
Abstract
In HIV-1 infection elevated serum levels of interferon-α (IFN-α) and interleukin-10 (IL-10) are associated with immune hyperactivation and disease progression. Recently, coexpression of CD49b and LAG-3 was shown to identify Type 1 regulatory T (Tr1) cells, which secrete large amounts of the immunosuppressive cytokine IL-10. We analyzed the frequency of CD49b/LAG-3+ Tr1 cells in the peripheral blood of HIV-infected individuals at different stages of the disease. We found increased levels of CD49b/LAG-3+ Tr1 cells as well as IL-10 in HIV patients. With disease progression, Tr1 cells negatively correlate with frequency of plasmacytoid dendritic cells (pDCs), the main producers of IFN-α. However, elevated IL-10 levels could not be ascribed to the CD49b/LAG-3+Tr1 cell population. Moreover, we showed in vitro that IFN-α leads to an upregulation of IL-10 as well as CD49b/LAG-3+ Tr1 cell counts in healthy controls, recapitulating effects observed in vivo during HIV infection. Our results suggest that overexpression of IFN-α during HIV infection drives the generation of CD49b/LAG-3+ Tr1 cells and the immunosuppressive cytokine IL-10. Furthermore, it remains unclear whether elevated IL-10 levels are beneficial or detrimental in regard to disease progression.
Introduction
T
Type 1 regulatory T (Tr1) cells are an antigen (Ag)-specific adaptive subtype of Tregs, which is induced in the periphery after chronic exposure to Ag. Tr1 cells are able to reestablish tolerance in immune-mediated diseases. 3 –5 They are defined by their cytokine response, especially by their capacity to produce, among others, high levels of the immunosuppressive cytokine interleukin-10 (IL-10). 6,7 Direct characterization of those cells has been limited in the past due to lack of appropriate surface markers.
To specifically identify Tr1 cells, previous researchers have used a variety of transcriptional and membrane-bound markers. Recently, it has been demonstrated that human and murine Tr1 cells—unlike other CD4+ T cell subsets such as TH1, TH2, TH17, and Foxp3+ Treg cells—coexpress CD49b and LAG-3 (lymphocyte-activation gene 3), 8 which enables tracking of these cells.
Previous studies have shown that IL-10 is upregulated in patients with progressive HIV disease. 9,10 Moreover, the potential role of IL-10 in disease progression has also been proposed in pathogenic simian immunodeficiency virus (SIV) 11 and other chronic viral infections. 12,13 IL-10 inhibits antigen presentation and suppresses the production of proinflammatory cytokines. 14 –17 However, blockade of the IL-10 pathway enhances HIV-specific CD4+ T cell function. 10,18,19 Interestingly, it has been shown that HIV+ individuals with severely compromised T helper function produce proportionally higher levels of IL-10. 19
Similar to IL-10, elevated serum levels of interferon (IFN)-α produced by plasmacytoid dendritic cells (pDCs) are associated with HIV disease progression. 20,21 IFN-α is upregulated in response to viral infections and displays both antiviral 22 –24 as well as immune-modulating properties. 25,26 During chronic HIV and SIV infection, excessive levels of IFN-α and IFN-stimulated genes (ISG) do not correlate with viral control but are associated with immune hyperactivation, which itself is predictive for HIV disease progression. 27 –29 However, the exact role of IFN-α and ISG in the course of HIV infection remains unclear.
Interestingly, it has been proposed that IFN-α plays a crucial role in promoting IL-10 production 30 and might limit the induction of overly intense immune responses through the generation of Tregs. 31,32 Yet, the interplay of IFN-α in the induction of IL-10-producing CD49b/LAG-3+ Tr1 cells has not been defined. Our study represents the first comparative analysis of CD49b/LAG-3+ Tr1 cells as described by Gagliani et al. 8 in peripheral blood of HIV+ individuals at different stages of the disease.
Material and Methods
Study participants
The study was carried out at the Department 1 of Internal Medicine, University of Cologne, Germany. After informed consent peripheral blood (pB) was drawn from 21 seronegative controls, 34 chronic viremic HIV+ subjects of different CDC categories (named as progressors) in the absence of antiretroviral therapy, and 6 slow progressors (Table 1). Slow progressors were defined as having a documented HIV-1 infection for more than 2 years, viremia <400 copies/ml and CD4+ T cells over 500/μl, and no antiretroviral treatment (ART). Slow progressors were excluded if they had two consecutive pVL above 400 copies/ml during the observation period. The study was approved by the Institutional Review Board of the University of Cologne, Germany (09-210 and 00067/2000). All standard laboratory measurements were performed at the central laboratory of the University of Cologne. Plasma HIV RNA levels were determined using Roche Amplicor Kits (Roche Diagnostics) with a lower limit of detection of 40 copies/ml.
Values for analytical results are presented as median (interquartile range).
CDC, Centers for Disease Control. CDC A and CDC C represent a classification system of clinical category of HIV infection. A, asymptomatic HIV infection; C, symptomatic HIV infection, AIDS.
Isolation of peripheral blood mononuclear cells (PBMCs) and CD4+ T cells
PBMCs were isolated from blood by Ficoll density gradient centrifugation. CD4+ T cells were enriched by negative selection from freshly isolated PBMCs using the CD4+ T cell Isolation Kit (Miltenyi Biotec) according to the manufacturer's instructions. The purity of isolated CD4+ T cells of every sample was measured prior to each experiment.
Flow cytometry
In brief, fresh PBMCs were incubated with the following antibodies and appropriate isotype controls: the CD49b/LAG-3+ Tr1 cell population was identified according to Gagliani et al. 8 CD49b/LAG-3+ Tr1 cells were stained with anti-CD3-APC (clone BW264/56) (Miltenyi Biotec), -CD4-PE/Cy7 (clone OKT4) (BioLegend), -CD49b-FITC (clone AK-7) (BD Biosciences), and -LAG3-PerCP (Clone HAS3) (R&D Systems) antibodies. Cells were detected by gating the CD3/CD4+ population within PBMCs or isolated CD4+ T cells and further selecting CD49b/LAG-3+ cells. pDCs were identified by anti-BDCA2-FITC (clone AC144) (Miltenyi Biotec) and -CD123-APC (clone AC145).
Stained cells were measured by flow cytometry (FACSCanto1) using FACSDiva software (BD Biosciences). In all cases, 250,000–500,000 events were acquired as assessed by forward and side light scatter profile (Supplementary Fig. S1; Supplementary Data are available online at
Measurement of cytokine expression
Levels of IFN-α in serum were quantified by enzyme-linked immunosorbent assay (ELISA; PBL Biomedical Laboratories, Piscataway, NJ).
Serum levels of IL-10 were investigated via ELISA (R&D Systems) and IL-10 expression on CD49b/LAG-3+ Tr1 cells was detected with the Secretion Assay Cell Enrichment and Detection Kit (PE) (Miltenyi Biotec) after activation with anti-CD3 (clone MOPC-173) (BioLegend, San Diego, CA) and -CD28 (clone L293) (BD Biosciences) antibodies according to the manufacturer's instructions.
To investigate the effect of IFN-α on CD49b/LAG-3+ Tr1 cells, the frequency of CD49b/LAG-3+ Tr1 cells was measured after 24 h of incubation with anti-CD3/CD28 in the presence or absence of 10 ng/ml IFN-α.
Statistical analysis
GraphPad Prism (GraphPad Software, La Jolla, CA) was used to graph and analyze data for statistical significance. Data were expressed as mean and standard error of the Mean (SEM) or median and interquartile range (IQR) as indicated in the table and figure legends. Both the Shapiro–Wilk and D'Agostino–Pearson tests were performed to determine normality distribution. Significant differences were tested using the two-tailed Mann–Whitney or Student's t-test. Pearson's r was used to describe correlations.
Results
CD49b/LAG-3+ Tr1 cell frequency increases with HIV disease progression despite CD4+ T cell loss
We found higher CD49b/LAG-3+ Tr1 cell frequencies in progressors compared to healthy controls and slow progressors (Fig. 1A). The highest amount of CD49b/LAG-3+ Tr1 cells was detected in HIV CDC stage C patients (n = 14, 3.1% ± 0.64 of total CD3+/CD4+ cells) compared to CDC stage A patients (n = 20, 1.27% ± 0.23, p = 0.0203), slow progressors (n = 6, 1.01% ± 0.18, p = 0.0393), and healthy controls (n = 21, 0.82% ± 0.15, p = 0.0035) (data are expressed as mean ± SEM). As classical Treg cells, including the Tr1 cell population, represent approximately 1–10% of the mature CD4+ T cell pool 33 and CD4+ T cells decrease with disease progression, we correlated the amount of CD49b/LAG-3+ Tr1 cells to the loss of CD4+ T cells. As shown in Fig. 1B we found a negative correlation between the frequency of CD49b/LAG-3+ Tr1 and CD4+ T cells (r = −0.5898, p < 0.0001). These findings indicate that CD49b/LAG-3+ Tr1 cells, which are also CD4+, are less affected by depletion than non-Tr1 CD4+ cells. Additionally, we found a significant correlation between the frequency of CD49b/LAG-3+ Tr1 T cells and viral load (r = 0.4606, p = 0.0009; Fig. 1C).
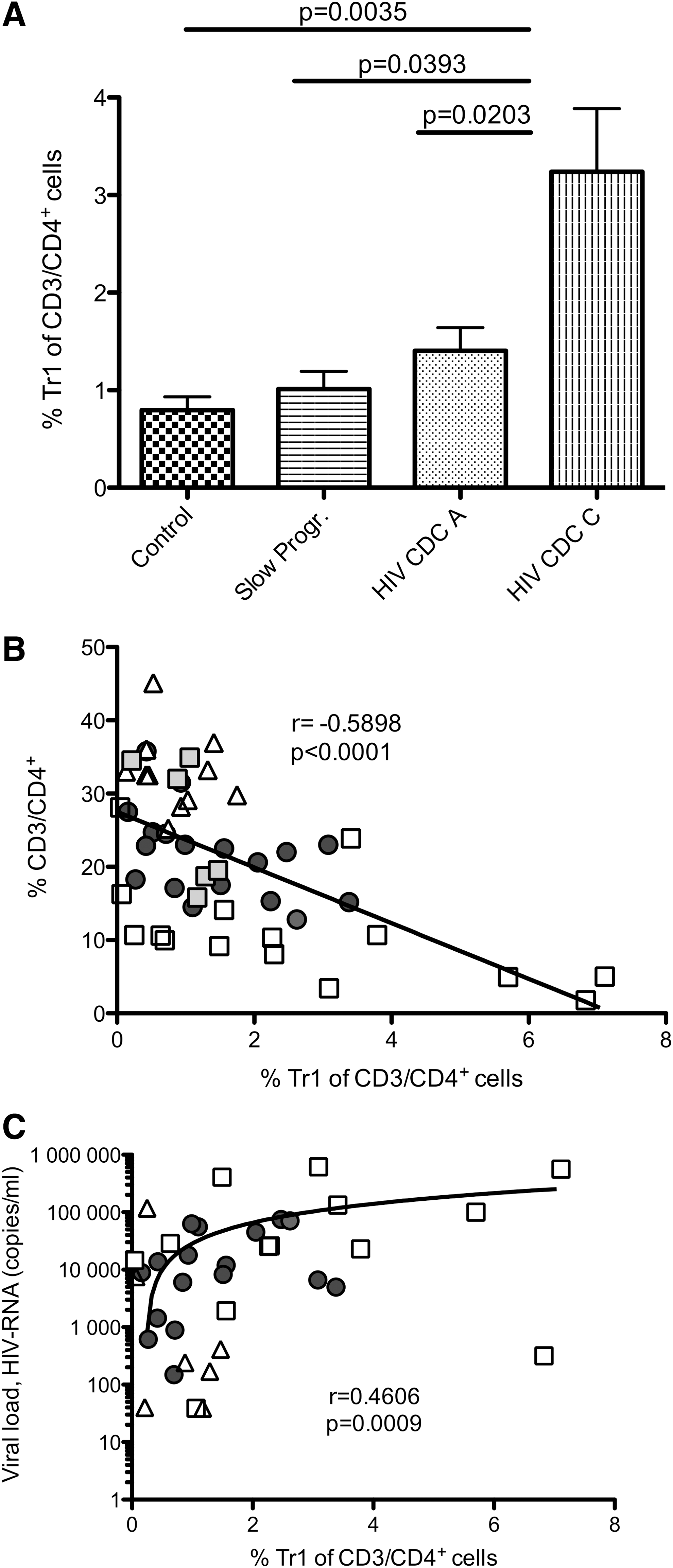
CD49b/LAG-3+ Tr1 levels are heightened in HIV+ patients and increase during HIV disease progression despite loss of CD4+ T cells.
Decreased frequency of pDCs correlates with increasing CD49b/LAG-3+ Tr1 cell counts and increasing serum IFN-α levels are associated with HIV disease progression
We were interested in pDCs as it was reported that these cells, despite their property of inducing hyperactivation, might prevent overly intense immune responses through the generation of IL-10-producing Tregs. 31,34
To reveal a possible interaction between IFN-α-producing pDCs and CD49b/LAG-3+ Tr1 cells, we measured pDC frequency in all study groups and correlated pDC with CD49b/LAG-3+ Tr1 cell frequency. We found that the frequency of circulating pDCs decreases with disease progression, and that slow progressor patients showed high pDC frequency values similar to the healthy control group [healthy controls (n = 12), 0.54% ± 0.06, versus progressors CDC stages A (n = 18), 0.31% ± 0.09 p = 0.0061, versus CDC stage C (n = 12), 0.15% ± 0.04 p = 0.0008, versus slow progressors (n = 6), 0.41% ± 0.08 p > 0.05; data are expressed as mean ± SEM]. Remarkably, the decline of pDCs during the course of HIV infection inversely correlated with CD49b/LAG-3+ Tr1 cell frequency (r = −0.3503, p = 0.0197; Fig. 2A).

Increasing CD49b/LAG-3+ Tr1 frequency negatively correlates with plasmacytoid dendritic cell (pDC) frequency during HIV disease progression and higher levels of serum interferon (IFN)-α in later stages of HIV disease.
In addition, we investigated IFN-α levels in the serum of a representative selection of patients (Fig. 2B). Progressors in CDC stage C (n = 5, 74.5 pg/ml ± 11.79) showed higher levels of serum IFN-α compared to patients in early stages of infection (n = 7, 44.38 pg/ml ± 12.61).
Recombinant IFN-α drives the generation of CD49b/LAG-3+ Tr1 cells
Based on our results in patients and on previous publications on work with IFN-α and Tr1 cells as described so far, we suggested that IFN-α also drives the generation of newly characterized CD49b/LAG-3+ Tr1 cells. Therefore, we wanted to investigate this hypothesis with special interest in the course of HIV infection. We isolated PBMCs of healthy controls (n = 5) as well as HIV+ patients (n = 6) and stimulated them for 24 h with anti-CD3/CD28 antibodies in the presence or absence of recombinant IFN-α (10 ng/ml). In healthy subjects, the presence of IFN-α resulted in an increase of the CD49b/LAG-3+ Tr1 cell frequency (3.6% ± 0.9) compared to controls without IFN-α (1.6% ± 0.86; p = 0.0401; data are expressed as mean ± SEM). We therefore concluded that in vitro IFN-α drives the generation of CD49b/LAG-3+ Tr1 cells phenotypically similar to those found in HIV+ patients. In contrast, in HIV+ patients (n = 5), recombinant IFN-α did not lead to an induction of CD49b/LAG-3+ Tr1 cells. Here, the presence of additional IFN-α did not have any further impact on CD49b/LAG-3+ Tr1 levels, which are already elevated in HIV+ individuals (−IFN-α: 5.75% ± 2.13; + IFN-α: 5.43% ± 2.4; Fig. 3).
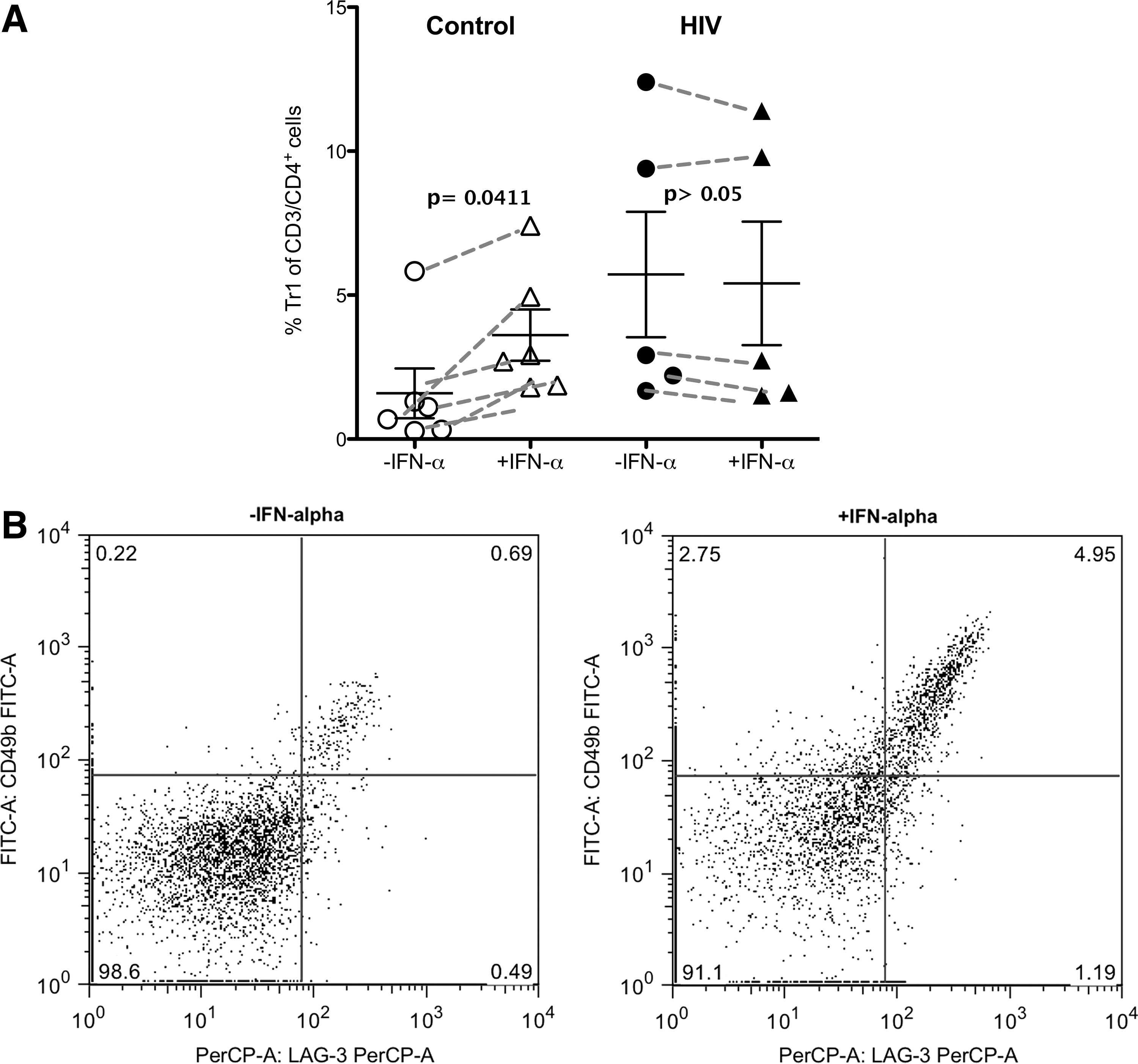
Recombinant IFN-α leads to generation of CD49b/LAG-3+ Tr1 cells in the healthy control group.
In vivo IL-10 levels are increased within the CD4+ but not within the CD49b/LAG-3+ Tr1 cell population in HIV+ patients
A number of studies reported that expression of the immune suppressive cytokine IL-10 correlates with disease progression. 19,35 We therefore measured IL-10 expression by CD4+ T cells in HIV+ patients (progressors and slow progressors) in different stages of disease by secretion assay analysis. We found that IL-10 secreted from CD4+ T cells is significantly increased in progressors (n = 12, 6.08 ± 0.71, p = 0.0043) as well as in slow progressors (n = 6, 6.33 ± 0.90, p = 0.0021) compared to the healthy control group (n = 15, 3.64 ± 0.26) (Fig. 4A). Interestingly, IL-10 levels in CD4+ T cells were significantly higher in CDC stage C patients (n = 6) compared to the CDC stage A (n = 6) group (p = 0.021). Moreover, significantly higher levels of IL-10 secretion per CD4+ T cell were detected in progressors (n = 15, 0.40 ± 0.05) and slow progressors (n = 6, 0.31 ± 0.08) compared to the healthy control group (n = 15, 0.13 ± 0.02, p < 0.001; Fig. 4B) despite HIV+ characteristic decreasing CD4+ T cell numbers.
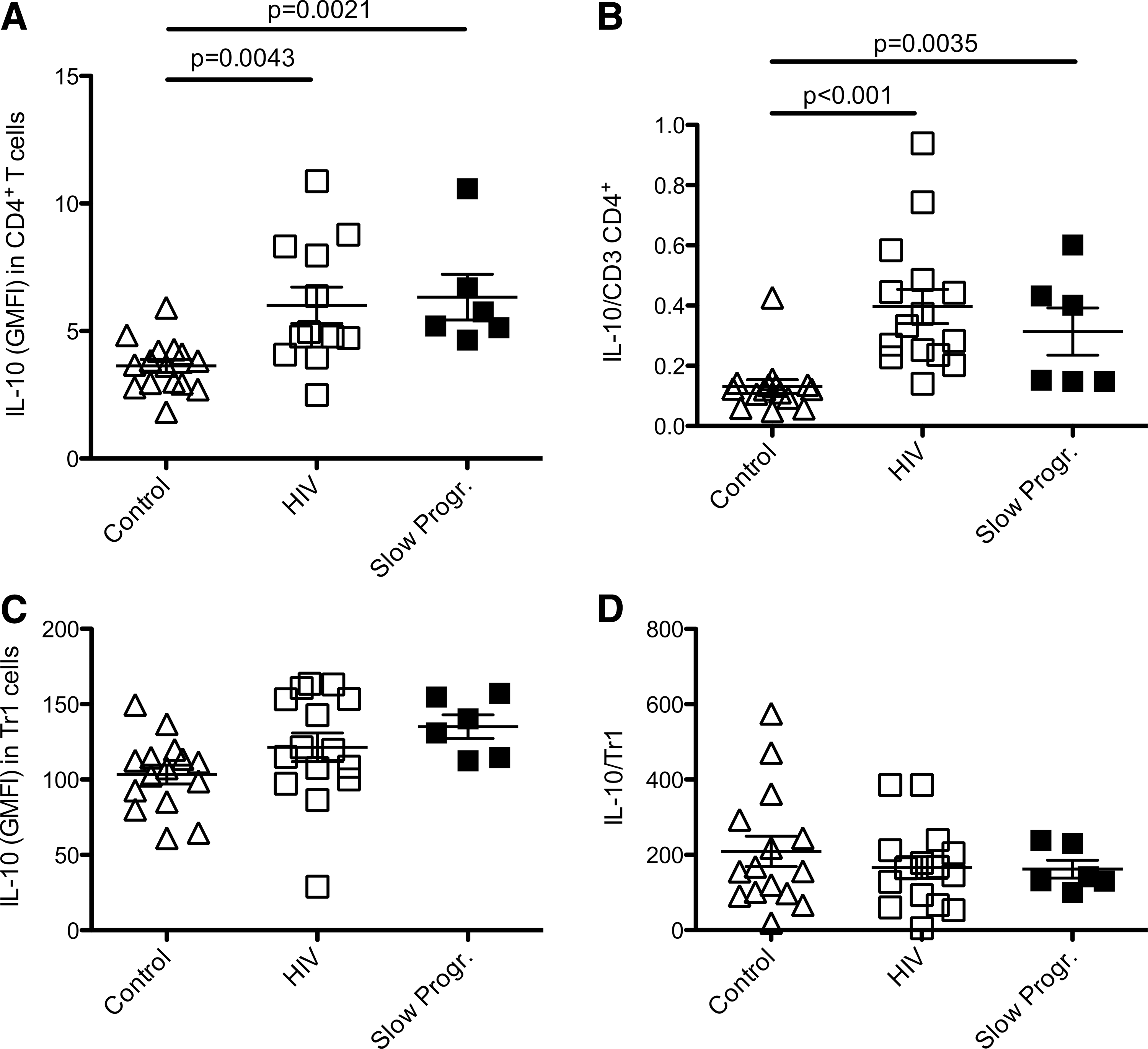
Increased interleukin (IL)-10 secretion within the CD4+ T cell population as well as the IL-10/CD4+ ratio is detected in HIV+ patients and slow progressors. IL-10 secretion within the CD49b/LAG-3+ Tr1 cell population and the ratio of IL-10 cytokine expression to CD49b/LAG-3+ Tr1 do not differ significantly between all study groups.
Our primary results on CD49b/LAG-3+ Tr1 cells (Fig. 1) showed that CD49b/LAG-3+ Tr1 cell frequency in slow progressors did not differ significantly from healthy controls. We therefore expected similar results for in vivo IL-10 levels. However, slow progressor results are not in line with the control group for IL-10 cytokine levels but resemble those of progressors. So far, published work comparing IL-10 levels in slow progressors and progressors reveals differential IL-10 levels between both groups. However, more studies are needed, as results regarding the role of IL-10 in slow progressors are not conclusive so far.
Moreover, as Tr1 cells with the newly described CD49b/LAG-3+ phenotype were shown to secrete large amounts of IL-108 we subsequently investigated whether IL-10 levels are upregulated in these cells in our study groups. Surprisingly, IL-10 secretion in CD49b/LAG3+ Tr1 cells did not differ significantly between healthy controls (n = 15, 103.4 ± 6.2), progressors (n = 15, 121.4 ± 9.5), and slow progressors (n = 6, 136.3 ± 7.2) (Fig. 4C). Additionally, progressors (n = 15, 166 ± 28.7) and slow progressors (n = 6, 153.8 ± 21.5) had lower IL-10/Tr1 cell ratios compared to healthy controls (n = 15, 209.3 ± 40.7; Fig. 4D). Our results show that CD49b/LAG-3+ Tr1 cells, as defined by Gagliani and colleagues, 8 are not the main producers of IL-10 in the context of HIV disease.
Finally, we investigated the effect of IFN-α in vitro on IL-10 production of CD4+ T cells versus CD49b/LAG-3+ Tr1 cells. First, we analyzed IL-10 levels using CD4+ T cells isolated from healthy subjects, stimulated for 24 h with anti-CD3/CD28 antibodies in the presence or absence of IFN-α. Figure 5 shows that in vitro IFN-α increased IL-10 expression by CD4+ T cells. We furthermore investigated the effect of IFN-α on IL-10 secretion specifically in CD49b/LAG-3+ Tr1 cells. As already observed in HIV+ patients, in vitro we did not observe an increase in IL-10 secretion within the CD49b/LAG3+ Tr1 cell population (Fig. 5B).

Recombinant IFN-α leads to increased IL-10 expression in CD4+ T cells but not in CD49b/LAG-3+ Tr1 cells.
Discussion
The aim of this study was to analyze the frequency of the newly described CD49b/LAG3+ Tr1 cells in HIV+ individuals and to investigate a possible association between IFN-α produced by pDCs and CD49b/LAG3+ Tr1 cells.
Treg activity has been reported to increase during several infections including HIV. 36 –40 It was shown that HIV infection directly and indirectly modulates Treg cells in vivo, as individuals with chronic HIV infection have higher frequencies of conventional Treg than individuals who control HIV or healthy subjects. 41,42 In the present report we show for the first time higher CD49b/LAG-3+ Tr1 cell frequencies in HIV+ patients with disease progression compared to slow progressors and healthy controls. Yet, it remains unclear whether the observed increase has beneficial or detrimental effects on the course of HIV infection.
On the one hand the high frequency of CD49b/LAG-3+ Tr1 cells could be harmful, as they might reduce HIV-specific responses and thus contribute to viral persistence. Previously, it has been suggested that conventional Tr1 cells could prolong pathogen persistence by suppressing protective Th1 response. 43,44 On the other hand increasing CD49b/LAG-3+ Tr1 cell levels observed in HIV infection could be beneficial by suppressing generalized chronic immune activation, especially through the inhibition of activated CD4+ T cells, one of the main hallmarks of HIV infection. However, even though CD49b/LAG-3+ Tr1 cells are elevated with HIV disease progression, they still seem to be insufficient in functioning as immune suppressors as they fail to control the hyperactivation and the ensuing loss of CD4+ T cells as already observed for the earlier described Tr1 cell phenotype. 1
A possible causative link between increased IFN-α expression, heightened CD49b/LAG-3+ Tr1 cell levels, and HIV disease progression was suggested by the fact that after ex vivo stimulation IFN-α can promote differentiation of conventional Tr1 cells as they have been described so far. 31,32 In our study, we show a decreased frequency of pDCs and increasing serum IFN-α levels in peripheral blood of HIV patients. The decreased pDC frequency correlates positively with increasing CD49b/LAG-3+ Tr1 cell counts and increasing serum IFN-α levels. Moreover, we demonstrated that in vitro recombinant IFN-α drives the generation of CD49b/LAG-3+ Tr1 cells phenotypically, similar to those found in HIV+ patients. These results support the hypothesis that elevated IFN-α levels, which are characteristic of HIV-infected individuals, might be responsible for the high CD49b/LAG-3+ Tr1 cell frequency observed in these patients, supporting the idea of thereby counteracting an overly intense immune response.
Consistent with a number of studies reporting that expression of the immune suppressive cytokine IL-10 correlates with disease progression 19,35 we showed that in vivo IL-10 levels are increased within the CD4+ cell population of HIV-infected individuals. Nevertheless, in vivo the CD4+ subgroup CD49b/LAG-3+ Tr1, as defined by Gagliani and colleagues, 8 was not the main source of IL-10 production. In vitro experiments confirmed that IFN-α increased IL-10 expression by CD4+ T cells, which parallels previous findings in CD8+ T cells. 45 However, we did not observe an increase in IL-10 secretion within the CD49b/LAG3+ Tr1 cells population in vitro.
Our results suggest that although CD49b/LAG-3+ Tr1 cells are not the main IL-10-producing cells during HIV infection, they might compensate for this inability with increasing cell numbers, which eventually leads to a general increase in IL-10levels. Yet it is not clear whether increasing IL-10 levels within the CD4+ T cell population are due to increasing numbers of IL-10-producing CD49b/LAG-3+ Tr1 cells or if IL-10 secretion in HIV+ patients is triggered within a different CD4+ T cell subgroup. Our results are in line with previous studies showing elevated IL-10 levels in HIV+ patients. 10,46,47 In addition, IL-10 has been observed in pathogenic—but not in nonpathogenic—SIV infection. 11,35 However, in the context of chronic HIV infection it does not seem that CD49b/LAG-3+ Tr1 cells, as defined by Gagliani and colleagues, 8 are the main producers of IL-10.
In conclusion, this is the first study reporting that the frequency of the recently described CD49b/LAG-3+ Tr1 cells increases in peripheral blood of HIV-infected patients (Fig. 6). Moreover, following anti-CD3/CD28 stimulation CD4+ T cells from HIV patients produced greater amounts of IL-10 than cells of uninfected donors. However, in the context of HIV infection we could not ascribe increasing IL-10 levels to the CD49b/LAG-3+ Tr1 cell population. Finally, we show that IFN-α promotes the generation of IL-10-producing CD49b/LAG-3+ Tr1 cells. These observations provide a mechanism through which excessive expression of IFN-α during HIV infection contributes to maintaining a status of elevated immune activation while at the same time suppressing protective immune responses.
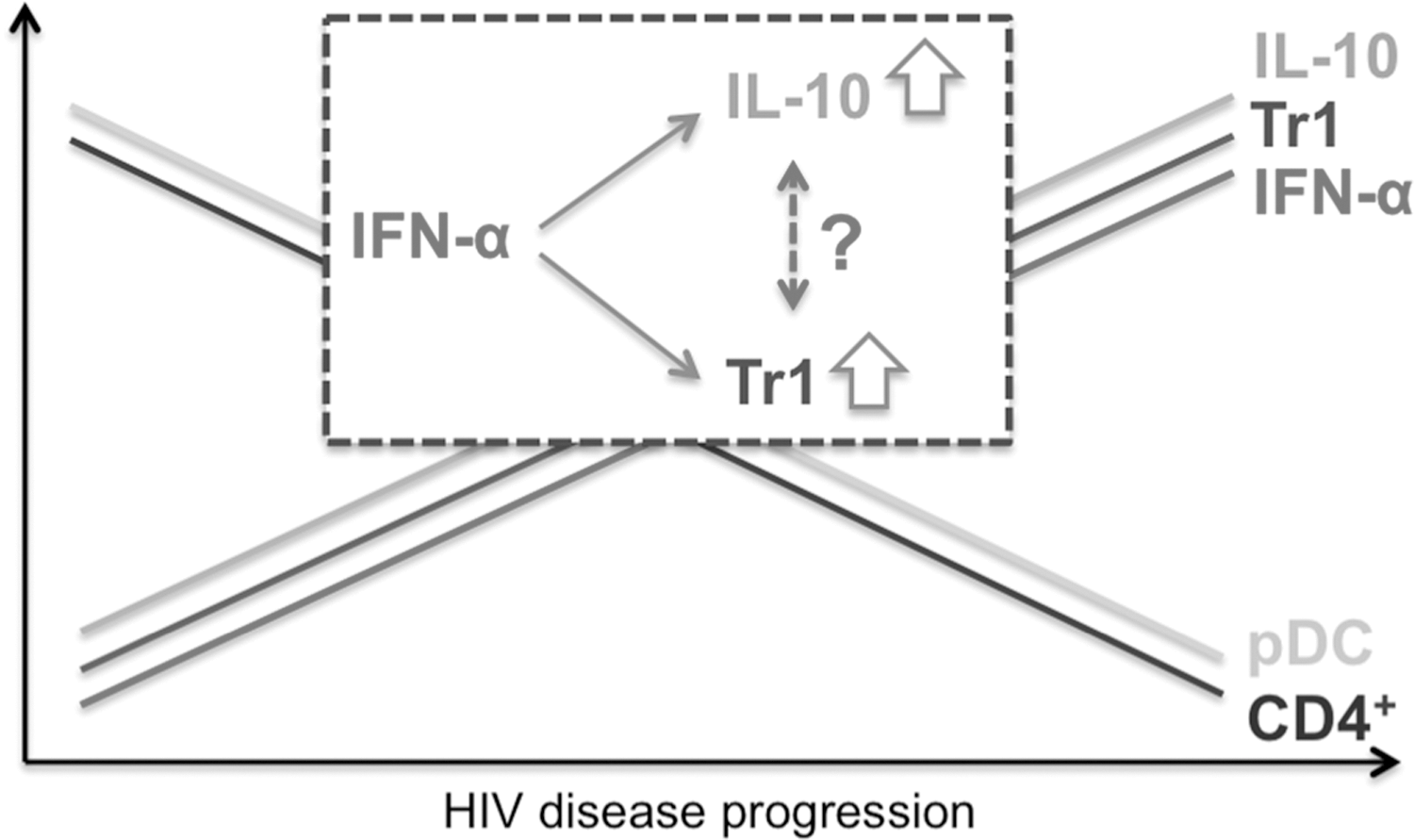
Overexpression of IFN-α during the course of HIV infection drives the generation of CD49b/LAG-3+ Tr1 cells and the immunosuppressive cytokine IL-10. While pDC and CD4+ T cell levels decrease with HIV disease progression, IFN-α, IL-10, and CD49b/LAG-3+ Tr1 cell levels increase. However, increasing IL-10 levels cannot exclusively be ascribed to the CD49b/LAG-3+ Tr1 cell population.
Footnotes
Acknowledgments
These findings were presented in part in abstract form (Abstract #P-C13) in March 2014 at the Conference on Retroviruses and Opportunistic Infections (CROI), Boston, MA. This work was supported in part by the German Centre for Infection Research (DZIF).
C.L. and K.K. designed the experiments; N.K. and U.S. performed the experiments; J.S. and N.J. discussed the results; G.F. and P.H. selected the patients and discussed the results; F.R. and G.F. contributed to the design of the study and data interpretation; C.L. and K.K. designed the study, analyzed the data, and wrote the manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.