
Abstract
We previously showed that a matrix (M) gene-deleted rabies virus (RABV)-based vaccine (RABV-ΔM) is highly immunogenic and induces potent B cell responses in the context of RABV infection. We speculated that RABV-ΔM expressing HIV proteins would also induce potent B cell responses against HIV antigens. As a prerequisite to future studies in nonhuman primates, we completed immunogenicity studies in mice to confirm the ability of RABV-ΔM to induce polyfunctional B cell responses in the context of HIV. To that end, the envelope protein from the mac239 strain of SIV (SIVmac239Env) was cloned into RABV-ΔM, resulting in RABV-ΔM-Env. Infectious virus was recovered following standard methods and propagated on baby hamster kidney cells stably expressing RABV M [>107 focus forming units (ffu)/ml]. Western blot analysis of cell lysates or of purified virions confirmed Env expression on the surface of infected cells and within virus particles, respectively. Positive neutralization activity against a neutralization-sensitive SIV strain and to a lesser extent against a neutralization-resistant SIV strain was detected in mice after a single intramuscular inoculation with RABV-ΔM-Env. The quality, but not quantity, of the antibody response was enhanced via boosting with recombinant gp130 or RABV-ΔM-Env as measured by an increase in antibody avidity and a skewing toward a Th1-type antibody response. We also show that an intradermal inoculation induces higher antibodies than an intramuscular or intranasal inoculation. An intradermal inoculation of RABV-ΔM-Env followed by a boost inoculation with recombinant gp130 produced anti-SIV antibodies with neutralizing and nonneutralizing antibody (nNAb) effector functions. Together, RABV-ΔM-Env induces B cells to secrete antibodies against SIV with the potential to clear both “free” and cell-associated virus. Strategies capable of eliciting both NAbs as well as nNAbs might help to improve the efficacy of HIV-1 vaccines.
Introduction
O
Correlates of protection for most vaccines used to prevent human infections are antibodies, most notably IgG. 2 Traditionally, vaccine-induced protection relies on antibodies that prevent or inhibit infection via the antibody variable domain (Fv). Fv binding to epitopes on the surface of microbes prevents infection by many pathogens, including RABV, poliovirus, and smallpox. In the context of HIV infection, passive transfer of NAbs demonstrated the effectiveness of Fv-mediated effector functions in various models of HIV vaccines. 3 –11 Importantly, antibodies that broadly neutralize (bnAbs) a wide range of HIV strains have been identified in patients during the course of natural infection, 12 –18 although the generation of these antibodies most often occurs too late in infection to prevent disease progression. Nonetheless, the isolation of monoclonal bnAbs has greatly increased our understanding of how antibodies can neutralize HIV, including key epitopes to target for vaccination. 19 –25 Despite evidence suggesting that NAbs will play an important role in HIV vaccination, no vaccine has been able to prevent infection through the induction of Fv-mediated NAbs, suggesting that other antibody effector functions might be beneficial.
In recent years, interest in identifying additional antibody effector functions that increase vaccine-induced efficacy against HIV has mounted. The antibody Fc fragment binds to activating or inhibitory Fc receptors (FcRs) on the surface of immune cells, resulting in nNAb effector functions, including antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), and the release of inflammatory mediators. 26,27 Each of these mechanisms inhibits viral infection and/or prevents cell-to-cell spread of HIV-1, thereby limiting the production of progeny virus and decreasing viral loads. 1
Experimental evidence suggests that nNAbs are associated with a decline in viral loads during the acute phase of infection thereby influencing disease progression. 4,7,9,14,28 Using Fc variants of the b12 human monoclonal NAb, Hessell et al. 4 showed that FcγRs contributed in part to protection against SHIV challenge. However, the Hessell study used passive transfer of purified monoclonal antibodies with known mutations, leaving unanswered the question of whether active vaccination can induce a polyclonal antibody response with similar functions and outcomes. This question was partially addressed by several recent studies that used HIV-1 gp120, 9 replication-competent adenovirus type 5 host range mutant (ad5hr)-SIV recombinants, 7,14 and the RV144 canarypox/rgp120 human clinical trial. 28 These studies provided evidence to suggest that nNAb antibody effector functions improved vaccination outcome, indicating the need for additional studies of nNAbs in HIV vaccine-induced protection.
Our laboratory has developed replication-deficient RABV-based vaccine vectors that lack one of the RABVs' five essential genes. 29 –34 A matrix (M) gene-deleted RABV, RABV-ΔM, is emerging as our most promising vector in the context of human rabies vaccines. RABV M is critical for RABV assembly and budding. 35 M gene-deleted RABVs lose their characteristic bullet shape and have 10,000-fold reduced infection efficiency compared to the parental strain due to a budding defect. 35 As such, RABV-ΔM contains all of the transcription and replication machinery necessary for gene expression and genome replication, resulting in high antigen expression levels. 29 Importantly, RABV-ΔM induces Th1-type polarized antibodies in mice and nonhuman primates (NHPs). 29 This observation, coupled with the known role of Th1-type antibodies in mediating Fc:FcR antibody effector functions, leads us to believe RABV-ΔM might make an effective HIV-1 vaccine capable of exploiting the effector functions of nNAbs.
Additional features suggest RABV-ΔM might make for an antibody-based HIV vaccine. Foreign glycoproteins can be expressed from recombinant RABV genomes leading to exposure on the surface of infected cells and incorporation into virus particles. 36 –39 This exposes antigen to B cells in the context of free and cell-associated virus and might increase the immunogenicity of the antigen. RABV-ΔM induces the development of effective innate, T helper, and B cell responses 32 that are consistent with the development of antibodies with a broad range of nNAb effector functions. RABV-ΔM does not spread to the central nervous system (CNS) of T and B cell-deficient mice and is safe in both mice and NHPs. 29 Importantly, suckling and adult mice inoculated intracranially with M gene-deleted RABVs do not manifest clinical disease 40 (J.P. McGettigan and S. Shen, unpublished). Together, preclinical testing of RABV-ΔM indicates that it is safe when injected peripherally or directly into the CNS. Finally, rabies remains a global health threat in much of the world, especially in Africa and Asia where improved RABV vaccines and the development of an HIV vaccine are critically needed. A bivalent, chimeric RABV-HIV vaccine might help to reduce the burden of two global health threats.
Here we report on the results of prerequisite immunogenicity studies in mice showing that a replication-deficient RABV-based vaccine vector expressing SIV Env induces NAb and nNAb effector functions against SIV, suggesting that a chimeric RABV-HIV vaccine vector might hold promise as an HIV vaccine.
Materials and Methods
Test vaccine construction, recovery, and propagation
Construction of the RABV-ΔM-Env vaccine was performed by fusing the ectodomain and transmembrane of SIVmac239Env (Env) (amino acids 1–714; GenBank M33262.1) to the RABV G cytoplasmic domain (CD) (GenBank M31046.1). The SIV Env gene was removed from the pSPBN-IG-Env 41 with BsiWI and HpaI and then inserted into RABV-ΔM-89.6-RVG (J.P. McGettigan, unpublished), resulting in RABV-ΔM-EnvRABVG, hereafter referred to as RABV-ΔM-Env. Recovery of the RABV-ΔM-Env vaccine was conducted using the previously described recovery system, 29,42 propagated on BSR-RABVM cells that stably provide RABVM in trans, 29,35 then purified and concentrated over 20% sucrose in an SW28 rotor (Beckman, Inc.) at 24,000 rpm for 1 h at 4°C. Purified virus pellets were suspended in sterile phosphate-buffered saline (PBS). Vaccines were titered three times, each in duplicate, on BSR-RABVM cells.
Vaccine characterization
Western blotting
Expression of virally encoded proteins on the surface of virus particles or in infected BSR cells was analyzed by Western blot as described 43 using an SIVmac251 gp120 monoclonal antibody (1:2,000; KK65; NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH) 44,45 to detect SIV Env or rabbit anti-RABV ribonucleoprotein (1:2,000, RNP; a kind gift from Dr. Matthias Schnell, Thomas Jefferson University).
One-step growth curves
Growth kinetics of RABV-ΔM-Env and RABV-ΔM were analyzed by preparing a one-step growth curve using BSR-RABVM cells as described previously. 29
Flow cytometry
Expression of SIV Env on the surface of infected BSR cells was analyzed by flow cytometry analysis. BSR cells were plated at 5×105 cells per well of a six-well plate and infected with a multiplicity of infection (MOI) of 3 with RABV-ΔM or RABV-ΔM-Env for 42 h. Cells were washed with PBS, then removed with a cell stripper. Cells were blocked with 1% bovine serum albumin (BSA)/10 mM glycine in PBS for 30 min at room temperature. Cells were stained with 1:200 KK65 in 1% BSA in PBS for 1 h at room temperature. After washing in 1% BSA, cells were stained with 1:450 goat antimouse IgG-647 for 1 h at room temperature in the dark. After being washed in BSA, cells were fixed in 2% paraformaldehyde in PBS for 30 min at 4°C. Cells were suspended in 2% heat-inactivated serum in PBS for analysis on a BD LSR II flow cytometer.
Immunization schedule
Priming immunization
Groups of five female 6- to 8-week-old C57BL/6 mice were immunized intramuscularly (i.m.), intradermally (i.d.), or intranasally (i.n.) with 4×105 or 106 foci-forming units (ffu) of RABV-ΔM or RABV-ΔM-Env or with an equal volume of PBS. Blood was collected via retroorbital bleeds at the specified weeks postimmunization to collect serum, which was stored at 4°C until analysis.
Boosting immunizations
Twenty-eight days after prime immunization, mice were boosted with 10 μg rgp130 SIVmac251 (ImmunoDX) or with 4×105 or 106 RABV-ΔM, RABV-ΔM-Env, or an equal volume of PBS. Blood was collected via retroorbital bleeds at weeks 2, 3, and 4 postboost to collect serum.
Enzyme-linked immunosorbent assays (ELISAs)
Serum anti-RABV G total IgG antibodies were determined by ELISAs as described previously. 30 Serum anti-SIV Env total IgG, IgG1, and IgG2c antibodies were determined as follows: 96-well plates (Nunc maxisorp) were sealed and incubated overnight at 4°C with 0.5 μg/ml rgp130 (ImmunoDX) in PBS with a volume of 100 μl per well. Antigen was removed and plates were blocked with 100 μl of 1% BSA in PBS, and incubated 1 h at room temperature (RT). Plates were then washed four times with 200 μl/well PBS/0.075% Tween-20 using a BioTek ELx405 plate washer. A 1:50 dilution of serum samples in PBS/0.075% Tween-20 in a volume of 100 μl was added in duplicate, serially diluted 1:3 twice, and then incubated for 1 h at RT. Plates were then washed four times with 200 μl/well PBS/0.075% Tween-20 using the plate washer. A 1:5,000 dilution of HRP-conjugated goat antimouse IgG in PBS/0.075% Tween-20 was added to each well in a volume of 100 μl, and incubated at 37°C for 30 min. Plates were then washed four times with 200 μl/well PBS/0.075% Tween-20 using the plate washer. SIGMAFast OPD (Sigma-Aldrich) was then diluted according to the manufacturer's instructions and 200 μl was added to each well and incubated in the dark for 30 min at RT. Then 50 μl 3 M H2SO4 was added to each well to stop the reaction. The optical density (OD) at 490 nm was measured. Statistical differences in OD490 between groups were determined with the Student's t test.
Virus neutralizing assay
Preimmune and postimmune serum samples from animals were assayed against the neutralization-sensitive SIVmac251.6 and the neutralization-resistant SIVmac239CS.23 Env-pseudotyped virus via the TZM-bl assay as described. 46 Animal sera can interfere with neutralization assays (both positively and negatively), especially using Env-pseudotyped luciferase assays. 47 This interference can be animal specific. To correct for intraanimal differences resulting from assay interference, we analyzed preimmune sera in parallel with postimmune sera to evaluate neutralization properties within an immunization group. Criterion for positive neutralization activity was a titer greater than 3-fold that was detected in preimmune serum from the same animal against the same virus. 4 –50
Antibody avidity assays
SIV Env-specific antibody avidity was determined by sodium thiocyanate ELISA as described elsewhere, 29 except that 96-well ELISA plates were coated with 0.5 μg/ml of rgp130 in PBS overnight at 4°C.
Antibody-dependent cellular cytotoxicity (ADCC) assay
Our ADCC assay is a modified version of the rapid and fluorometric antibody-dependent cellular cytotoxicity assay (RFADCC) described by Gomez Roman et al. to measure ADCC activity from immunized mice. 51 Briefly, target cells, murine 3T3.T4.CCR5 fibroblasts (NIH AIDS Reagent Program) 52 were labeled with 4 μM PKH-26 and 10 μM CFSE and pulsed with recombinant Env at a concentration of 3 μg/1.6×106 cells in a 100 μl volume. Mouse splenocytes enriched for natural killer (NK) cells with interleukin (IL)-15 and IL-2 were used as effector cells. For the assay, 5×103 PKH/CFSE-labeled and rgp130-pulsed target cells were incubated in a 96-well U-bottom plate with sera from immunized or control mice at 37°C for 30 min before the addition of 5×105 effector cells to each well. Plates were incubated for 4 h at 37°C before the cells were fixed in 2% PFA. Cells were resuspended in 2% heat-inactivated serum in PBS and then analyzed on the BD LSR II flow cytometer. CFSE was read with the FITC channel and PKH-26 was read with the PE channel. ADCC activity was measured as the percent of target cells killed (CFSE negative) out of the total target cell population (PKH-26 positive). ADCC levels three times higher than background levels were considered significant. 51
Results
Construction, recovery, and characterization of RABV-ΔM expressing SIV Env (RABV-ΔM-Env)
To investigate whether RABV-ΔM could serve as an antibody-based HIV vaccine, we constructed and recovered RABV-ΔM expressing the ectodomain and transmembrane domain of the envelope protein from the mac239 strain of SIV (SIVmac239Env) (Fig. 1A) fused in-frame with the RABV CD. The RABV G CD incorporates foreign glycoproteins into the surface of RABV particles. 38,41,53
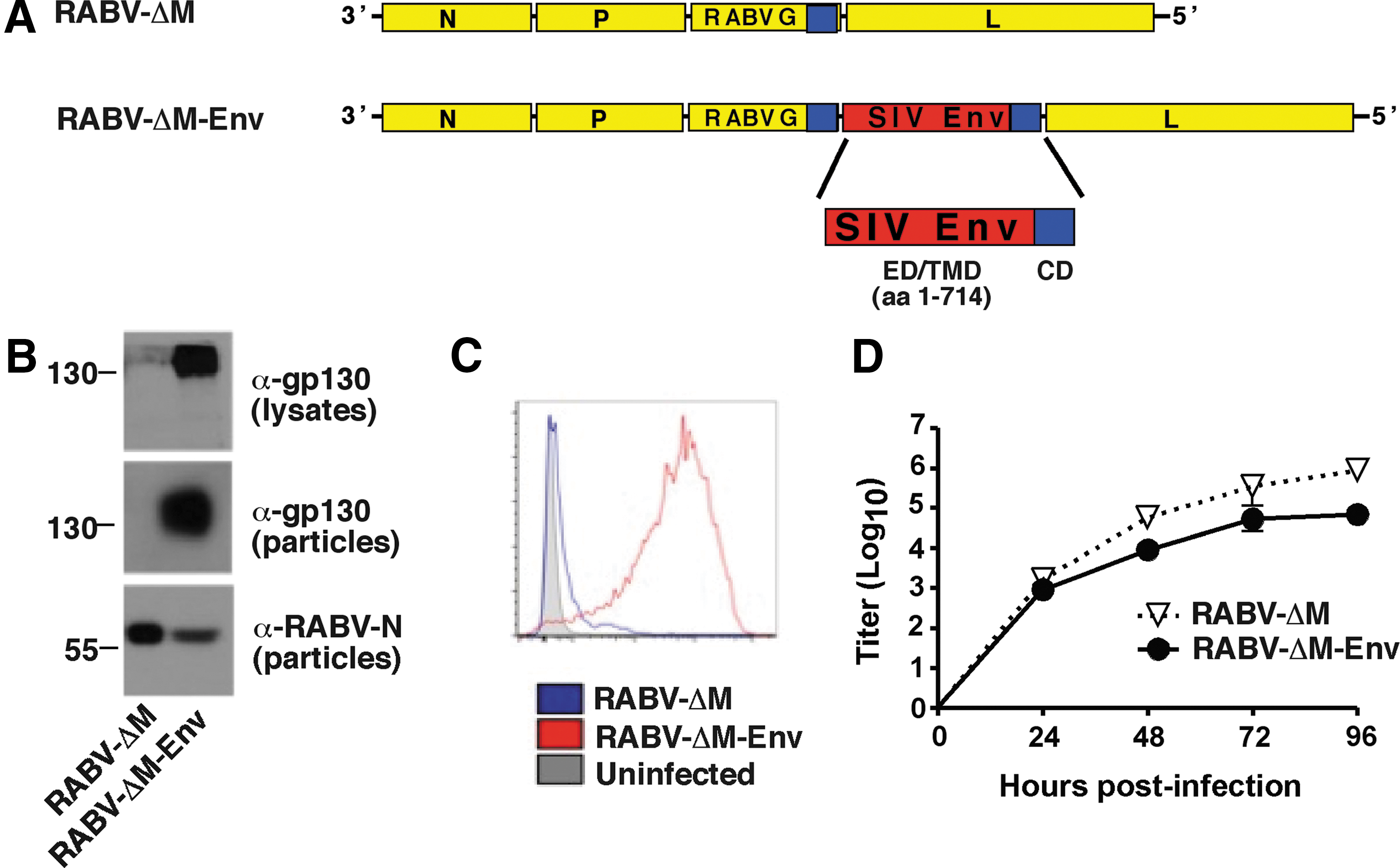
Construction and characterization of rabies virus (RABV)-ΔM-Env.
Western blot analysis of lysed BSR cells infected with RABV-ΔM-Env confirmed the expression of SIV Env from RABV-ΔM-Env but not from RABV-ΔM (Fig. 1B, top blot). Incorporation of SIV Env into viral particles was verified via Western blot analysis of sucrose-purified RABV-ΔM-Env particles (Fig. 1B, middle and bottom blots). Expression of SIV Env on the surface of infected BSR cells was also confirmed via flow cytometry analysis (Fig. 1C). A multicycle growth curve shows that RABV-ΔM-Env grows to 5- to 10-fold less titers than does RABV-ΔM on BSR-RABVM cells (Fig. 1D), which was sufficient to complete the vaccine studies described in this article. All together, a replication-deficient RABV-based vaccine expressing SIV Env on the surface of infected cells and that incorporates SIV Env into viral particles can be recovered and grown to sufficient titers.
RABV-ΔM-Env induces anti-SIV Env and anti-RABV G antibody responses
To determine whether a single inoculation with RABV-ΔM-Env was sufficient to induce seroconversion against both SIV Env and RABV G, mice were immunized i.m. with 106 ffu/mouse with RABV-ΔM-Env, RABV-ΔM, or PBS (mock). Serum was collected at various times postimmunization to evaluate the kinetics of the induced antibody responses. As shown in Fig. 2A, RABV-ΔM-Env induced significant anti-SIV Env antibodies compared to mice immunized with RABV-ΔM or PBS alone. Anti-RABV G antibodies were maintained in mice immunized with RABV-ΔM-Env compared to RABV-ΔM (Fig. 2B), indicating that the expression of two envelope proteins from a chimeric RABV-ΔM/SIV virus does not adversely affect the induction of effective anti-RABV immunity.
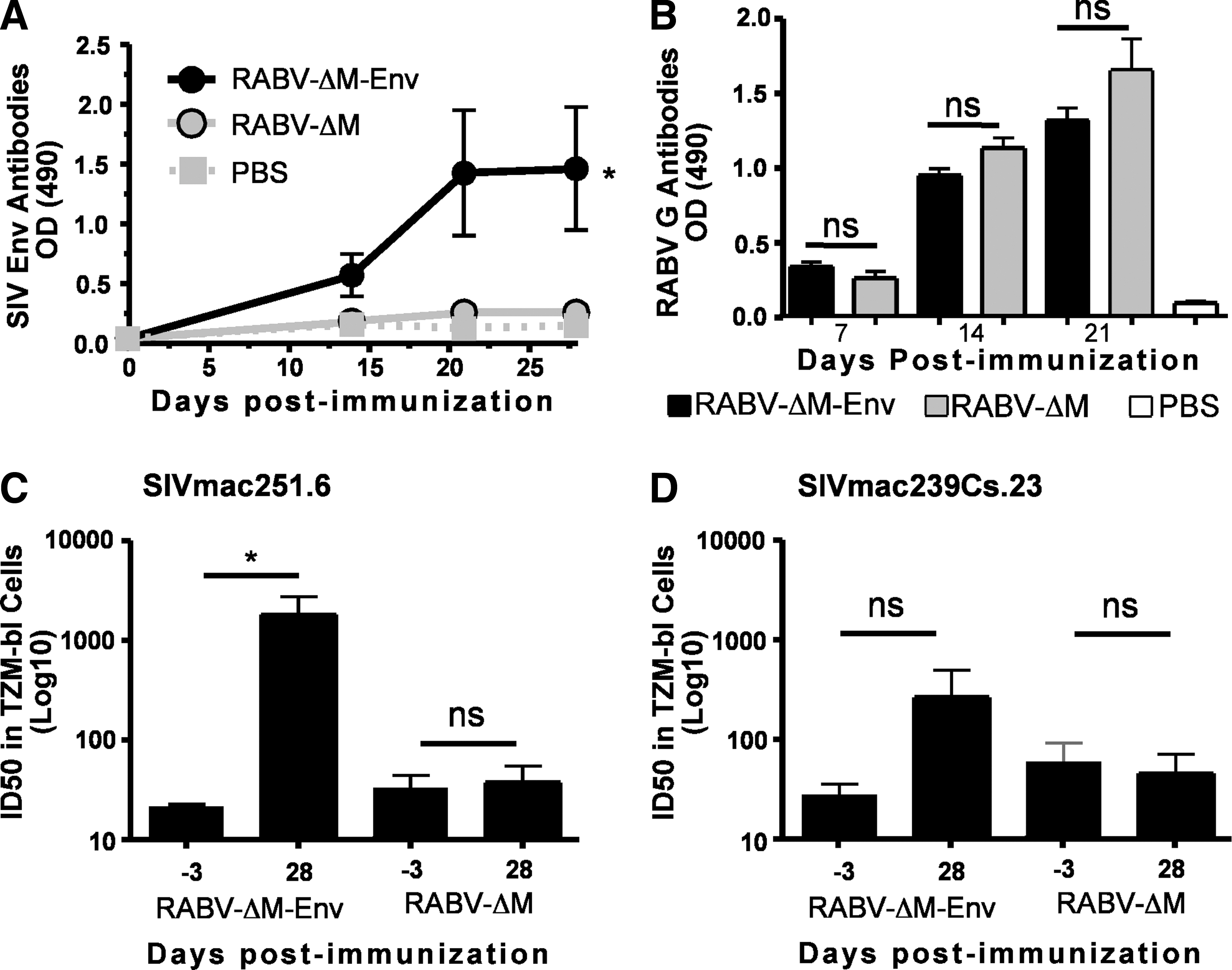
A single intramuscular inoculation of mice with RABV-ΔM-Env induces significant levels of anti-SIV and anti-RABV antibodies. Groups of 15 mice were immunized in the gastrocnemius muscle with 106 ffu of RABV-ΔM-Env, RABV-ΔM, or an equal volume of phosphate-buffered saline (PBS).
Consistent with antibodies detected by ELISA, positive SIV-specific neutralization activity was detected in mice immunized with a single dose of RABV-ΔM-Env 28 days postimmunization against a neutralization-sensitive SIV strain (SIVmac251.6; Fig. 2C). Low levels of VNAs against a neutralization-resistant SIV strain (SIVmac239Cs.23; Fig. 2D) were detected in mice primed with RABV-ΔM-Env by day 28 postimmunization, although the NAb titer did not exceed the criterion for positive neutralization activity (i.e., >3×the preimmune neutralization titer for the same animals against the same virus). Together, the data indicate that a single i.m. dose of RABV-ΔM-Env induces NAb responses against a neutralization-sensitive SIV strain. In addition, vaccine-induced anti-RABV G antibodies are not adversely affected by the expression and incorporation of SIV Env into viral particles, suggesting a chimeric RABV-ΔM/SIV virus might serve as a vaccine against two relevant pathogens posing public health threats.
A boost inoculation with recombinant SIV gp130 or RABV-ΔM-Env enhances the quality, but not the quantity, of the anti-Env antibody response
Two prime-boost strategies were tested to determine the effect of boosting the immune response compared to a prime-only strategy: (1) a live virus prime (RABV-ΔM-Env) followed by a live, homologous virus boost (RABV-ΔM-Env) and (2) a live virus prime (RABV-ΔM-Env) followed by a recombinant SIVmac251 Env protein boost (rgp130). Consistent with the results shown in Fig. 2, positive neutralization activity was detected against a neutralization-sensitive SIV strain in groups of mice primed with RABV-ΔM-Env on days 21 and 28 postboost, although boosting with rgp130 or RABV-ΔM-Env did not appear to increase the magnitude of the response compared with boosting with PBS alone (Fig 3A). Mice primed with RABV-ΔM, which does not express SIV Env, did not induce SIV-specific NAbs against neutralization-sensitive SIV, except when boosted with RABV-ΔM-Env, also confirming that a single dose of RABV-ΔM-Env induces significant anti-SIV NAb titers (Fig. 3A).

A single inoculation with RABV-ΔM-Env induces NAbs against neutralization-sensitive and -resistant SIV strains. The mice in Fig. 2 were divided into three groups of five mice/group and then boosted 35 days postimmunization with PBS (i.m), 10 μg recombinant gp130 (i.d.), or 106 ffu RABV-ΔM-Env (i.m.). On days 21 and 28 postboost (i.e., 56 and 63 days postprime), blood was collected and sera analyzed for their ability to neutralize the neutralization-sensitive SIV strain SIVmac251.6
Positive neutralization activity was detected against a neutralization-resistant SIV strain 63 days postimmunization with RABV-ΔM-Env (PBS boosted) (Fig. 3B, black bars, day 28 postboost), consistent with the notion that NAbs against heterologous or neutralization-resistant HIV take time to develop via extensive somatic hypermutation. 54 –61 Boosting mice with RABV-ΔM-Env did not appear to increase the magnitude of the response compared with boosting with PBS alone (Fig 3A). It is interesting to note that mice primed with RABV-ΔM-Env and then boosted with rgp130 did not show neutralization activity against the neutralization-resistant SIV strain. Based on the importance for priming with RABV-ΔM-Env to induce neutralization activity against neutralization-sensitive and -resistant SIV strains, the lack of neutralization activity in mice primed with RABV-ΔM-Env and then boosted with rp130 most likely resulted from these mice not reacting to the initial prime, rather than the rgp130 inhibiting neutralization activity. All together, the data indicate that a single inoculation with RABV-ΔM-Env induces neutralization activity against a neutralization-sensitive SIV strain rapidly while neutralization activity against a neutralization-resistant SIV strain takes time to develop. In addition, the data indicate that a boost inoculation does not increase the magnitude of these responses.
While we show that a boost inoculation does not enhance the magnitude of the induced antibody response, a boost inoculation has the potential to enhance the quality of an antibody response. Mice primed with RABV-ΔM-Env and then boosted with gp130 or RABV-ΔM-Env showed enhanced antibody avidity compared to preboost avidity results (Fig. 4A and B). In addition, mice primed with RABV-ΔM-Env and then boosted with rgp130 showed an increase in the ratio between IgG2c and IgG1 antibodies (Fig 4C and D), suggesting a Th1-type bias with this immunization protocol. The avidity and antibody isotype data indicate that a prime-boost strategy involving either a boost with RABV-ΔM-Env or gp130 has the potential to enhance the quality of the antibody response compared to a prime inoculation alone.

A boost with RABV-ΔM-Env or rgp130 enhances the quality of the RABV-ΔM-Env-induced antibody response. Sera collected from mice for Figs. 2 and 3 were analyzed for Env-specific antibody avidity 28 days postprime inoculation
A low-dose i.d. vaccination induces significant antibodies compared to an i.m. inoculation
Next, we sought to determine whether i.d. or i.n. inoculation enhances vaccine-induced antibody responses compared with an i.m. inoculation. Mice were immunized i.m., i.d., or i.n. with a low-dose of RABV-ΔM-Env (4×105 ffu/mouse). As shown in Fig. 5A, RABV-ΔM-Env induced significant anti-SIV antibodies when administered via the i.d. route compared to either i.m. or i.n. inoculation, and anti-RABV G antibodies were lower but were still maintained in mice inoculated i.d. (Fig. 5B). These data indicate that the i.d. route of inoculation might make for an effective low-dose RABV-ΔM-Env vaccination strategy against both rabies and HIV, which could be especially beneficial in developing countries.
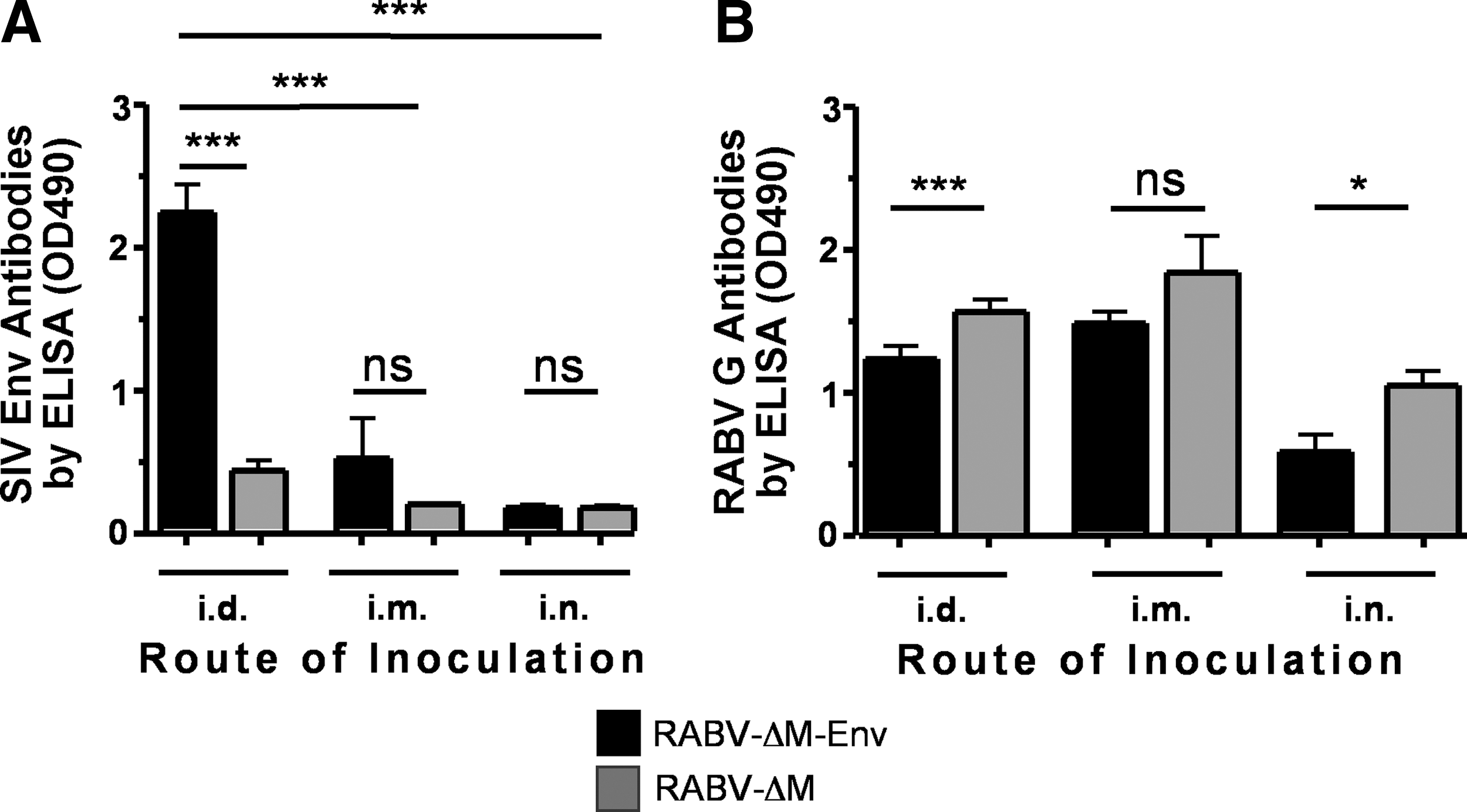
A low-dose intradermal inoculation induces significantly higher anti-SIV Env antibody responses than an intramuscular or intranasal inoculation. Groups of five mice were inoculated i.d., i.m., or i.n. with 4×105 ffu of RABV-ΔM-Env or RABV-ΔM and blood was collected 28 days postimmunization. Sera (1:50) were analyzed for anti-SIV Env antibodies
An intradermal boost with rgp130 or RABV-ΔM-Env enhances the quality, but not the quantity, of vaccine-induced antibody responses
As shown above, an intramuscular prime-boost strategy has the potential to enhance the quality but not the quantity of the induced antibody response compared to a prime-only approach. To determine whether a prime-boost strategy involving i.d. inoculation modulates the quantity and/or quality of the antibody response, mice were primed with a low dose of RABV-ΔM or RABV-ΔM-Env and then boosted with PBS, rgp130, or RABV-ΔM-Env. As shown in Fig. 6A, positive neutralization activity was detected against a neutralization-sensitive SIV strain in mice primed with RABV-ΔM-Env, either after a prime alone (day 28) or after a boost inoculation with PBS, rgp130, or RABV-ΔM-Env.
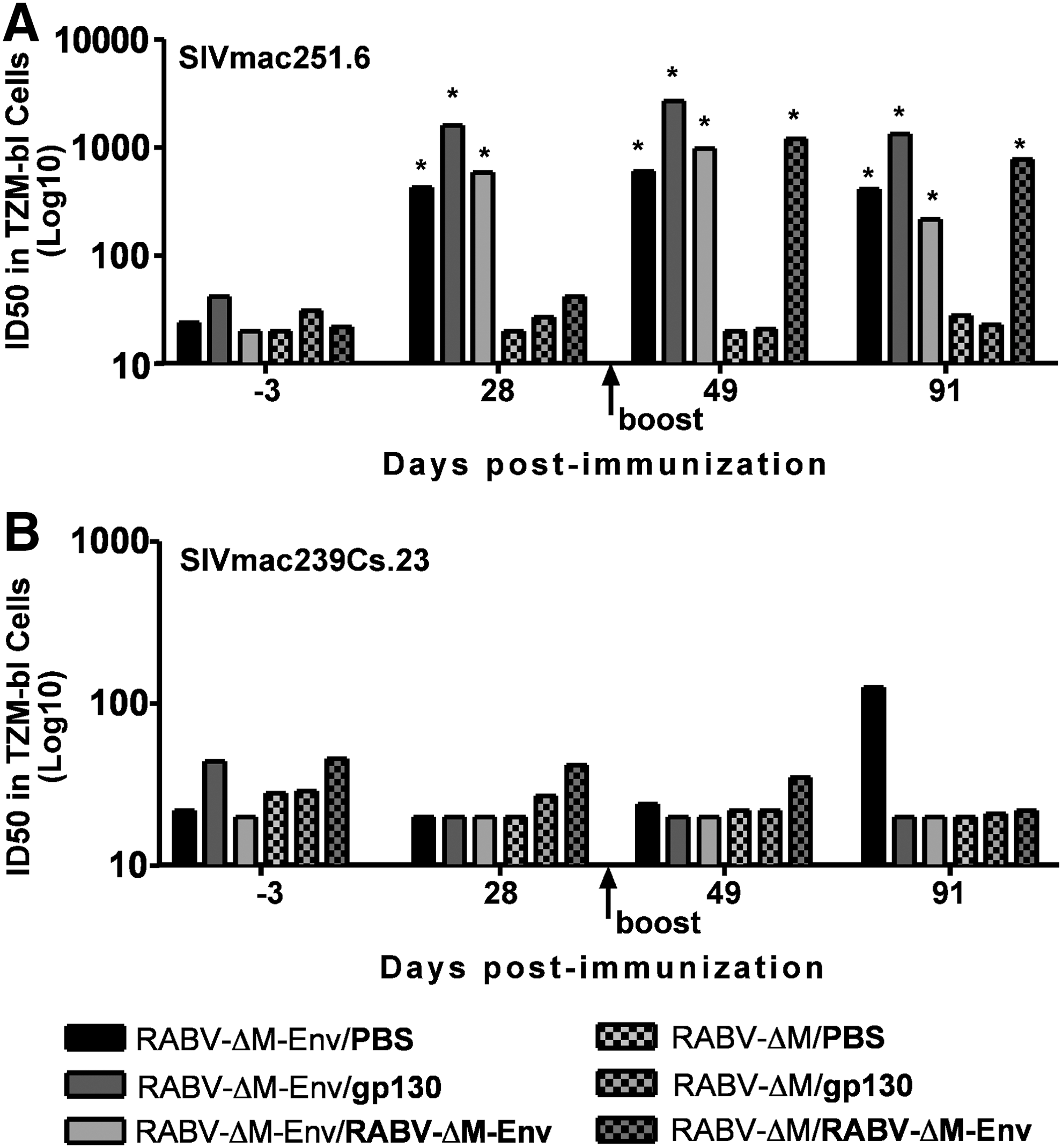
A low-dose i.d. prime with RABV-ΔM-Env induces NAb against the neutralization-sensitive SIV strain SIVmac251.6. Groups of mice were inoculated with 4×105 ffu RABV-ΔM-Env intradermally and then boosted with PBS (mock), 10 μg rgp130, or 4×10
5
ffu RABV-ΔM-Env. Blood was collected 28 days postprime inoculation or 14 and 56 days postboost and
Consistent with the neutralization data presented above in Figs. 2 and 3, boosting with rgp130 or RABV-ΔM-Env did not appear to increase the magnitude of the response compared with boosting with PBS alone (Fig. 6A). Mice primed with RABV-ΔM, which does not express SIV Env, did not induce SIV-specific NAbs against neutralization-sensitive SIV, except when boosted with RABV-ΔM-Env, also confirming the results above that a single dose of RABV-ΔM-Env induces significant anti-SIV NAb titers (Fig. 3A). At this low dose of vaccine, neutralization activity against a neutralization-resistant SIV strain was not detected above the level of a positive NAb result (Fig. 6B). While the boost inoculation did not increase the magnitude of the induced antibody response, a boost with either rgp130 or RABV-ΔM-Env skewed the immune response toward a Th1-type response, characterized by an increase in the IgG2c/IgG1 ratio detected 56 days postboost [Fig. 7A (postprime) and 7B (postboost)]. Together, the data indicate that a low dose of RABV-ΔM-Env administered i.d. is capable of inducing anti-SIV and anti-RABV G antibodies and enhances the Th1 bias of the antibody response.

A boost inoculation with rgp130 or RABV-ΔM-Env enhances the quality of the RABV-ΔM-Env-primed response. Groups of mice were inoculated with 4×105 ffu RABV-ΔM-Env, 4×105 ffu RABV-ΔM, or an equal volume of PBS intradermally and then boosted with PBS (mock), 10 μg rgp130, or 4×105 ffu RABV-ΔM-ENV. Blood was collected 28 days postprime inoculation
RABV-ΔM-Env induces antibodies with ADCC activity
The data above indicate that a single inoculation with RABV-ΔM-Env induces potent anti-SIV antibody responses and that a boost inoculation with either RABV-ΔM-Env or rgp130 influences the quality of the immune response, including the skewing toward a Th1-type response. Th1-type antibodies, characterized by IgG2c antibodies in C57BL/6 mice, are potent inducers of ADCC activity due to their affinity for activating FcRs on immune cells. To determine whether RABV-ΔM-Env alone or in combination with a boost inoculation is capable of inducing antibodies with ADCC activity, mice were immunized i.d. with a low-dose RABV-ΔM or RABV-ΔM-Env and then boosted with rgp130 or RABV-ΔM-Env. PBS-boosted mice served as controls. As shown in Fig. 8A, a slightly higher, but not significant percent killing was detected in mice primed with RABV-ΔM-Env compared to RABV-ΔM-immunized mice. However, mice primed with RABV-ΔM-Env and then boosted with rgp130 showed an increase in ADCC activity by day 14 postboost (Fig. 8B, top panel), which significantly increased to over 20% by day 52 postboost (Fig. 8B, bottom panel). The data presented in this section show that a prime-boost approach consisting of low-dose RABV-ΔM-Env followed by a rgp130 boost induces significant levels of antibodies with ADCC activity.

RABV-ΔM-Env induces antibodies with nonneutralizing antibody effector functions. Groups of mice were inoculated with 4×105 ffu RABV-ΔM-Env, 4×105 ffu RABV-ΔM, or an equal volume of PBS intradermally and then boosted with PBS (mock), 10 μg rgp130, or 4×105 ffu RABV-ΔM-Env. Blood was collected 28 days postprime inoculation
Discussion
In this article, we show that a matrix gene-deleted RABV-based vaccine expressing SIV Env is able to induce antibodies with neutralizing and nonneutralizing effector functions in mice. Together, these data suggest that antibodies induced by RABV-ΔM-Env might prevent infection and clear virally infected cells ex vivo, two key attributes for an antibody-based HIV-1 vaccine.
The rabies envelope glycoprotein (G) is acquired from the host cell plasma membrane during the budding process. The rabies CD of G is generally required for incorporation of G or foreign glycoproteins into the virus particle. 62 In the experiments described here, we fused the ectodomain and transmembrane domain of SIV gp130 to the CD of RABV G. Similar strategies have been used previously to successfully incorporate foreign proteins into RABV particles. 38,39,41,63 SIV, instead of HIV, was selected to facilitate future studies in nonhuman primate models of SIV vaccine efficacy.
The RABV-SIV fusion proteins are functional and alter the tropism of RABV particles, as demonstrated by a recombinant RABV-based vaccine expressing HIV Env fused to the RABV G CD. 63 The expression of functional, membrane-anchored SIV Env in RABV particles or on the surface of infected cells has implications for vaccine design. 64 HIV Env is naturally expressed as a trimer on the surface of viral particles and infected cells. Each trimer consists of a membrane-distal gp120 noncovalently linked to a transmembrane protein, gp41. 65 Once gp120 binds to the cellular receptor, CD4, conformational changes result in membrane fusion. 65 NAbs and nNAbs bind to conserved conformational epitopes exposed before and during receptor binding and membrane fusion. 66 Importantly, antibodies targeting epitopes on HIV Env during the entry process, as well as targeting epitopes on HIV Env during the release from infected cells, might help to prevent infection, kill infected cells, and prevent release of viral particles, 66 which are most likely critical attributes of an effective HIV-1 vaccine.
Our data suggest that RABV-ΔM-Env might induce such antibodies, as demonstrated by the neutralization and ADCC data. However, despite the development of NAb and nNAbs by RABV-ΔM-Env, the potential exists that the antigenic structure of Env expressed by RABV-ΔM-Env could be improved to enhance antibody effector functions. The introduction of intermolecular disulfide bonds between gp120 and the gp41 ectodomain (i.e., gp140 SOS molecules) produces antigen that maintains critical neutralizing epitopes in gp120 and gp41, binds to CD4, and exposes CD4i epitopes. 67 –71 We and others have shown that gp140 SOS molecules can be incorporated into viral particles, 53,72 presenting membrane-anchored antigen that reacts with MAbs 2612, b12, and 2F5. 53 A similar strategy was recently used to construct soluble, cleaved Env trimers that resemble native Env spikes both structurally and antigenically. 73
In addition to modifying the structure of the virally encoded Env protein, evidence from and expanding on the RV144 clinical trial may shed light onto the protein boost. In the RV144 vaccine trial, the protein boost consisted of two protein immunogens (A244-rp120 and MN-rgp120). Modification of A244-rp120 by an N-terminal deletion of 11 amino acids improved antigenicity and immunogenicity to conformational V1/V2 epitopes. 74 Together, modifications to the virally encoded Env proteins and the Env protein boost might help to increase the magnitude and quality of vaccine-induced antibody responses in the context of RABV-ΔM-Env.
Polyfunctional antibody responses, characterized by neutralizing and nonneutralizing effector functions, will most likely play key roles in HIV vaccine-induced efficacy. In the studies described herein, we have shown that a single inoculation with RABV-ΔM-Env induces NAbs against neutralization-sensitive and low-level neutralization against neutralization-resistant SIV strains (Figs. 2, 3, and 6). RABV-ΔM-Env induced NAbs directed against the neutralization-sensitive SIV strain rapidly, within 28 days postimmunization. On the other hand, RABV-ΔM-Env-induced NAbs directed against the neutralization-resistant SIV strain were slow to develop and were not detected until 63 days postimmunization (Fig. 3B). This is consistent with the findings that bNAbs are slow to develop, requiring 2 to 3 years during natural infection and occurring only in less than 20% of patients. 12 –14,75,76
During natural infection, the virus persists and is continuously mutating and exposing B cells to changing epitopes, increasing the affinity and breadth of the B cell repertoire. High levels of somatic hypermutation are sometimes characteristic of bNAbs, suggesting long-term B cell maturation processes. However, RABV-ΔM-Env is replication deficient and, therefore, is not expected to persist or to expose B cells to changing antigen over long periods. Despite the apparent lack of persistence of replication-deficient RABV-based vaccines, 29,30 RABV-ΔM vectors demonstrate increased antibody avidity against rabies antigens 180 days postimmunization of rhesus macaques, 29 suggesting the long-term presence of antigen following immunization. The mechanism by which RABV-ΔM promotes longer-term B cell maturation processes is not known, but could have an impact on vaccine development where extensive somatic hypermutation might be desirable. Furthermore, a single strain of Env was used in both priming and boosting for the experiments described in this article (SIVmac239Env), which might limit B cell maturation in this vaccine strategy.
One potential mechanism to exploit and enhance somatic hypermutation resulting from immunization with a replication-deficient RABV-based vaccine would be boost with Envs from other strains to promote the expansion of B cell clones and affinity maturation, thereby potentially increasing the breadth of the antibody response. 77 –81 Finally, strain diversity is lacking among SIV strains, limiting the ability to detect the breadth of antibody responses in this model. Nonetheless, the preliminary studies presented here indicate that RABV-ΔM-Env holds promise to induce NAb responses, although improvements would be needed, as discussed below.
In addition to detecting NAbs against neutralization-sensitive and neutralization-resistant SIV strains, we detected anti-SIV nNAbs, as measured by ADCC activity. Our assay used primary NK cells enriched from naive mice. However, the potential exists that mechanisms in addition to NK-mediated ADCC activity are induced by RABV-ΔM-Env and would be important for nNAb effector functions against HIV, including antibody-dependent cellular phagocytosis (ADCP) and antibody-dependent complement-dependent cytotoxicity (ADCDC). Collectively, the inhibition of viral production from these effector functions is termed antibody-dependent cell-mediated viral inhibition, or ADVI. 26 During these processes, antibodies bind to HIV-1-specific antigens displayed on the viral or infected cell surface. The Fc antibody fragment then interacts with FcRs on immune cells, resulting in phagocytosis of viral particles or the release of mediators that kill infected cells. These mediators include perforin, granzymes, cytokines, chemokines, proteases, nitric oxide, reactive oxygen radicals, and/or Fas/FasL interactions. 1,82 –86 Due to the importance of antibody Fc binding to FcRs on immune cells, qualitative antibody attributes are important for the induction of vaccine-induced effector functions, including antibody affinity to antigen and FcRs, 87 epitope targeting, 66,88 antibody titer, subclass, 28 and glycosylation state. 26,27,89,90
An important parameter for the effectiveness of different IgG subclasses to induce ADCVI relies on binding efficiencies to either activating or inhibiting Fc receptors (FcRs) on cells of the innate immune system. 91 –93 In mice, IgG1 antibodies bind to the inhibitory FcγR, FcγRIIB, with a much higher affinity than to activating FcγR, FcγRIII. 94 Due to this higher binding affinity to FcγRIIB than to FcγRIII, IgG1 provides a dampening effect that tightly controls the resulting effector functions. 94 Conversely, IgG2a binds with much higher affinity to activating FcγRIV than inhibitory FcγRIIB. In addition, it appears that Th1 induction increases the expression of both IgG2a and its activating FcγRs (FcγRI and FcγRIV), which amplifies the IgG2a-mediated effector function. 92 This is consistent with the findings that IgG2a antibodies are potent inducers of antiviral effector functions. Therefore, a vaccine that can elicit potent IgG2a antibodies, while minimizing or eliminating IgG1 responses, may prove beneficial. Designing vaccines to induce specific qualitative and quantitative attributes will be important for the rational design of improving RABV-ΔM-Env and other vaccine platforms. 26
Other viral vector systems have been tested for their ability to induce HIV-specific B cell responses. 95 –97 Replication-deficient poxvirus- and adenovirus-based vaccines are perhaps the best studied viral vectors in the context of an HIV vaccine. Early immunogenicity studies of nonreplicating adenovirus expressing HIV-1 env in mice showed that a boost was required to induce anti-HIV-1 Env antibodies using higher titers of vaccine. 98 In addition, efficacy using Ad5-based vectors was limited in phase IIb clinical trials. 96 Early human clinical studies using a canarypox-based vector (ALVAC) demonstrated modest HIV antibodies and low cellular responses. 99,100 However, a prime-boost regimen with ALVAC and a recombinant gp120 boost showed significant protection against acquiring HIV infection in the RV144 vaccine trial. 101 While efforts are ongoing to identify immune markers responsible for this modest protection, 102 polyfunctional B cell responses might have contributed to the protection observed. 28,103
In this article we describe prerequisite mouse immunogenicity studies indicating that a newly developed RABV-ΔM-based vaccine expressing SIV Env induces antibodies with neutralizing and nonneutralizing effector functions. Future work includes identifying antigen structures that enhance NAb and nNAb effector functions, determining the longevity and breadth of the antibody response, identifying those effector functions that significantly contribute to the control of free and cell-associated HIV virus, identifying novel adjuvants that promote multifunctional B cell responses, and testing HIV vaccine efficacy in nonhuman primate models.
Footnotes
Acknowledgments
This study was supported by (1) the National Institute of Allergy and Infectious Diseases (NIAID) Division of Intramural Research Grant R21AI096907 to J.P.M. and (2) NIAID Contract #HHSN27201100016C to D.C.M. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The following reagents were obtained through the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH: (1) SIVmac251 gp120 Monoclonal Antibody (KK65) from Dr. Karen Kent and Miss Caroline Powell and (2) 3T3.T4.CCR5 from Dr. Dan R. Littman.
Author Disclosure Statement
No competing financial interests exist.