
Abstract
Many posttranscriptional processes are known to regulate gene expression and some of them can act as an antiviral barrier. The nonsense-mediated mRNA decay (NMD) was first identified as an mRNA quality control pathway that triggers rapid decay of mRNA containing premature stop codons due to mutations. NMD is now considered as a general posttranscriptional regulation pathway controlling the expression of a large set of cellular genes. In addition to premature stop codons, many other features including alternative splicing, 5′ uORF, long 3′ UTR, selenocystein codons, and frameshift are able to promote NMD. Interestingly, many viral mRNAs exhibit some of these features suggesting that virus expression and replication might be sensitive to NMD. Several studies, including recent ones, have shown that this is the case for retroviruses; however, it also appears that retroviruses have developed strategies to overcome NMD in order to protect their genome and ensure a true expression of their genes. As a consequence of NMD inhibition, these viruses also affect the expression of host genes that are prone to NMD, and therefore can potentially trigger pathological effects on infected cells. Here, we review recent studies supporting this newly uncovered function of the NMD pathway as a defense barrier that viruses must overcome in order to replicate.
Introduction
S
The NMD Pathway
mRNA translation is a major step in determining the fate of an mRNA. The translation initiation is handled by functionally distinct complexes at the 5′ extremity of mRNA. 5 Thus, the pioneer round of translation is initiated on mRNA bound to the nuclear CBP80-CBP20 complex, while during steady-state translation, the 5′ cap is associated with EIF4E instead. While it has been well described that ribosome scanning of CBP80-CBP20-bound mRNAs before steady-state translation 4,6 –9 supports NMD, recent studies revealed that NMD could occur on EIF4E-bound mRNAs as well. 10,11
NMD initiation: UPF1 recruitment
According to the current model, once the ribosome is stalled on a stop codon, the eukaryotic release factors eRF1 and eRF3 are recruited. Under normal translation termination, these factors bind to PABPC1, which is associated with the poly(A) tail, and ribosome release is therefore stimulated.
12,13
In addition, PABPC1 represses NMD by outcompeting the NMD initiator factor UPF1 for an association with eRF3.
2,4
By contrast, the exon junction complex (EJC), which is deposited onto mRNA as a consequence of splicing, stimulates NMD initiation when positioned downstream of a stalled ribosome by favoring the recruitment of the NMD factors UPF2 and UPF3.
14
–16
UPF1 also accumulates on 3′ untranslated regions (3′ UTR) in a translation-independent manner,
17
–19
which might facilitate its further association with the ribosome. Thus, translation termination events that trigger NMD depend on the organization of the RNA sequence downstream of the first stop codon encountered by the ribosome. UPF1 is then recruited to the ribosome by eRF3 along with the SMG1 kinase complex (SMG1-SMG8-SMG9)
8,20,21
to form the SURF (
Subsequently, UPF1 associates with UPF3 via UPF2. UPF3 is described as an EJC component. 2,22,23 Alternative NMD pathways have been described with UPF1 interacting with UPF2 and UPF3 in the absence of an EJC, 24,25 with UPF2 alone, or even with the EJC factor BTZ replacing UPF2 and UPF3. 26,27 The association of the SURF complex with the EJC induces UPF1 phosphorylation. 8,28 UPF1 phosphorylation status is related to its ATPase and helicase activities 29 –32 and phospho-UPF1 associates with the translation initiation factor eIF3, 33 a modular complex that binds the 40S ribosome subunit and recruits capped mRNAs by interacting with CBP20-80 or eIF4F. 34,35 Interestingly, phospho-UPF1 has been shown to interact with EIF3A, while UPF2 interacts with EIF3E/INT6. 33,36 UPF1/eIF3 interactions, which repress translation by preventing the ribosome reassociation, are thought to be an important step for mRNA decay. 33
mRNA degradation
It has recently been established that phospho-UPF1 constitutes a platform for recruiting the SMG6 endonuclease and the SMG5/SMG7 complex through N-terminal and C-terminal phosphoresidues, respectively. 37,38 SMG6 directly cleaves targeted mRNA near the PTC while SMG5/SMG7 may serve as a link between UPF1 and mRNA decay factors such as XRN1 and DCP2. 39 –42 Alternatively, DCP1a is recruited by SMG5 in complex with PNRC2. 43 UPF1 dephosphorylation is achieved by the SMG5/SMG7/PP2A phosphatase complex 37 and is dependent on UPF1 ATPase activity. 44 UPF1 ATPase and presumably helicase activities would promote mRNP remodeling and would facilitate mRNA degradation and factors recycling in an yet unknown exact timing. 31
The subcellular localization of mRNA decay is still undefined. By using a chemical NMD inhibitor or by knocking down DCP1 and XRN1, cytoplasmic aggregates called processing bodies (P-bodies) accumulate NMD-targeted mRNAs and NMD factors. 45,46 However, the impairment of the formation of P-bodies by knocking down its components does not impede NMD and NMD-deficient UPF1 mutants still colocalize with P-bodies. 47,48 Thus, it appears that decay can occur independently of P-bodies.
The Retroviral Replication Cycle
Retroviruses are enveloped viruses with two identical copies of a single-stranded (+) RNA molecule, which are 5′ capped and 3′ polyadenylated. After the virus attaches to specific cell receptors, viral glycoproteins mediate the fusion of virus envelope with the cell membrane. The single-stranded (+) RNA genome is then delivered into the cytoplasm, partially uncoated and reverse transcribed. The reverse transcription is initiated by a tRNA, specific of each virus, and synthesizes a DNA strand complementary to the viral RNA. 49 Then, the RNase activity of the reverse transcriptase degrades the RNA strand before a second DNA strand is synthesized to generate a double-stranded DNA. By this process, the RNA sequences R-U5 (at the 5′ extremity) and U3-R (at the 3′ extremity) generate two long terminal repeats (LTR) sequences (U3-R-U5) flanking the viral coding region. The viral DNA is circularized, moves in the nucleus, and randomly integrates into host genomic DNA. 50,51 The integrated provirus utilizes promoter elements in the 5′ LTR to drive transcription by host RNA polymerase II.
Four classes of mRNA are finally exported to the cytoplasm (Fig. 1): • Unspliced mRNA (genomic RNA), which are exported as genomic RNA and are either packed into virion particles or translated into structural proteins Gag and Gag-Pro-Pol and/or Gag-Pro (in the case of alpha, beta, and deltaretroviruses). • Singly spliced: the env mRNA from all retroviruses is produced after a single splicing event. Complex retroviruses also use singly spliced RNA for some auxiliary protein translation. • Doubly spliced mRNA: lentiviruses and deltaretroviruses RNA also undergoes alternative splicing leading to tat, rev, and nef mRNA and tax, rex, and p30 mRNA, respectively. • Antisense RNA: the LTR promoter initiates transcription in both directions. Antisense ORFs were found in lentiviruses
52,53
and deltaretroviruses
54
and are transcribed under the control of the 3′ LTR.
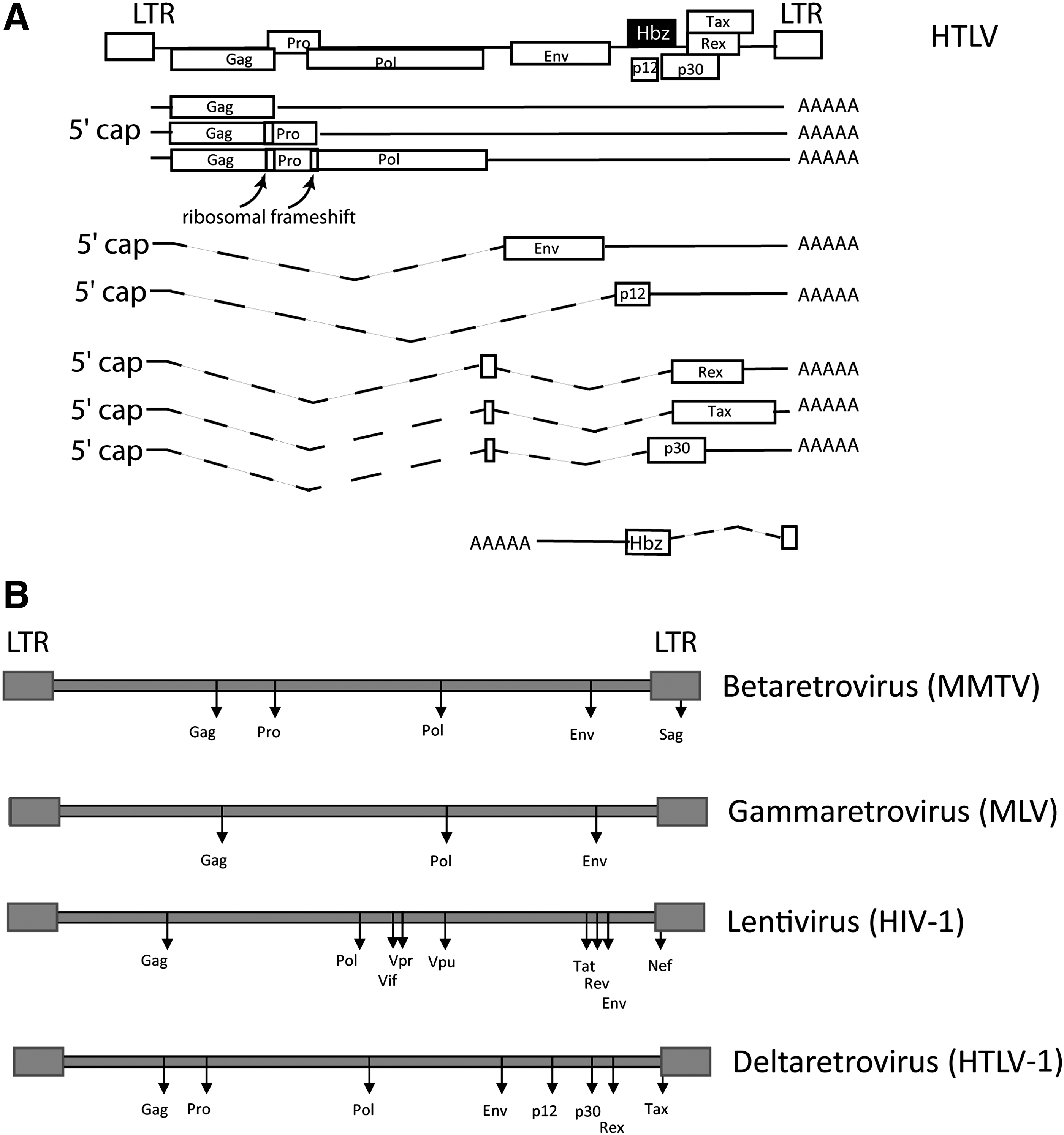
Genomic organization of retrovirus. (
Retroviral mRNA Propensity to NMD: From Analysis of Initiating Features to Viral Bypass Strategies
Originally, NMD-sensitive PTC were defined as stop codons located at least 50 nucleotides (nt) upstream of an exon–exon junction. In these conditions, an EJC located downstream of the stop codon stimulates the association of UPF3 and UPF2 with UPF1. Thus, point mutations that create a PTC upstream of an EJC, either directly or as a consequence of frameshifting or splicing alteration, render mRNA sensitive to NMD. During hematopoiesis, V(D)J rearrangements for the T cell receptor beta or immunoglobulin genes are a major source of PTC-containing mRNA, with two-third of the recombination events being nonproductive. These transcripts generated from out of frame alleles are efficiently degraded by NMD. 55 –57
All retroviral genomic RNAs, and therefore all retroviral mRNAs regardless of their class, are frequently altered not only due to the low proofreading activity of polymerases (especially retroviruses reverse transcriptases that introduce 10−3 to 10−4 error per incorporated nucleotide) but also as a consequence of RNA editing. 58,59 Among RNA editing factors, the members of the APOBEC family are cytidine deaminases that generate G-to-A hypermutations and can promote the appearance of stop codons. APOBEC3G (A3G) markedly affects the human immunodeficiency virus (HIV) genome, with up to 20% of all minus-strand dC residues being deaminated to dU. However, HIV-1 Vif counteracts this A3G effect by inducing its proteolytic degradation. 60 Although the murine leukemia virus (MLV) is also highly sensitive to A3G, 61 the impact of A3G on human T cell leukemia virus (HTLV) RNA is still unclear. 62 –64
In terms of surveillance of the quality of gene expression, NMD inhibition may seem counterproductive for the virus since it would allow the expression of truncated proteins that could not support viral replication. However, it may provide an opportunity for immune system evasion by allowing the production of nonimmunogenic albeit active truncated proteins. However, NMD as a general posttranscriptional regulator of gene expression can be activated in a much more complex context: programmed frameshift, long 3′ UTR, 5′ uORF, selenocysteine codons, and alternative splicing events, since all of them would create a 3′ UTR with an abnormal organization. Interestingly, the various classes of retroviral mRNA exhibit almost all these features (Fig. 1). Hereafter, we will describe their localization on retroviral mRNA and analyze the propensity of these mRNA to be efficiently degraded.
Unspliced RNA
Termination suppression and programmed frameshifting
As mentioned previously, for all retroviruses, a fraction of transcribed RNA needs to be exported into the cytoplasm without being spliced. Depending on the virus, the size of these full-length RNA varies from 6 kb to 9 kb. They are either packaged into new virions as genetic material or translated into structural proteins named Gag, Pro (the protease), and Pol (the reverse transcriptase). The regulation of pro and pol expression is performed by termination suppression or programmed frameshifting (except for spuma viruses). In gammaretroviruses such as MLV and epsilonretroviruses, pol gene expression is regulated by translational read-through of an in-frame UAG codon between the gag and pol coding regions.
Suppression of this termination codon is mediated by insertion of a glutamine residue with a frequency of about 5%. 65,66 Other retroviruses exploit frameshifting: this process necessitates a homopolymeric sequence called a slippery sequence and a specific pseudoknot (PK) downstream of the stop codon. 67 Those elements cause a translocation of the ribosome and a −1 codon frameshift (−1FS) leading to the repositioning of the ribosome on the pol ORF and further synthesis of a fused Gag-Pol polypeptide. In the case of deltaretroviruses, a second frameshift is necessary since Pro and Pol are also on different ORFs. 68
Such events are rare in the human genome and are thought to be remnants of ancient retroviral insertion. 66 –68 A recent study identified seven cytokine receptors with a -1FS, including CCR5, the HIV coreceptor. 69 CCR5 -1FS could be reminiscent of the HIV-1 -1FS. Similar to what computational analysis suggested on yeast, 70 the CCR5 -1FS event directs ribosomes to prematurely terminated ORFs defining a role for the combination “-1FS/NMD” in gene expression regulation. In these conditions, the -1FS is a cis-acting mRNA destabilizing element.
However, in the case of viral full-length RNA, frameshifts do not redirect ribosome on any PTC but instead avoid translation termination events that would otherwise occur. That is why it is questioned whether the slowdown of the ribosome is sufficient to initiate NMD on the genomic viral RNA. Hogg et al. 19 analyzed the impact of termination readthrough and frameshifting on NMD by studying the stability of reporter constructs with the MLV readthrough and the mouse mammary tumor virus (MMTV) -1FS. 19 They did not observe a destabilization of reporter mRNA unless they introduced a stop codon immediately after their respective signals. In contrast to what has been observed with host mRNA, gammaretroviral termination readthrough and betaretroviral frameshifting would protect viral mRNA against NMD. Although the mechanism is yet to be deciphered, it cannot be excluded that nearby RNA sequences or the pseudoknot itself inhibits NMD.
Nakano et al. analyzed the stability of constructs containing HTLV-1 genomic RNA sequences (nt 1677–2594), containing Gag and Pro -1FS signals, within the second exon of the β-globin reporter plasmid. 71 In opposition to Hogg et al., they found that this hybrid mRNA was significantly upregulated upon UPF factor depletion, suggesting an NMD sensitivity of the HTLV-1 RNA due to its frameshift sequences. 71 The discrepancy between these results could be explained by (1) the specificity of each -1FS or PK sequence, (2) the fact that Hogg et al. did not analyze the stability of their reporter in the absence of the UPFs, and (3) the fact that Nakano et al. studied steady-state mRNA levels instead of the half-lives of their reporter RNA.
Long 3′ UTR can free NMD from splicing and EJC deposition
The previously described experiments exploit reporter plasmids with at least one EJC downstream of the PK/-1FS, while genomic viral RNA is not spliced and therefore not marked by any canonical EJC. With the teachings of NMD in yeast, flies, and worm, it emerges that canonical EJCs are strong NMD stimulators in the case of PTC-triggered NMD, 72 –74 while the size and spatial configuration of the 3′ UTR would be the key feature that lead to mRNA degradation following arrest of the ribosome even at the natural stop codon (NSC). 75 –77 NMD-prone mRNA with long 3′ UTR but without a PTC are described in yeast and plants. 78,79 In human cells, the artificial lengthening of 3′ UTR renders mRNA sensitive to NMD. 25,80,81 By using a minigene coding the Ig-μ lacking its last intron, Bülher et al. also demonstrated that a PTC in the penultimate exon, downstream of the last EJC assembly site, is still able to initiate NMD. 24 NMD initiation would then rely on the enrichment of UPF1 on long 3′ UTR, 82 which may favor its competition over PABPC1 [bound to the poly(A) tail] for binding terminating ribosomes. Therefore, unspliced mRNA may undergo NMD depending on its 3′ UTR conformation.
Interestingly, an astounding characteristic of retroviral full-length transcripts is the size of their 3′ UTR. In HIV, HTLV, MMTV, MLV, or Rous sarcoma virus (RSV), gag 3′ UTR are ∼6,500 nt. As a comparison, human mRNA 3′ UTR have a mean size of 750 nt and 3′ UTR as short as 500 nt are able to trigger NMD. 25,80,83 These data in humans as well as those in yeast and plants thus illustrate the propensity of intronless mRNA such as retroviral full-length RNA to be targeted by NMD.
Impact of long 3′ UTR in full length retroviral RNA
For the RSV, pertinent data were obtained from experiments designed to determine whether the genomic mRNA was translated before packaging. By introducing a PTC upstream from the Gag NSC, the packaged unspliced mRNA was identified to be NMD sensitive, indicating that translation occurred before packaging. 84 The authors found that the deletion of a 155 nt sequence (termed RSE), located 110 nt after the Gag NSC, makes the RSV Gag mRNA NMD sensitive. 85 By specifically structuring the Gag 3′ UTR, this motif might prevent the initiation of NMD by UPF1. 86 They also observed that the RSE protects heterologous RNA from NMD when located at the downstream proximity of any stop codon. While the question was not addressed in this work, the sensitivity of the RSV unspliced mRNA in the absence of the RSE is unlikely due to the ribosomal slowdown associated with the frameshifting element since the removal of the pseudoknot as well as the RSE still leaves the Gag mRNA NMD sensitive. 85 Therefore, in the absence of the protective RSE, RSV unspliced mRNA are sensitive to NMD mostly through the presence of a long 3′ UTR downstream of the Gag ORF.
As an indication of this possibility, the full-length mRNA is half as stable upon insertion of a PTC within the Pol region. 85 The RSE is conserved among all avian retroviruses 87 and could be shared by other retroviruses. Moreover, regarding HIV-1, Hogg et al. reported that a reporter plasmid harboring the HIV 3′ LTR region as a 3′ UTR is coated with UPF1 protein in a length-dependent manner, similar to what is found with cellular NMD-sensitive mRNA. 19 However, in the case of HIV, UPF1 was shown to exert a positive effect as UPF1 expression increases the level of the 2-, 4-, and 9-kb mRNA and enhances pr55Gag expression, in opposition to what would be expected due to an NMD-inducing activity. 88 However, it is noteworthy that in this study, only steady-state RNA levels were quantified, instead of RNA half-lives. In addition, a recent study from Serquina et al. could not confirm the loss of viral mRNA after UPF1 depletion. 89
It has been reported that UPF1 and UPF3 are included into the HIV-1 ribonucleoprotein complex composed of pr55Gag and STAUFEN-1 at sites of HIV-1 virion assembly. 88 Serquina et al. also demonstrated that UPF1 is positively involved in HIV mRNA reverse transcription. 89 Although the exact details of this paradox remains to be deciphered, it is possible that HIV-1 favors the incorporation of UPF1 in specific complexes at the expense of those triggering NMD, thereby hindering mRNA decay. Corroborating those data, UPF1 and UPF3 have also been shown to be necessary during the yeast Ty1 late steps of retrotransposition such as virion-like particles (VLP) assembly or RNA remodeling during reverse transcription. The Ty1 RNA must also transit through P-bodies, which are sites where NMD should not occur (only decay intermediates aggregate here when NMD is inhibited). 90 Perhaps Ty1 RNA protects itself from NMD degradation by integrating these loci where translation is repressed before initiating VLP assembly. Similarly, RNA protection against NMD due to localization in translation-incompetent loci was also hypothesized for RSV Env RNA. 86,91
Concerning HTLV-1, we recently observed that silencing of UPF1 causes an increase in the amount of several RNA expressed from transfected HTLV-1 proviruses including the unspliced ones. 92 These experiments analyzed steady-state mRNA levels instead of mRNA half-lives and may correspond to direct as well as indirect effects of NMD inhibition. A direct effect was afterward confirmed for the unspliced mRNA by Nakano et al., who found a stabilizing effect of UPF1 knockdown over time after transcription inhibition. 71 Interestingly, HTLV-1 offers an example of a retrovirus controlling NMD sensitivity in trans through the expression of viral proteins. Indeed, we have reported that the HTLV-1 Tax protein exerts a negative effect on NMD. By interacting with both UPF1 and INT6/EIF3E, Tax inhibits the association between these two proteins, which normally occurs on NMD-sensitive mRNA. It also affects the recycling of UPF1 that remains locked in cytoplasmic P-bodies under its phosphorylated forms. 92 Nakano et al. have recently shown that in addition to Tax, the p21 and p27 forms of Rex also exhibit an NMD inhibitory effect through molecular mechanisms that remain to be understood. 71
Spliced RNA
NMD associated with alternative splicing
The genomic RNA has to be spliced to produce the env mRNA as well as mRNA coding accessory proteins. This event is shared among all retroviruses. As described previously, lentiviruses and deltaretroviruses also produce doubly spliced mRNA as well as mRNA from antisense transcription; this complex combination of transcription and splicing allows retroviruses to overcome the reduced coding capacity of their genome. However, the polycistronic nature of several retroviral mRNA as well as retention of intronic sequences are likely to favor sensitivity to NMD by introducing multiple stop codons into the same mRNA (Fig. 1B). It has been suggested that one-third of alternative splicing events could generate PTC-containing mRNA. 93 This coupling of alternative splicing and NMD (AS-NMD) is more than a simple way to control unproductive splicing products; it is also involved in the regulation of gene expression in metazoans. 94 –97
Many of these genes are splicing regulators, including SR proteins and hnRNPs, which indicates an important feedback regulation of splicing by NMD. 98 –100 In yeast, it has been shown that NMD regulates polycistronic mRNA. 101 The HIV-1 provirus is transcribed as a 9-kb polycistronic pre-mRNA that contains multiple alternative 5′ and 3′ splicing sites enabling the generation of more than 40 different mRNAs including 4 Vif, 5 Nef, 8 Tat, 12 Rev, and 16 Env alternative mRNAs. These alternative mRNAs exhibit different levels of expression and while 16 mRNAs encode gp160/120, one single isoform accounts for 80% of gp160/120 production. 102 –104 However, as described for point mutations, preventing AS-NMD would rather be counterproductive for the virus. On the other hand, it is possible that complex retroviruses regulate alternative splicing via NMD feedback modulation. HTLV-1 was shown to modulate host RNA alternative splicing and NMD. 105 This correlation would be very interesting to investigate in more detail.
5′ uORF
The superposition of multiple layers of coding sequences on a single molecule of mRNA also leads to the presence of 5′ uORF. Depending on their recognition by the translational 43S preinitiation complex and their size and position, 5′ uORF can be translated and the resulting stop codon can be recognized as a PTC with regard to the main ORF covered with EJC. 18,106 –108 For example, ATF4 mRNA includes two uORF and its half-life markedly increased by UPF1 or UPF2 knockdown, demonstrating a direct regulation by the NMD, despite the absence of PTC. 109 –111 While the presence of one uORF does not automatically lead to NMD, 112 it is noteworthy that around 49% of human transcripts contain at least one 5′ uORF. 113 HIV-1 exhibits a uORF located upstream of the Vpu AUG and overlapping with it. This minimal uORF is conserved and plays an important role in env translation. 114 RSV also has three uORF preceding the AUG initiation codon of Gag, Gag-Pol, and Env. The uORF 1 and 3 play an important role in stimulating translation and packaging of the RSV genome. 115 In these various instances, it will be interesting to determine if the observed effects rely merely on translational regulation or also affect RNA stability.
Selenocystein codon can trigger NMD
Notably, HIV-1 mRNA exhibits clusters of UGA and several potential selenocystein insertion elements (SECIS) spanning the end of Env and the start of Nef, suggesting selenoproteins may be encoded by this virus. 116,117 As discussed before, selenocysteine incorporation is triggered by a UGA codon that can be interpreted as a PTC and induces NMD, especially in the case of selenium deficiency. 118 Interestingly, some cases of HIV-1 infection 119 were correlated with subnormal selenium plasma levels, suggesting that RNA harboring this codon, especially HIV RNA, might become sensitive to NMD.
Endogenous retrovirus
Mendell et al. observed an increase of the HERV-H mRNA concentration in HeLa cells depleted of UPF1. 110 Interestingly, plasmas of HIV patients reveal a significant increase in HERV-K Env-derived RNA. 120,121 Since we suggested previously that HIV-1 could favor the incorporation of UPF1 in specific complexes at the expense of those triggering NMD, those combined data might support the possibility that NMD could control the expression of endogenous retrovirus.
Long 3′ UTR in spliced RNA
Spliced RNA shares the same polyadenylation signal as unspliced RNA, which is located at the 3′ extremity of the LTR sequence 122 and therefore generates long 3′ UTR sequences. Thus, a similar propensity to NMD should be observed for these spliced RNAs due to their 3′ UTR size as for unspliced RNAs. As an example, Withers et al. showed that in RSV, 400 nt of the 3′ UTR region of Src, can substitute for the RSE at the natural gag termination codon. As a consequence, the presence of an NMD insulator element in the 3′ UTR sequence of Src suggests its own sensitivity. 86 Similar results may be observed in HTLV and HIV where 3′ UTR ranges between ∼500 nt (Tax) and 2,100 nt (Env) (the 3′ UTR of the Hbz mRNA being about 1,450 nt long 123 ) and between ∼3,900 nt (Vif) and ∼700 nt (Env) (Nef 3′ UTR being ∼100 nt), respectively.
As first evidence, by knocking down UPF1, we observed a significant increase in all HTLV-1 singly or doubly spliced mRNA. 92 However, as mentioned earlier, these experiments analyzed steady-state mRNA levels instead of mRNA half-lives and may correspond to the direct as well as indirect effects of NMD inhibition. This increase could then be due to a transcription activation at the favor of some NMD-dependent stabilization of transcription factor. This will be discussed in the last part of this article.
Considering all RNA features inducing NMD (Fig. 2), computational analysis estimated that ∼10% of human mRNAs are directly or indirectly regulated by NMD. 109,110 Similar evaluations were obtained for A. thaliana, C. elegans, S. cerevisiae, and Drosophilia. 101,124 –127 In line with these observations, retroviruses also include various NMD-inducing features (Fig. 2). In light of this review, it appears that long 3′ UTR are a dominant feature for NMD sensitivity of retroviral RNA: all the retroviruses display this common feature in their full-length unspliced mRNA but also in some spliced ones.
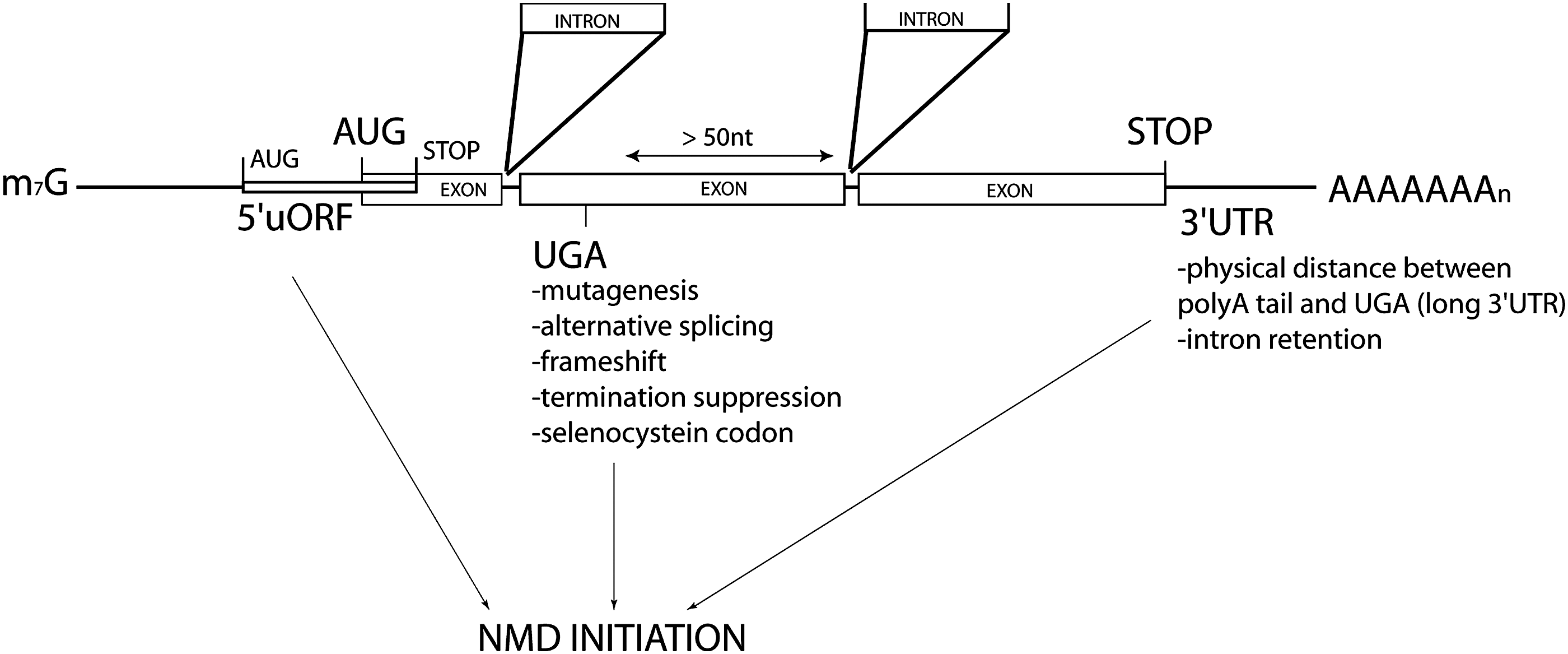
NMD-inducing features found in host and viruses RNAs. It has been extensively demonstrated that stop codons (UGA, UAG, or UAA) positioned at least 50 nt upstream of an exon–exon junction lead to nonsense-mediated mRNA decay (NMD) initiation. These codons can be generated by point mutation inducing or not frameshifting. Similarly, alternative splicing events can introduce frameshifts, which make an mRNA sensitive to NMD. Suppression of termination processes can also lead to NMD initiation. Selenocystein is encoded by a UGA codon that can be considered a premature termination codon (PTC). Finally, there is a body of evidence demonstrating that natural mRNAs, with their normal stop codon, can also be NMD sensitive: that is the case of some mRNA including a 5′ uORF (overlapping or not the main ORF) and some mRNA with a long 3′ UTR sequence, physically distancing the poly(A) tail from the stop codon. In retroviruses, the very long 3′ UTR of many RNA (especially the genomic isoform) seems to be a strong determinant of NMD sensitivity. In contrast, the programmed frameshift present between the structural genes seems to be protective, although this is still controversial.
While all genera of retroviruses were not equally studied in this matter, the demonstration of their sensitivity is assessed at least in alpharetroviruses and deltaretroviruses for their unspliced mRNA; but we believe that the conservation of retroviral sequence organization is a strong hint that suggests a general sensitivity of at least unspliced retroviral RNA to NMD due to the size of their 3′ UTR. The impact of retroviral frameshift elements is less obvious: while the host -1FS usually redirects the ribosome on a PTC-containing ORF (which explains its association with NMD), retroviral frameshifts extend the course of the ribosome beyond the Gag NSC, which per se already favors the formation of a sensitive mRNA with long 3′ UTR.
Apparently, the frameshift pseudoknot induces a specific outcome depending on the type of virus: in betaretroviruses and epsilonretroviruses, it seems to prevent RNA decay in contrast to deltaretroviruses. It is possible that through evolution, inhibitory sequences were incorporated within the frameshift pseudoknot (betaretroviruses and epsilonretroviruses), after the frameshift pseudoknot (RSV), or were nonexistent due to other defense mechanisms (HTLV-1). Thus, while specific features of the retroviral RNA can make them prone to NMD degradation, several retroviruses including RSV, HIV-1, MMTV, MLV, and HTLV-1 have acquired the means to overcome this potential negative effect via specific RNA motifs or expression of proteins negatively affecting NMD. Recently, such RNA elements were also identified in human mRNA with long 3′ UTR. 128 Whatever the inhibition strategy used, compromising the NMD is then essential for retroviruses to ensure further replication as mRNA coding structural proteins would be otherwise degraded by this pathway. Therefore, the importance of protecting the mRNA encoding structural factor overcome the potential benefit of removing PTC-containing RNA genomes with lower replication fitness from the viral population.
Deregulation of Host Gene Expression by Viral Interference with NMD
Modulation of the NMD pathway by depletion/sequestration of important factors might also impact host gene expression by affecting posttranscriptional regulation, thereby potentially favoring multiplication of the virus (Fig. 3).
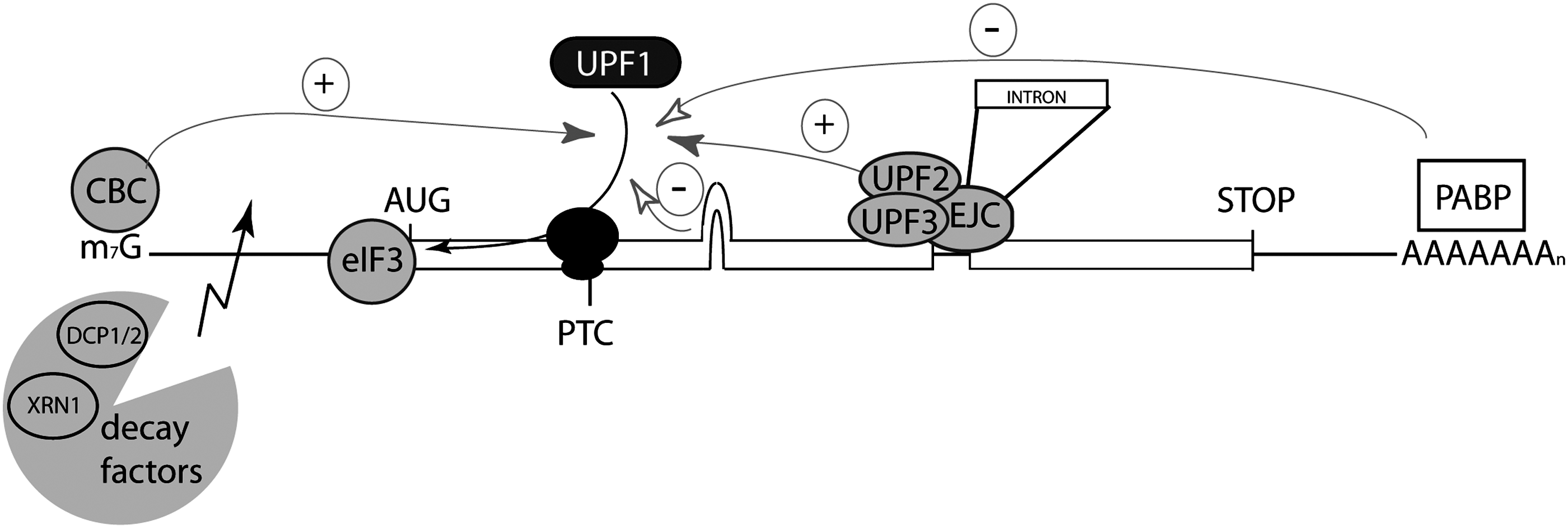
The activity of known NMD effectors can be modulated or bypassed by viruses. The recruitment of UPF1 on a stalled ribosome is enhanced in the presence of the cap binding complex (CBC). The presence of the exon junction complex (EJC) more than 50 nt downstream of the PTC with the cofactors UPF2 and UPF3 is also a strong activator of NMD initiation. The close presence of the poly(A) tail represses NMD due to the competition between UPF1 and PABPC1 for the interaction with the ribosomal complex. The interaction of UPF1 with eIF3 is considered as a primordial step in translation repression and further mRNA degradation executed by decay factors such as DCP2 and XRN1. Viruses are thought to modulate or bypass NMD by targeting these effectors through factor quenching, compartmentalization of the viral cycle, translation initiation bypass, or specific RNA secondary structures. In retroviruses, the association of viral proteins with UPF1 and eIF3 subunits was shown to inhibit NMD (HTLV-1). The specific RNA sequence in RSV blocks NMD initiation. It is suggested that the composition of HIV RNP could antagonize the induction of NMD.
By inhibiting NMD, HTLV-1 jointly protects its own genome and deregulates the posttranscriptional regulation of several cellular genes such as ATF4, GADD45α, SLIT2, BAG1, MAP3K14, IL6, DUSP10, Fyn, PTPRF, ARHGEF18, ASNS, and DEXI. Half-lives of these NMD-sensitive mRNA increased when Tax and/or Rex were expressed. 71,92 This effect might be important with respect to the disorders induced by the virus and also indirectly favors viral replication. In agreement with this, 293FT cells treated with siRNA against UPF2 and then infected by coculture with MT2 cells expressed more Gag proteins. 71 That can be correlated with the report describing the NMD target ATF4 as a potent transactivator of the HTLV-1 promoter. 129 It is noteworthy that genome-wide analyses of mRNA destabilization after UPF factor knockdown have pointed to several genes playing important roles in lymphocyte activation and survival. For instance, this is the case for the interleukin 6 and interleukin 7 receptor genes. 109,110 In addition, by generating dominant negative Upf1 mutant mice, Frischmeyer-Guerrerio et al. observed a defect in pre-TCR signaling as suggested by a decreased cell number, a reduction of CD4/CD8 double-positive thymocytes, a diminished expression of TCR-β via reduced endogenous Vβ-to-DβJβ rearrangements, and an increased expression of CD25 in fetal thymocytes. These observations suggest that NMD contributes to thymocytes maturation and expansion. 130 This hypothesis is also supported by the phenotype of Upf2 null mice. 57 Indeed hematopoietic-specific deletion of Upf2 resulted in the accumulation of nonproductive rearrangement by-products from the TCRβ locus, which are detrimental to developing T cells. As HTLV-1 can be transmitted in early childhood and infects immature T-lymphocytes, the effect of Tax on NMD might strongly interfere with the natural differentiation of thymocytes by affecting this process. A significant amount of work, including systematic approaches, is still required to validate this theory, but present data already highlight how NMD inhibition could favor the infection context by deregulating the host genes expression.
The NMD Restricts Viral Infection
By targeting viral RNA and controlling several genes involved in the immune response, NMD exhibits antiviral functions 131 similar to other retroviral restriction factors such as the APOBEC proteins family, SAMHD1, TRIM5α, and tetherin. However, the relative contribution of these various factors is still unknown. The reality of the NMD antiviral functions has been reinforced by the Helenius and the Voinnet groups, 132,133 showing the sensitivity of (+) RNA viruses (nonretroviruses) replication to UPF1 knockdown. On the other hand, the insensitivity of (–) RNA virus 132 poses the following question: Is the RNA from (+) RNA virus and retrovirus sensitive to NMD before reverse transcription in vivo? That could explain the seemingly complete resistance of the (–) RNA virus. Moreover, observations were made showing that peptides for the class I MHC would be favorably produced from NMD-prone targets. 134 NMD activation would then constitute a crucial pathway for host and virus interplay that viruses must circumvent for many reasons: protection of their RNA, limitation of peptides available for class I MHC, and modification of host transcriptional programs.
Conclusions
All gathered data indicate that retrovirus mRNAs are likely to be targeted by the mRNA stability control pathway called NMD, mainly because of the size of their 3′ UTR. In parallel, viruses are able to strongly modulate NMD efficiency, each in their own way (in a cis or trans manner), leading to the protection of their RNA (Fig. 2). This protection can be associated with one specific stop codon of the viral RNA, such as in RSV with its protective sequences (cis manner); it can also directly affect the NMD process as HTLV-1 does, and can possibly increase the availability of host transcriptional factors to increase the expression as well as the stability of viral RNA (trans manner). Additionally, when NMD is inhibited in a trans manner, it can lead to the alteration of the expression of important cellular genes. Considering both aspects, NMD inhibition is necessary for retroviral replication and to create deleterious cellular conditions, which might induce cell destruction or oncogenic transformation as well as modulation of the immune response. While at present studies in this field are limited to retroviruses, recent data with other mammalian and plant viruses tend to generalize the points we described here 132,133 and support the need for greater development of the relatively young virus/NMD field of investigation. Finally, using viruses as inhibitory tools would be of significant interest to continue deciphering the NMD process, its regulation in human cells, and the pathologies such as cancer in which its involvement has not yet been fully appreciated.
Footnotes
Acknowledgments
We are grateful to S. Garvis for critical reading of the manuscript. This study was supported by funds from the French National Research Agency (ANR), the Association de Recherche contre le Cancer (ARC), and the Association Française contre les Myopathies (AFM).
Author Disclosure Statement
No competing financial interests exist.