
Abstract
Lentivirus infections are characterized by a dramatic loss of mucosal CD4+ T cells, breakdown of the gut mucosa, and subsequent chronic immune activation. Residual immune activation persists even in patients controlling virus replication and remains a significant source of ongoing disease morbidities, but the causes are unclear. Plasmacytoid dendritic cells (pDCs), primary producers of interferon (IFN)-α, have been previously shown to be depleted from peripheral blood of HIV patients and simian immunodeficiency virus (SIV)-infected macaques, and most recently have been shown to accumulate in the gut mucosa. Although previous work has shown that pDC frequencies can be reduced in the circulation of HIV-1 Elite Controllers, it is unknown if gut-homing also occurs. In this new study we found that during progressive HIV-1 infection pDCs were depleted in peripheral blood compared to seronegative controls, and, correlating with plasma viremia, the remaining pDCs upregulated the gut-homing marker, α4β7. Even in HIV-1 Elite Controllers pDCs were significantly reduced in blood and α4β7 expression was still significantly upregulated compared to seronegative controls. Interestingly, pDC trafficking to the gut was associated with increased Ki67 and HLA-DR on circulating CD4+ and CD8+ T cells. Overall, these data suggest that gut trafficking of pDCs is independent of virus replication and could be mediated by alternative mechanisms, which in turn could contribute to residual immune activation in HIV-1 Elite Controllers.
P
Studies of acute pathogenic lentivirus infection in macaques have demonstrated a transient increase of pDCs in peripheral blood due to rapid egress from the bone marrow. 5,7 pDCs also redistribute to the lymph nodes early after infection where they undergo rapid apoptosis, but are excluded from lymph nodes during chronic infection due to ongoing fibrosis. 7,9,10 Interestingly, others and we have very recently refined this model to show that in fact pDCs are not necessarily depleted by HIV/SIV infection, but can accumulate in the gastrointestinal tract, depending on cell surface α4β7 expression. 11,12 pDCs have also been shown to accumulate in the spleen and lymph nodes in some human subjects, 13,14 suggesting pDCs in the circulation may not necessarily be depleted, but rather are redistributing to other tissues. Interestingly, these phenomena do not seem to occur in the nonpathogenic SIV natural hosts, sooty mangabeys and African green monkeys. 12,15
In this study we evaluated the frequency of pDCs and myeloid dendritic cells (mDCs) in the circulation of HIV-1 Elite Controllers, as well as α4β7 expression on both subsets as a surrogate marker for gut homing. Both the University of Alabama at Birmingham (UAB) and Harvard University IRB reviewed and gave approval for the studies herein. Written, informed consent was obtained from each of the study subjects.
HIV-1 Elite Controllers were defined by stable CD4+ T cell counts and sustained plasma viral loads below the threshold of detection as described previously.
16
The median ages of persons included in this study were 45.4, 34.6, and 46.8 years in seronegative subjects, viremic HIV+ subjects, and Elite Controllers, respectively. Genders were 50%, 80%, and 60% male in the seronegative subjects, viremic HIV+ subjects, and Elite Controllers, respectively. Plasma HIV-1 RNA levels were determined using the Amplicor ultrasensitive HIV-1 monitor assay at UAB hospitals (version 1.5; Roche Diagnostics Systems) and absolute CD4+ T cell counts were determined in UAB diagnostic laboratories as described previously.
17
All Elite Controllers had sustained viral loads of <50 copies/ml, plasma, while viremic HIV+ subjects had a median viral load of 50,800 copies/ml, plasma (range, 22,900 to 341,000 copies). Median CD4+ T cell counts were 781 and 639 in Elite Controllers and viremic HIV+ subjects, respectively. mDCs and pDCs were identified in peripheral blood, distinguishable by mutually exclusive expression of the cell-surface integrin, CD11c, and the IL-3Rα chain, CD123, respectively (Fig. 1A and B). pDCs were further confirmed by expression of blood dendritic cell antigen 2 (BDCA-2). Full phenotypes identified pDCs as CD3−CD14−CD20−HLA-DR+CD123+CD11c−BDCA-2+ and mDCs as CD3−CD14−CD20−HLA-DR+CD123−CD11c+ (the full antibody list is in Supplementary Table S1; Supplementary Data are available online at
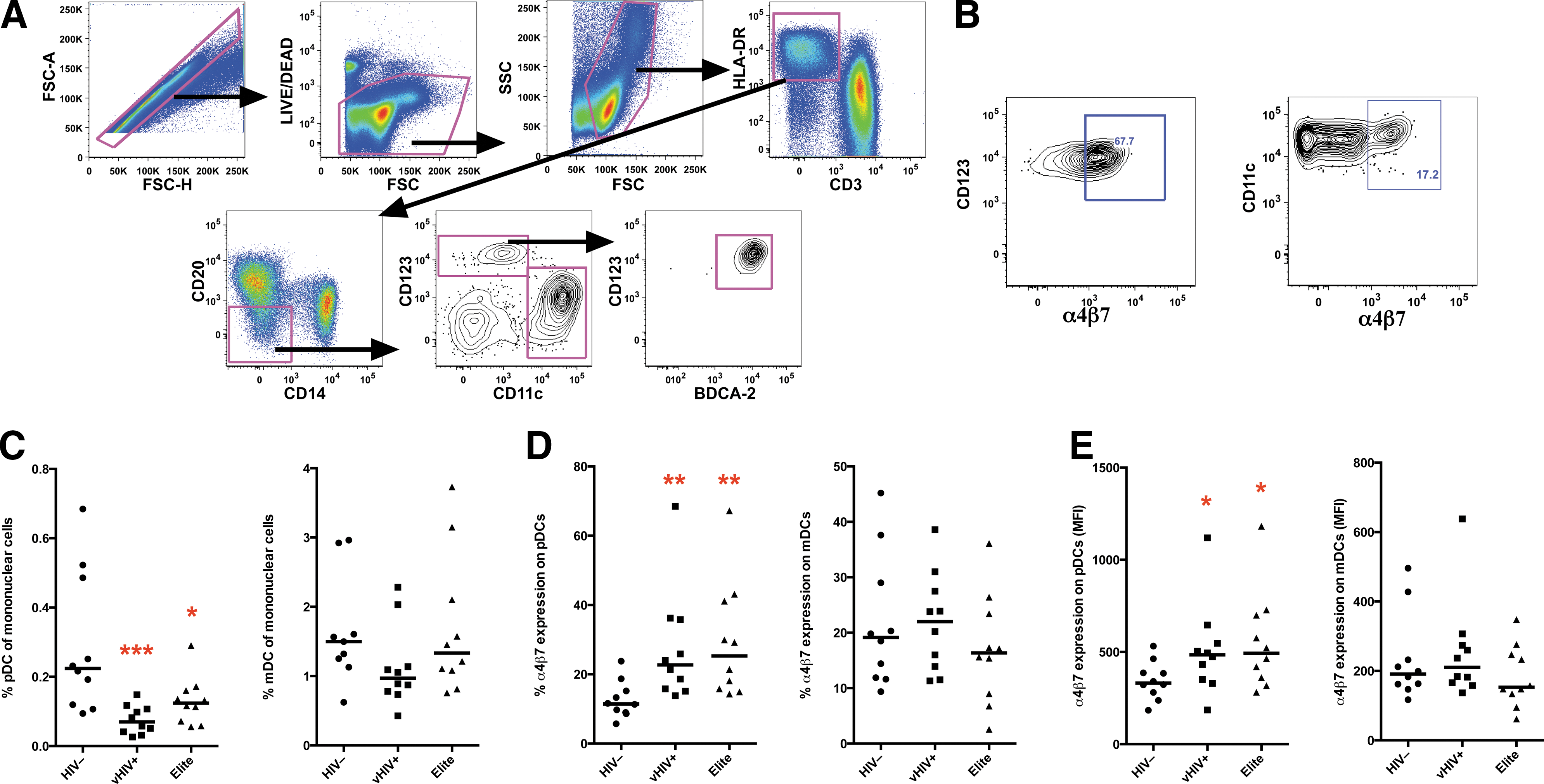
Plasmacytoid dendritic cells (pDCs) but not myeloid dendritic cells (mDCs) are reduced in peripheral blood of HIV-1 viremic patients and Elite Controllers and have a gut-homing phenotype.
Since inflammatory cytokines such as IFN-α can induce apoptosis, generalized immune activation, and may be associated with breakdown of the gut mucosae and microbial translocation, we next evaluated whether measures of pDC gut homing were associated with systemic markers of chronic immune activation. Interestingly, α4β7 expression positively correlated, whereas declining frequencies of circulating pDCs (presumably homing to the gut) negatively correlated with HLA-DR and Ki67 expression in T cells as markers of immune activation (Table 1). No association, however, was found with CD4+ T cell counts or viral load. These data suggest that HIV disease induces gut trafficking of pDCs, which is in turn linked to immune activation and may occur independently of virus replication.
VL, viral load; pDC, plasmacytoid dendritic cell.
Despite its importance in trafficking and maintaining pDCs in the gastrointestinal tract, the mechanism of α4β7 imprinting remains unknown. Whether this occurs in the gastrointestinal tract (GI) or on de novo pDCs developing in the bone marrow is unclear. Recent reports indicate that α4β7 expression is well-correlated with an actual accumulation of the cells in the GI tract in both HIV-infected humans and macaques models, 11,12,18,19 and thus measuring α4β7 expression in blood pDCs could be a useful surrogate biomarker to predict gut homing and potentially ongoing immune activation.
Footnotes
Acknowledgments
This work was supported by an amFAR research grant (108547-53-RGRL) to R.K.R.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.