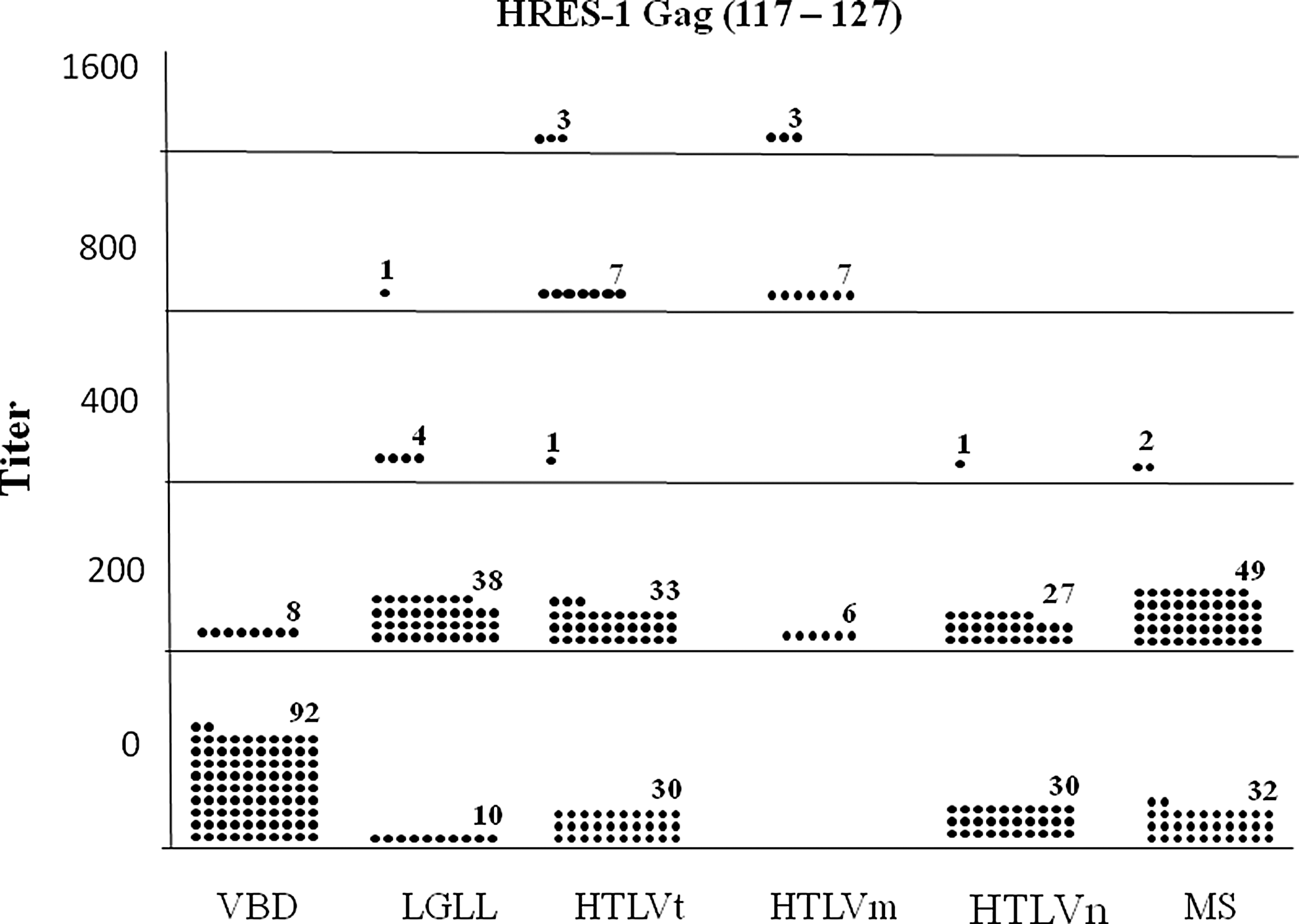
Distribution of seropositive titers for the anti-HRES-1 Gag (117–127) peptide ELISA antibody assay among various subject populations. Each dot represents one subject. The means for VBD, LGLL, HTLVt, HTLVm, HTLVn, and MS subjects were 16, 188, 230, 725, 100, and 125, respectively. VBD vs. LGLL, p≤0.001; VBD vs. HTLVt, p≤0.001; VBD vs. HTLVm, p≤0.001; VBD vs. HTLVn, p≤0.001; VBD vs. MS, p≤0.001; HTLVm vs. HTLVn, p≤0.001.
HRES-1 14–27: VBD vs. LGLL, p=0.225; VBD vs. HTLVt, p<0.001; VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p<0.001; VBD vs. MS, p=0.033; HTLVm vs. HTLVn, p<0.013.
HRES-1 117–127: VBD vs. LGLL, p=0.001; VBD vs. HTLVt, p<0.001; VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p<0.001; VBD vs. MS, p<0.001; HTLVm vs. HTLVn, p<0.001.
Both: VBD vs. LGLL, p=0.001; VBD vs. HTLVt, p<0.001; VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p<0.001; VBD vs. MS, p=0.454; HTLVm vs. HTLVn, p<0.001.
VBD, volunteer blood donor; LGLL, large granular lymphocytic leukemia; HTLV, human T cell lymphoma/leukemia virus; MS, multiple sclerosis; HRES-1, HTLV- related endogenous seguence-1.
When antibody positivity to both HRES-1 peptides is considered, the LGLL (13%), HTLVt (34%), HTLVm (81%), and HTLVn (21%) subjects all had statistically significant higher prevalence rates than the VBD (0%), while the MS (1%) patients did not (Table 1). Again, the HTLVm patients had a statistically significant higher prevalence rate than the HTLVn subjects. Of the 16 HTLV myelopathy patients, eight were HTLV-1 positive and eight were HTLV-2 positive. All 16 were positive for HRES-1 (117–127) antibodies, while six of the HTLV-1-infected and seven of the HTLV-2-infected patients were positive for HRES-1 (14–24) antibodies. Obviously, there were no statistically significant differences in these seroprevalence rates between the HTLV-1-infected and HTLV-2-infected patients. Likewise, among the HTLVn subjects, there was no statistically significant difference between HTLV-1-positive and HTLV-2-positive subjects' reactivity to the HRES-1 peptides. Furthermore, there were no statistically significant differences in anti-HRES-1 peptide OD and titers between HTLV-1-infected and HTLV-2-infected individuals.
Table 2 shows the data regarding positive antibodies to either HERV K-10 Gag and Pol peptides and/or HRES-1 Gag peptides. As can be seen, the HTLVm and HTLVn patients had a statistically significant greater reactivity to a higher number of the four endogenous retroviral peptides compared to the VBD, while the HTLVm had statistically significant greater reactivities than the HTLVn patients.
VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p<0.001; HTLVm vs. HTLVn, p=0.041 for 0 out of four reactivities. VBD vs. HTLVm, p=0.032; VBD vs. HTLVn, p<0.001; HTLVm vs. HTLVn, p<0.001 for one out of four reactivities. VBD vs. HTLVm, p=0.258; VBD vs. HTLVn, p<0.001; HTLVm vs. HTLVn, p=0.254 for two out of four reactivities. VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p=0.048; HTLVm vs. HTLVn, p<0.035 for three out of four reactivities. VBD vs. HTLVm, p<0.001; VBD vs. HTLVn, p=0.017; HTLVm vs. HTLVn, p<0.001 for four out of four reactivities.
Discussion
An increased risk for developing HAM is associated with blood-borne viral transmission, increased proviral DNA loads and anti-HTLV antibodies, polymorphisms in the interleukin (IL)-10 promoter and IL-28B gene, HLA–A* 02
It has been estimated that 8% of the human genome is composed of endogenous retroviral elements. 37 Most of these are defective and transcriptionally inactive, but many are capable of RNA and protein expression. 38 –40 If expressed prior to the ontogeny of the host immune system, they would be recognized as “self”; if subsequent to the formation of a competent immune system, they would be recognized as “foreign” and a potential cause of autoimmune diseases. 40,41
HERV-K10 is a member of the HML-2 class of endogenous retroviruses. 42 It has been shown to be expressed in numerous human tissues including the central nervous system, and it has been a suggested target of autoimmune responses. 43,44 Such a hypothesis is consistent with observations that increased expression of HERV-K in HIV-1-infected patients leads to a cytotoxic T cell response toward HERV proteins. 45 –47 Expression of HERV-K proteins and immune responses thereto have been associated with a variety of human malignancies and autoimmune diseases. 48 –50 Furthermore, the HTLV-1 Tax protein has been shown to transactivate HERV-K LTR controlled transcription. 10
HRES-1 is an incomplete human retrovirus that is distinct unto itself compared to the three main classes of human endogenous retroviruses. 51,52 It entered the primate genome millions of years ago after they had separated from other mammalian groups. 11 It has been shown to be transcriptionally expressed in both human lymphocytes and brain cells. 17,53 Some controversy exists as to the exact sequence of HRES-1. The initial sequence derived from cultured lymphocytes from a patient with cryoglobulinemia contained an open reading frame that encoded a putative p28-kDa protein, which was shown to be expressed in H9 cells, a cell line derived from human malignant CD4 cells. Expression of HRES-1 protein(s) was localized to the cytoplasm and to nuclear bodies. 17 Peptides within p28 are homologous to HTLV Gag peptides.
Subsequent sequencing studies, including our own reported herein, indicate that a stop codon would prevent the translation of the p28 kDa, and rather encode for two smaller proteins, p8 and p15, which contain the aforementioned peptides homologous to HTLV-1 p24 Gag peptides. The p8 peptide also contains an additional 24 aa sequence homologous to the HTLV-1 p19 Gag protein, reactivity to which we have yet to analyze. 16 Using antibodies made against these HRES-1 peptides, some investigators have shown expression of the peptides in human cell lines, while others have not. 16,17,53 Interestingly, two groups have identified an 80-kDa protein recognized by the anti-p117–127 antibodies in cultured human cells. 16,17 It is unclear if this 80-kDa protein is encoded by HRES-1 or is a cellular protein that is immunologically cross-reactive. Several studies have shown increased seroreactivity to these HRES-1 peptides in patients with autoimmune diseases. 14,15,53,54
The hypothesis that the pathogenesis of HTLV-related neurodegenerative diseases could involve an anti-HTLV cross-reactive autoimmune response directed against homologous human endogenous retroviral peptides seems plausible. Our data indicate that patients with either HTLV-I or HTLV-II neurologic diseases have statistically significant increased antibody reactivity to at least four such different peptides encoded by two genetically distinct endogenous retroviruses, HERV K-10 or HRES-1. Obviously, the data presented herein alone would be insufficient to make the case that cross-reactivity to endogenous retroviral peptides is the cause of HTLV neuropathy. Future studies to explore this hypothesis could involve analyses of specific T cell responses to these same peptides and/or prospective or retrospective studies to explore whether immunoreactivity to endogenous retroviral peptides predicts for the development of neurodegenerative diseases in HTLV-infected subjects.
The observation that a statistically significant minority of LGLL patients had increased antibodies to both HRES-1 Gag peptides and a majority of LGLL and MS patients have antibodies to the HERS-1 117–127 peptide is also interesting. It is possible that the pathogenesis of some cases of LGLL could involve chronic activation of LGL by an antigen (including endogenous retrovirus peptides) and subsequent malignant transformation. These observations deserve further study as well.
Footnotes
Acknowledgment
The study was supported by the Barbara Kopp Cancer Research Fund.
Author Disclosure Statement
No competing financial interests exist.