
Abstract
Bacterial vaginosis (BV) has been linked to an increased risk of human immunodeficiency virus (HIV) acquisition and transmission in observational studies, but the underlying biological mechanisms are unknown. We measured biomarkers of subclinical vaginal inflammation, endogenous antimicrobial activity, and vaginal flora in women with BV and repeated sampling 1 week and 1 month after completion of metronidazole therapy. We also compared this cohort of women with BV to a healthy control cohort without BV. A longitudinal, open label study of 33 women with a Nugent score of 4 or higher was conducted. All women had genital swabs, cervicovaginal lavage (CVL) fluid, and cervicovaginal biopsies obtained at enrollment and received 7 days of metronidazole treatment. Repeat sampling was performed approximately 1 week and 1 month after completion of therapy. Participant's baseline samples were compared to a healthy, racially matched control group (n=13) without BV. The CVL from women with resolved BV (Nugent 0–3) had significantly higher anti-HIV activity, secretory leukocyte protease inhibitor (SLPI), and growth-related oncogene alpha (GRO-α) levels and their ectocervical tissues had significantly more CD8 cells in the epithelium. Women with persistent BV after treatment had significantly higher levels of interleukin-1β, tumor necrosis factor alpha (TNF-α), and intercellular adhesion molecule 1 (ICAM-1) in the CVL. At study entry, participants had significantly greater numbers of CCR5+ immune cells and a higher CD4/CD8 ratio in ectocervical tissues prior to metronidazole treatment, compared to a racially matched cohort of women with a Nugent score of 0–3. These data indicate that BV is associated with changes in select soluble immune mediators, an increase in HIV target cells, and a reduction in endogenous antimicrobial activity, which may contribute to the increased risk of HIV acquisition.
Introduction
B
Most women acquire human immunodeficiency virus type 1 (HIV-1) through heterosexual contact with an infected male partner. 6 Cross-sectional studies show that HIV-1 is more prevalent in women with BV than in women with normal vaginal flora. 7 –11 Observational, population-based, studies have found a two to four times increase in the odds of acquiring HIV-1 among women with incident BV. 12 –16 An observational study of serodiscordant couples found that HIV-1-positive women with BV were more likely to transmit the virus to their male partners. 17 The association between BV and HIV-1 acquisition has led to HIV-1 prevention efforts that include prophylactic screening and treatment of BV, but the results of these efforts have been mixed. 18 –20
It is hypothesized that BV functions as a cofactor in HIV-1 transmission by altering epithelial integrity, genital immune cell populations, and local mucosal immune responses. 21,22 However, few studies have directly evaluated these potential mechanisms. Therefore, we followed 33 women diagnosed with BV or intermediate vaginal flora by Nugent score 23 longitudinally and obtained cervicovaginal biopsies and genital samples in the presence of BV or intermediate vaginal flora (Nugent 4–10) and approximately 1 week and 1 month after completing metronidazole therapy.
The objective of the study was to determine how BV and its treatment were related to ectocervical tissue lymphocyte populations, soluble immune and inflammatory mediators, and the endogenous antimicrobial activity of genital tract secretions. We specifically focused on the anti-HIV activity and the anti-Escherichia coli activity of cervicovaginal lavage (CVL) fluid because the former may contribute to an innate host defense against HIV
24
–27
and the latter has been associated with a healthy microbiome.
28,29
We also compared these 33 women with BV at baseline to a racially matched cohort of women, enrolled in a separate CONRAD study (CONRAD D11-119,
Materials and Methods
This study was approved by the Chesapeake Institutional Review Board (IRB) (Pro #00006122) with a waiver of oversight from the Eastern Virginia Medical School (EVMS) and was registered in
At the screening visit (V1), BV or intermediate vaginal flora was confirmed by a Nugent score of 4 or higher. 23 Patients with a qualifying Nugent score had additional screening tests performed including a pap smear (Thin Prep, Hologic, Bedford, MA) and screening for additional reproductive tract infections (RTIs) including Neisseria gonorrhoeae (NG) and Chlamydia trachomatis (CT) (Gen-Probe, Aptima, San Diego, CA). Point-of-care testing was performed to exclude pregnancy, recent vaginal semen exposure (ABAcard, Abacus Diagnostics, West Hills, CA), HIV-1 infection (Oraquick Advance, Orasure Technologies, Bethlehem, PA), and Trichomonas vaginalis (TV) infection (OSOM, Genzyme, Cambridge, MA). Women with no exclusion criteria identified at V1 underwent genital tract sampling on the day of the screening visit, prior to antibiotic therapy.
After biological specimens were obtained at V1, all women were provided with 14 500-mg metronidazole tablets and were instructed to take the tablets, with food, twice daily for 7 days. The first dose was administered in the clinic under direct observation. Participants then returned 7–10 days after completing a 7-day course of metronidazole therapy for visit 2 (V2) (14–17 days after visit 1) and 28–32 days after completing treatment for visit 3 (V3) (35–39 days after visit 1). Compliance with metronidazole therapy was assessed by returned pill counts and participant report. Patients were instructed to abstain from intercourse for 48 h prior to each visit. If BV was not resolved by V3, participants were retreated with either oral metronidazole or metronidazole vaginal gel.
We did not delay metronidazole treatment to synchronize visits to the phase of the menstrual cycle because (1) participants were symptomatic and seeking treatment for BV and (2) data support that the vaginal flora fluctuates from day to day, particularly in women with non-Lactobacilli-dominated flora. 30 The day of the menstrual cycle was recorded at each visit. Participants were considered to be in the luteal phase, as opposed to the follicular phase, of the menstrual cycle if they were in the last 14 days of their menstrual cycle.
At all three visits, the following samples were obtained: vaginal semen testing (ABAcard, West Hills, CA) (a negative result was required per the protocol), vaginal pH, gram stain for Nugent score, and semiquantitative vaginal flora culture. A CVL was performed with 10 cm3 of normal saline. After the CVL and vaginal swabs were obtained, topical benzocaine gel was applied to the cervicovaginal tissues and three full-thickness biopsies (one ectocervical and two vaginal) were obtained using a Tischler biopsy forcep. Monsel's solution and pressure were applied to the biopsy sites to control bleeding. Patients were instructed to place nothing in the vagina for 5 days after the biopsy procedures.
Semiquantitative vaginal cultures
Two Dacron swabs were used to collect vaginal fluid, which were placed in a Port-A-Cul transport tube (Becton, Dickinson, Sparks, MD) and transported on ice to the laboratory of S. Hillier, PhD (Magee-Womens Research Institute, Pittsburgh, PA) within 24 h of collection for quantitative culture analysis. The swabs were used to inoculate the following media: Brucella sheep blood agar (Hardy Diagnostics, Santa Maria, CA), Columbia sheep blood agar, laked-blood kanamycin agar, two human bilayer Tween agar (Becton, Dickinson, and Company, Sparks, MD), Rogosa agar, A-8 agar, and Mycoplasma broth (prepared in house). The Columbia blood agar, A-8, Mycoplasma broth, and one human blood tween (HBT) plate were incubated at 37°C in 6% CO2 for 48 h. The remaining media was incubated at 37°C within an anaerobic chamber (Anaerobe Systems, Morgan Hill, CA) for a minimum of 4 days. The aerobic and anaerobic organisms were identified to the genus, species, or groups of bacteria using phenotypic tests, Gram stain and colony morphology, and aerotolerance as described in the Manual of Clinical Microbiology. 31 The quantity of bacterial growth was graded as 0 (no growth on plate) to 4 (growth in all four quadrants of the plate), a scale that has been correlated to log concentrations of bacteria. 32 Lactobacillus species were tested for hydrogen peroxide production in a qualitative assay on tetramethylbenzidine agar plates. 33
Antimicrobial activity of the CVL
Within 30 min of collection, the CVL was centrifuged at 4°C for 10 min at 500×g. Aliquots of CVL supernatant were stored at −80°C and shipped to the laboratory of B. Herold, MD at Albert Einstein College of Medicine for testing of the antimicrobial activity of the CVL and the laboratory of R. Fichorova, MD, PhD at Brigham and Women's Hospital/Harvard Medical School for testing of secretory leukocyte protease inhibitor (SLPI), soluble intercellular adhesion molecule-1 (sICAM-1), cytokines, and chemokines in CVL.
The activity of CVL against HIV-1 and E. coli was measured within 12 months of collection, as previously described. 34 For anti-HIV activity, TZM-bl cells were cultured in 96-well dishes overnight. The cells were infected with HIV-1BaL (approximately 103 TCID50) mixed 1:1 with CVL or control buffer (normal saline containing 200 μg/ml of bovine serum albumin). After 48 h incubation at 37°C, the inoculum was removed by washing, cells were lysed with the addition of luciferase cell culture lysis reagent (Promega, Madison, WI), and the plates were stored at −80°C until assessed for luciferase activity, which was measured in relative light units (RLU). Mock-infected cells served as a negative control and wells in which tenofovir (100 μg/ml) was added served as an internal positive assay control. TZM-bl inhibition was measured as mean percent reduction compared to control. All samples were tested in triplicate in at least two independent experiments.
To assess the anti-E. coli activity, bacteria (ATCC strain 4382627) were grown overnight to stationary phase and then 3 μl of bacteria (∼109 cfu/ml) was mixed with 27 μl of CVL or control buffer (20 mmol/liter potassium phosphate, 60 mmol/liter sodium chloride, 0.2 mg/ml albumin, pH 4.5) and incubated at 37°C for 2 h. The mixtures were further diluted in buffer (to yield 800–1000 colonies on control plates) and plated in duplicate on agar enriched with trypticase soy broth. Colonies were counted using ImageQuant TL v2005 after an overnight incubation at 37°C. Results are presented as the mean percentage inhibition relative to colonies formed on control plates. Bacteria cultured with penicillin/streptomycin served as an internal positive assay control and consistently showed 100% inhibition of colony-forming units (cfu).
Cytokine and chemokine concentrations in the CVL supernatant
SLPI was measured in CVL samples by ELISA (R&D Systems, Minneapolis, MN) using a Victor2 reader (Perkin Elmer Life Sciences, Boston, MA). For initial screening, all samples were diluted 80-fold in the sample diluent provided by R&D Systems, and all samples showing levels above the assay detection range were additionally diluted to obtain accurate measurements. Interleukin (IL)-1β, IL-6, IL-8, IL-10, ICAM-1, and tumor necrosis factor alpha (TNF-α) were measured in undiluted CVLs by a multiplex electrochemiluminescence (ECL) assay (Meso Scale Discovery, Gaithersburg, MD). All measurements were performed in duplicate. A split quality control pool prepared from CVLs was tested on each plate showing an SLPI interplate coefficient variation of 21.1% and between 1.8% and 12.6% for all markers on MSD multiplex plates, and intraplate variation <16% (6.4%±8.0%) for SLPI and <10% for all markers on MSD mutiplex plates.
Analysis of ectocervical immune cell populations and histology
For the immunohistochemistry (IHC) analyses, one ectocervical biopsy from each visit was placed in 10% neutral buffered formalin for 24–48 h. After the completion of fixation, the tissues were transferred to an embedding cassette and processed overnight in a Tissue-Tek V.I.P, Vacuum Infiltration Processor (E150 Series) (Sakura Finetek, Torrance, CA). The infiltration program involves a scheduled sequence of solutions [from PBS buffer, through an increasing percent grades of ethanol solutions, to xylene, and finally, to paraffin at (58°C)]. After the completion of paraffin processing, the cassettes were removed and the tissue was placed in fresh paraffin on a metal mold and allowed to cool and harden.
Paraffin-embedded tissue blocks of the ectocervical biopsies were cut into 5-μm sections. IHC staining of tissue sections was performed using the ABC method [avidin:biotinylated enzyme complex from Vector laboratories (Burlingame, CA)]. Briefly, the slides were deparaffinized, dehydrated, and rehydrated followed by antigen retrieval in citrate buffer (pH 6.2, DAKO) at high temperature. Nonspecific binding was blocked using a specific protein block for 30 min at room temperature. After washing with phosphate-buffered saline (PBS), the slides were incubated overnight with primary antibody at 4°C. The slides were then washed with PBS and subjected to biotinylated secondary antibody followed by ABC reagent. The antigen was localized by incubation with AEC chromogen–substrate (skyTek labs, Mississauga, Ontario, Canada) and finally mounted with Accergyl mounting media (Accurate Chemicals, Westbury, NY) with a coverslip. Positive stained cells were either counted under the microscope (Nikon E-800) or analyzed using Image J software (NIH, Bethesda, MD), as described below.
Cell phenotype was identified using specific monoclonal antibodies against CD45, CD3, CD8, CD1a, CD68, CCR5, CD4, and HLA-DR. Cell density was expressed per high-power area (cells/HPA). Tight junction proteins E cadherin (rabbit monoclonal, Abcam, Cambridge, MA) and desmoglein 3 (DSG3) (mouse monoclonal, Invitrogen, Grand Island, NY) were stained with monoclonal antibodies. For these markers, the immunolabeling of the protein was analyzed semiquantitatively using Image J software (NIH, Bethesda, MD). In brief, five to six areas were randomly selected using a Nikon E800 microscope from each section and these images were captured using a CCD camera (Spot Camera, Diagnostic Instruments, Sterling Heights, MI). The integrated optical density (IOD) in each area was calculated for the positive staining color. The IOD value of the negative control (no primary antibody) was subtracted from the IOD values for each tissue and the mean value was calculated for the areas of each tissue sample.
Subset analysis of immune cell populations among a control population of healthy women without BV
A subset analysis was performed, comparing the ectocervical tissue immune cell phenotype and activation status obtained from participants at V1, prior to metronidazole treatment, compared to the immune cell phenotype and activation status of racially matched women in another CONRAD study (CONRAD D11-119,
Statistical analyses
Descriptive statistics include mean, median, standard deviation, and range for continuous variables and frequencies and percentages for categorical variables. The principal of inference in this study was the changes of the main outcomes across three different visits. We used SAS PROC MIXED to analyze the data by fitting mixed regression models with a repeated statement to account for the within subject correlation. All models were adjusted for the effect of race (non-Hispanic black, non-Hispanic white versus mixed race).
Comparisons of ectocervical immune cell phenotype and activation status among women in this study, compared to women without BV in the CONRAD D11-119 study, was done using the Wilcoxon–Mann–Whitney test (PROC NPAR1WAY). For the analyses of cytokine and chemokine data, calculations were done with natural log transformation, to account for variability in these endpoints. Statistical calculations were performed in SAS (version 9.3, SAS Institute Inc., Cary, NC). Statistical significance was determined at the level of alpha=0.05. We performed univariate linear regression analysis using the anti-HIV activity of the CVL as the dependent variable and each of the study endpoints as independent variables, in an attempt to determine which biological mechanisms may correlate with anti-HIV activity. After confirming univariate models, we grouped the study endpoints by possible biological mechanisms: immune cells (CD45, CD3, CD8, CD4, CCR5, HLA-DR, CD1a, CD68), cytokines and chemokines (IL-1B, IL-8, TNF-α, IL-10, SLPI, ICAM-1), and semiquantitative levels of bacterial species (Gardnerella, Ureaplasma, Myoplasma, and H2O2-producing lactobacilli) and performed repeated measured mixed regression model analysis to determine which endpoints were correlated with anti-HIV activity.
Results
Figure 1 details the number of women who were screened, enrolled, and provided complete follow up-data. Sixty-nine women with symptomatic vaginal discharge were screened; 35 had a Nugent score of 4 or higher and underwent genital tract sampling at V1. Of the 35 patients sampled at V1, 33 completed all three visits (n=99 samples) and represent the population for data analysis. Demographic and medical history variables for the 33 participants are outlined in Table 1.
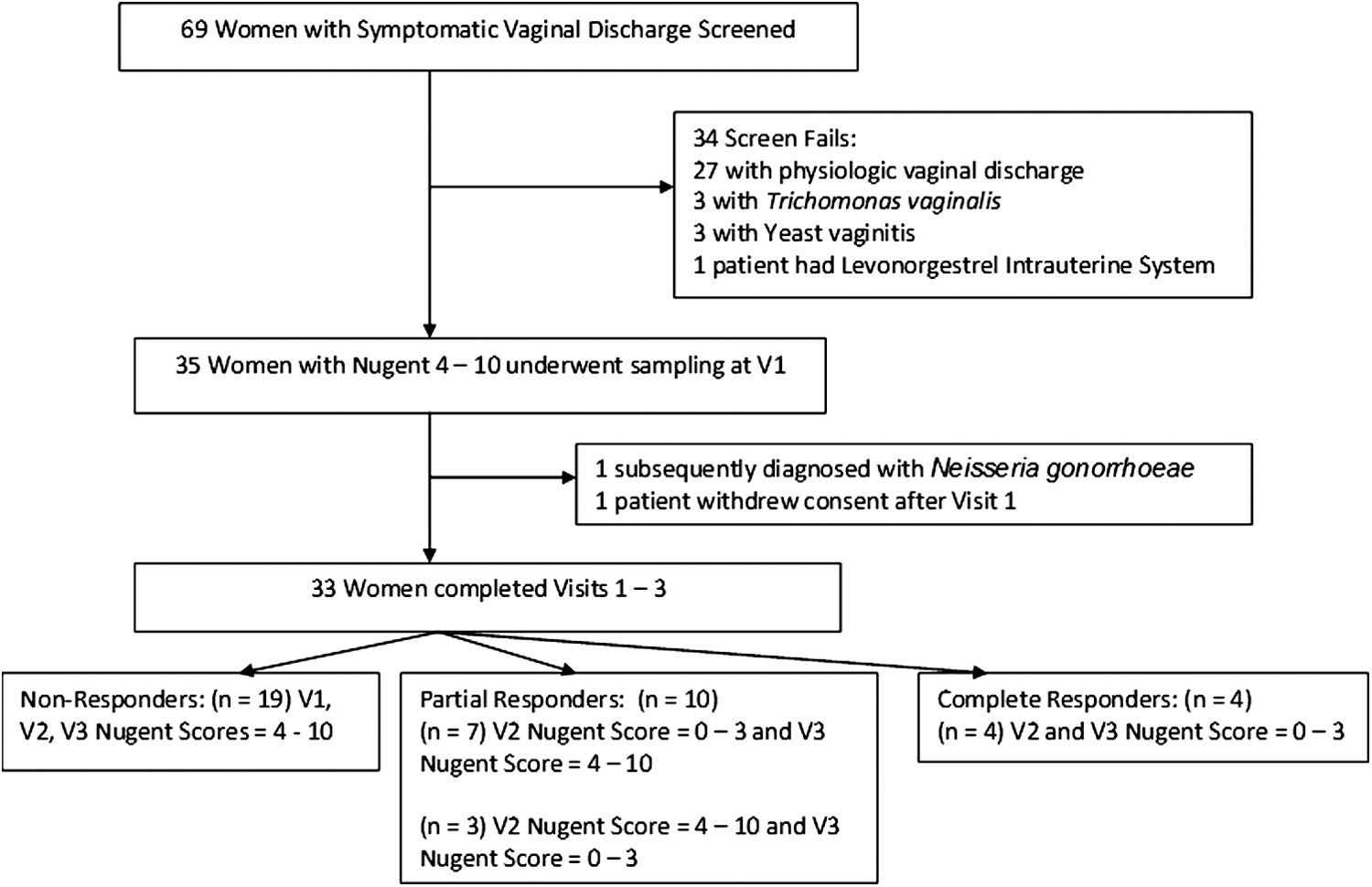
Study Population for CONRAD Protocol D11-115 (
BMI, body mass index; HSV, herpes simplex virus.
Thirty participants had an initial Nugent score of 7–10 and 3 women had an initial Nugent score of 4–6. Following metronidazole treatment, four women responded with normal Nugent scores at V2 and V3; 10 were partial responders and either had a normal Nugent score at V2 but not V3 (n=7) or at V3 but not V2 (n=3). The other 19 women had persistently abnormal Nugent scores at both V2 and V3 and are classified as nonresponders. Notably, among the three women with an initial Nugent score of 4–6 at V1, two of these women had Nugent scores of 7 or higher at V2 and V3, while the remaining woman had a Nugent score of 1 at V2 and a Nugent score of 8 at V3.
Visit 1 was more commonly conducted in the luteal phase of the menstrual cycle (p = <0.01), compared to visit 2. There were no significant differences in the mean menstrual cycle day or the proportion of samples obtained in the luteal phase between visits 1 and 3 or between visits 2 and 3, based on repeated measure analyses. There were no significant differences in the mean age, body mass index (BMI), years of education, using male condoms for birth control, phase of the menstrual cycle at the time of treatment, and number of sexual partners in the last month (all p values>0.05, data not shown) between women who were nonresponders (n=19) versus the 14 women who achieved at least one normal Nugent score at V2 and/or V3. Women who reported their ethnicity as non-Hispanic black or mixed race were significantly more likely to be nonresponders (16/27, 59.2%) and (3/3, 100%), respectively, compared to women who identified themselves as non-Hispanic white (0/3 0%) (Fisher exact test p=0.02). We therefore controlled for race (non-Hispanic black, non-Hispanic white, or other) in the subsequent analyses.
Changes in biomarkers based on visit: effect of metronidazole therapy
In the analyses of the effect of metronidazole treatment on study endpoints (Table 2), we controlled for race and categorized the Nugent score as normal (0–3) versus intermediate (4–6) versus BV (7–10), using a repeated measures model. Following metronidazole treatment, women experienced significant, acute increases in the anti-E. coli activity of CVL (p=0.02) and a significant decrease in semiquantitative levels of vaginal Mycoplasma species (p=0.04) at V2. However, the levels of both of these endpoints decreased and returned to levels similar to baseline by V3. Reflecting the high recurrence rates, there were no significant differences in the quantity of hydrogen peroxide (H2O2)-producing lactobacilli or Gardnerella species based on the visit.
CVL, cervicovaginal fluid.
In terms of soluble inflammatory mediators in the CVL supernatant, 1 week after metronidazole therapy, women experienced significant increases in IL-6, IL-8, and IL-10 at V2 (all p values<0.05). All these mediators then decreased significantly by V3, returning to baseline V1 levels.
In the analyses based on visit, there were no significant differences in immune cell phenotype or activation markers in ectocervical tissue epithelia or lamina propria (CD45, CD3, CD8, CD1a, CD68, CCR5, CD4, HLA-DR) or markers of genital epithelial integrity (E cadherin and DSG3) between samples obtained at V1 versus V2 versus V3, controlling for race and categorizing the Nugent score, in paired comparisons (all p values>0.05, data not shown).
Analysis of posttreatment samples based on Nugent score
Because BV did not resolve in the majority of participants, we next analyzed the 66 samples obtained posttreatment, at V2 (n=33) or V3 (n=33), based on the Nugent score, controlling for race in a repeated measures model (Table 3).
The endogenous antiviral activity of the CVL against HIV was significantly increased in samples obtained from women with normal flora (Nugent 0–3), compared to women with BV (Nugent 7–10) (p=0.02). Most importantly, women with BV (Nugent 7–10) had mean anti-HIV activity approaching 0% inhibition, indicating that their CVL provided no defense against HIV infection of TZMbl cells in vitro; these data are represented in Fig. 2a.
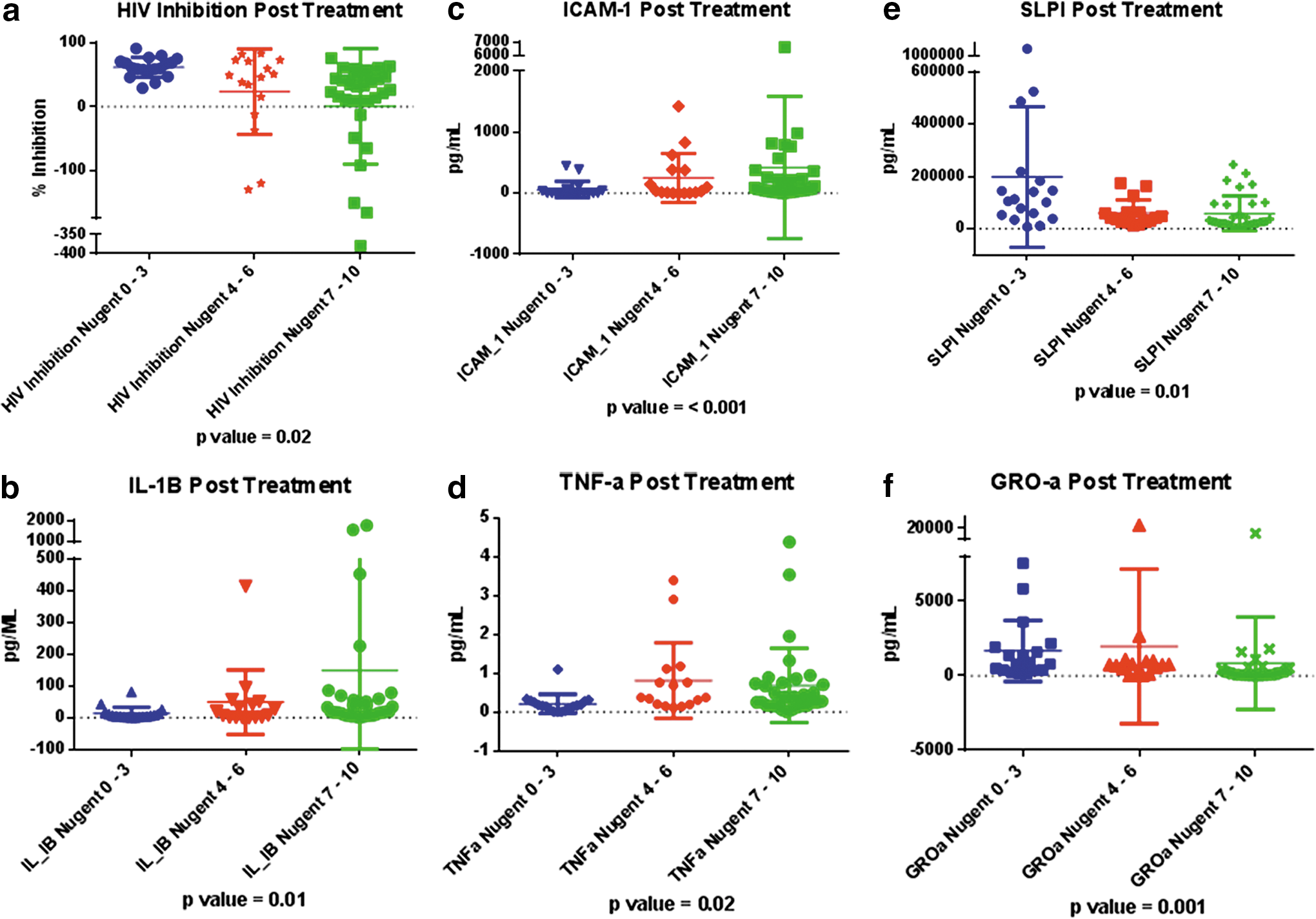
IL-1β, TNF-α, and ICAM-1 levels were significantly elevated in samples obtained posttreatment from participants with persistent intermediate flora or BV compared to samples obtained in the presence of normal flora (p<0.05). SLPI and GRO-α levels were significantly decreased among samples obtained posttreatment in the setting of BV or intermediate flora, compared to women with normal flora (p=0.02) (Fig. 2b–f and Table 3). As expected, the quantity of Gardnerella species was significantly higher in samples obtained posttreatment with a Nugent score of 7–10 or 4–6 compared to samples obtained after resolution of BV (p=<0.01). Finally, ectocervical samples obtained posttreatment in the presence of normal vaginal flora had significantly higher numbers of CD8 cells in the epithelium compared to samples obtained from participants with persistent BV (p=0.03).
Table 4 demonstrates differences in ectocervical immune cell phenotype and activation status among the 33 participants in this study at V1, prior to metronidazole therapy, compared to 13 racially matched (10 non-Hispanic black women and 3 non-Hispanic white women) (Fisher exact test=0.07) women who had baseline biopsies in the follicular and luteal phases of the menstrual cycle (26 samples).
Contraceptive use and/or sexual practices among these 13 control participants were as follows: condom use (n=4), abstinence (n=4), fallopian tube occlusion or partner's vasectomy (n=3), and same-sex relationship (n=2). We did not query participants in the CONRAD 119 study regarding the number of sexual partners in the past month or vaginal douching practices. In this control population, we found no significant differences in immune cell populations between the follicular (menstrual cycle days 7–13) and luteal phases of the menstrual cycle (cycle days 18–26). At the time of these biopsies in the 119 study, a vaginal swab was obtained and the Nugent score was 0–3 for all control participants at each visit. The mean Nugent score of this subset was 1.12±1.45 compared to the mean Nugent score of the BV study participants at V1 (8.3±1.6) (p<0.001).
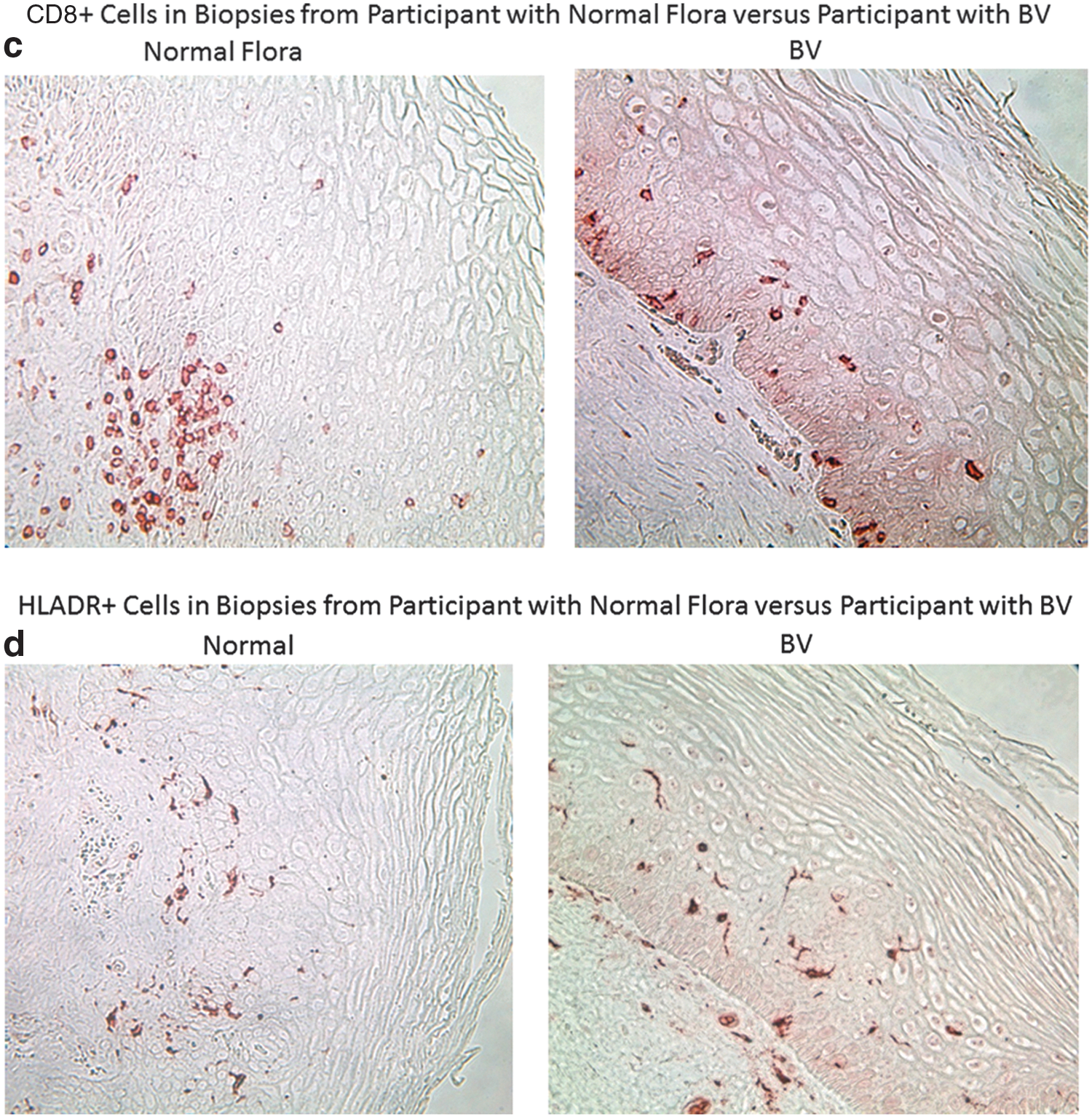
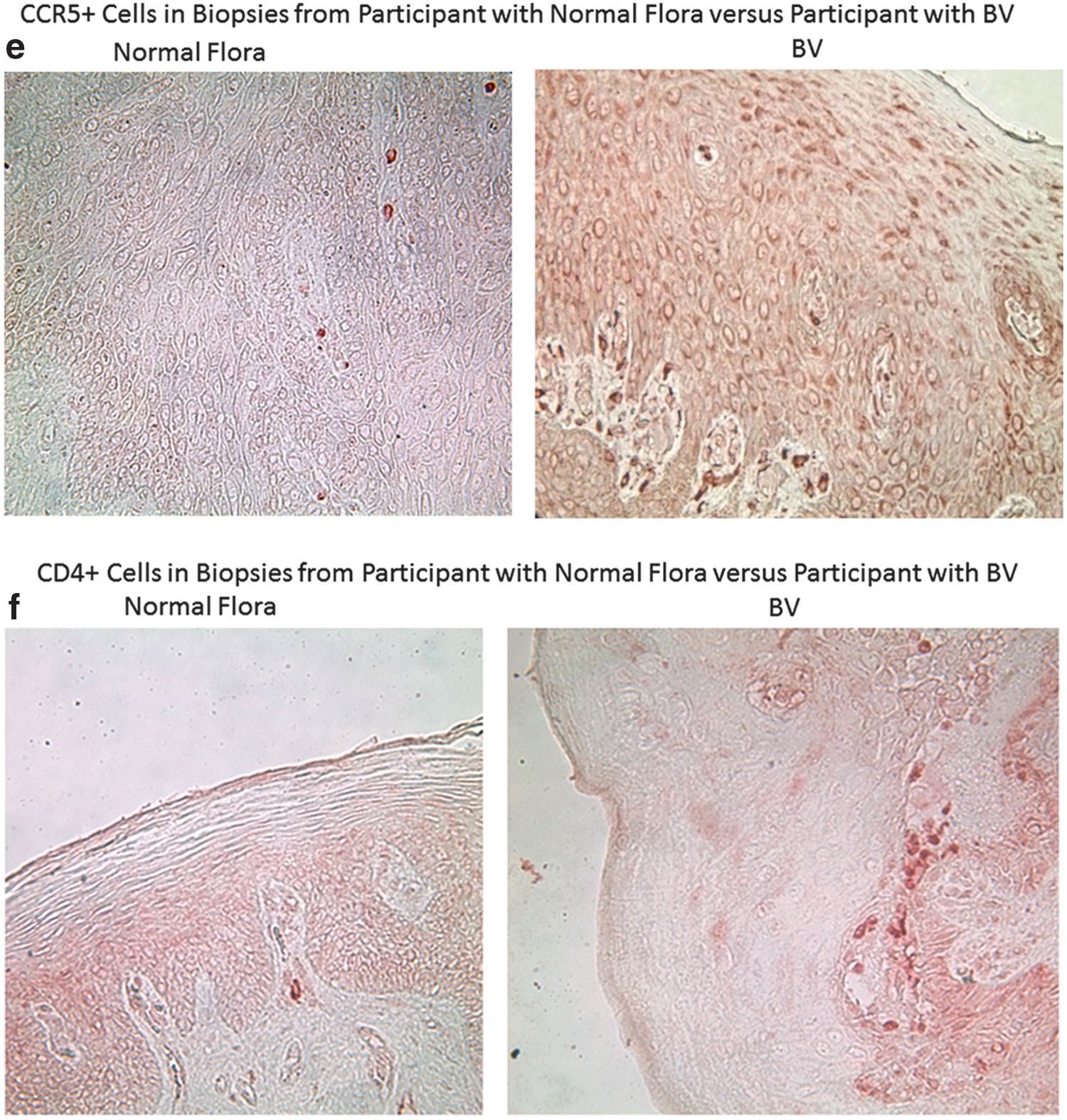
The vaginal pH of the women with normal vaginal flora (4.15±0.20) was also significantly lower than the vaginal pH of the BV study participants at V1 (5.09±0.3) (p<0.001). We found that control participants in the CONRAD 119 study, with normal vaginal flora, had significantly higher numbers of CD45+, CD3+, and CD8+ cells in the genital epithelium and lamina propria (p values<0.05). In addition, women with normal flora had significantly higher numbers of HLA-DR + immune cells in the ectocervix. Women with abnormal vaginal flora showed a trend toward higher numbers of CD4+ cells (p=0.12) and significantly more CCR5+ cells than women with normal flora (p=0.02). Finally, the ratio of CD4/CD8 cells in the lamina propria was significantly higher among participants enrolled in the BV study compared to controls (p=0.02). Representative images for immune cell populations for control participants enrolled in the CONRAD 119 study versus BV study participants at visit 1 are shown in Fig. 3a–f.

We performed univariate mixed model analysis with visit as a random effect to control for repeated measures at different times, using the anti-HIV activity of CVL as the dependent variable and each of the study endpoints as independent variables, in an attempt to determine which biological mechanisms may correlate with anti-HIV activity. After confirming univariate models, we grouped the study endpoints by possible biological mechanisms: immune cells (CD45, CD3, CD8, CD4, CCR5, HLA-DR, CD1a, CD68) and soluble cytokines, chemokines, and immune mediators (IL-1β, IL-8, TNF, IL-10, SLPI, ICAM-1, GRO-α, Elafin, MIP-3α, RANTES).
Using univariate models, we found that CVL levels of Elafin (p=0.02), GRO-α (p=0.0002), MIP-3α (p=<0.001), and RANTES (p=0.01) were significantly correlated with the anti-HIV capacity of the CVL in the mixed-effects model.
In multivariate models, we found that increasing levels of IL-8 (p=0.0006) and MIP-3α (p=0.01) and decreasing levels of IL-1β (p=0.0003) were significantly associated with increasing anti-HIV activity of the CVL.
Discussion
Among the novel findings of this clinical study is that persistent BV posttreatment was associated with a significant reduction in the anti-HIV activity of CVL compared to the CVL obtained from women posttreatment who achieved a normal Nugent score (Table 3). Our data support several biological processes that are likely associated with a significant reduction and/or absence of anti-HIV activity associated with BV.
At study entry, participants had significantly higher numbers of CCR5+ immune cells in the genital epithelium compared to racially matched women with normal vaginal flora (Table 4). An expansion in CCR5+ cells is a key alteration of the genital mucosa that could potentially enhance susceptibility to HIV-1 infection. 35 –37 This finding is in accordance with a previous study of 15 HIV+ women, 8 of whom were on antiretroviral therapy, which showed that successful resolution of BV was associated with a significant reduction in the number of endocervical CD4+ T cells, including those expressing the CCR5 coreceptor and the activation marker CD69, in endocervical cytobrush specimens. 38
At baseline, participants also had a significantly higher ratio of CD4+ to CD8+ immune cells in the ectocervical lamina propria compared to controls with normal flora (Table 4). They had significantly fewer genital leukocytes (CD45+ cells) and immune cells bearing CD3, CD8, and HLA-DR in the ectocervical epithelium and lamina propria compared to samples obtained from healthy controls (Table 4). These findings are in accordance with a previous cross-sectional study, 39 which found that women with BV had significantly fewer genital tract leukocytes, measured by flow cytometry, compared to women with normal flora and women with yeast vaginitis, which is classically associated with overt inflammatory symptoms. 40
Participants with persistent BV after metronidazole therapy had significantly fewer CD8 cells in the ectocervical epithelium as compared to women with resolved BV (Nugent score 0–3) (Table 3). In addition to being more prevalent than CD4+, CD8+ T cells in the cervicovaginal tissue have a higher expression of activation markers and an increased effector memory phenotype. 41 A sequestered population of CD8+ tissue-resident memory T (T RM) cells appears to be common at skin and mucosal surfaces, including that of the vagina, and can be recalled by antigen-specific and nonspecific stimuli. 42,43 The presence of CD8+ T cells in the vaginal epithelium appears to be protective against various pathogens, 44 as CD8+ T cells coexpressing the transcription factor Foxp3 can also exert suppressive functions through a regulatory T cell phenotype (Treg) 45 and the suppression of these cells in women with persistent BV may signal a reduced capacity to respond to immunological threats.
Consistent with the high recurrence rates of BV, 5 only four women cleared their BV at V2 without a recurrence at V3 and therefore most samples obtained in the presence of a normal Nugent score came from women who experienced a recurrence of BV during the study. We would ultimately need to biopsy large numbers of women frequently and over long periods of time to determine how quickly cervicovaginal immune cells changed in response to alterations of vaginal flora. Our data indicate that our participants with BV at V1, who all had previous episodes of symptomatic BV and had high recurrence rates despite metronidazole therapy, had genital immune cell milieus that were significantly different from healthy comparators without chronic BV infections (Table 4).
Although the number of non-Hispanic white women in this study was small (n=3), they were significantly more likely to resolve their BV than women who reported their race as non-Hispanic black or mixed. These findings are consistent with other studies that found that the incidence and prevalence of BV and nonlactobacilli predominant flora are higher among non-Hispanic black women. 2,46 –48 Therefore, we controlled for race in the statistical analyses.
In contrast to immune cell populations, we found that genital tract cytokines and chemokines changed significantly based on both metronidazole treatment (Fig. 2b–f and Table 2) and posttreatment, depending on the Nugent score (Table 3). Specifically, there were significant increases in IL-6, IL-8, and IL-10 1 week after women received metronidazole therapy. Possible explanations for the increase in proinflammatory mediators associated with treatment include the fact that metronidazole would preferentially target anaerobic organisms, potentially selecting out BV-associated, pathogenic aerobic species, 49 or that inflammatory responses may have been triggered from dying bacteria. 50 It is not clear whether the modest but statistically significant increases in immune mediators in response to antimicrobial treatment are biologically relevant and whether or how this would impact the risk of HIV-1. Of note, the anti-HIV activity of the CVL did not change significantly in response to metronidazole treatment (Table 2).
In accordance with other studies, we found that samples obtained in the presence of abnormal vaginal flora (Nugent 4–10) had significantly higher concentrations of IL-1β 38,51 –60 (Table 3). It is thought that these proinflammatory cytokines, along with the presence of BV-associated bacteria, can up-regulate local HIV replication through activation of the long terminal repeat promoter region of the virus. 61 –66
IL-8 is a potent chemotactic and activating factor for neutrophils in vaginal fluid 52 and vaginal levels of IL-8 correlate with inflammatory tissue influx of CD45+ and CD3+ cells. 67 IL-8 can be produced in response to factors other than IL-1β. 54,68 One group postulated that factors secreted by the microbial flora of BV, such as sialidases, blunt the IL-8 response despite elevations in IL-1β and that this leads to the hallmark of BV, a lack of overt inflammatory symptoms and vaginal fluid neutrophils despite evidence of local mucosal inflammation. 52,54
CVL obtained after treatment, with persistent abnormal vaginal flora, had significantly higher concentrations of ICAM-1, a member of a family of adhesion proteins that serve as the counterreceptor for leukocyte function-associated antigen-1 (LFA-1) 69 (Table 3). ICAM-1 is highly expressed in epithelial cells, 70 including HIV-infected CD4+ T lymphocytes. 71 It has been shown that soluble ICAM-1 levels produced by human vaginal, ectocervical, and endocervical epithelial cells are increased by proinflammatory cytokines as well as by infection with STI pathogens such as N. gonorhoeae and T. vaginalis. 72 –74 Elevated levels of ICAM-1 have been demonstrated with in vitro application of proinflammatory compounds, 69 or after in vitro infection of VK2E6E7 cells with NG, 72 and it is considered a biomarker of inflammatory response. 69 Data indicate that antibodies against ICAM-1, secreted by transformed lactobacilli, blocked cell-mediated transepithelial HIV-1 transmission in vitro. 75
CVL TNF-α levels were also significantly higher among participants with persistently abnormal vaginal flora after treatment, compared to women who resolved their BV (Table 3). In vitro treatment with TNF-α has been shown to induce robust inflammatory responses by human vaginal, ectocervical, and endocervical epithelial cells 72 and to increase HIV-1 transmission by Langerhans cells. 76
In accordance with other investigators, we found that CVL samples obtained in the presence of abnormal vaginal flora had significantly lower levels of SLPI 51,77 –81 (Table 3). A previous study of hormonal contraceptive users found significantly reduced levels of SLPI in the CVL among women who subsequently acquired HIV-1. 82
CVL from women with BV also had significantly reduced levels of growth-related oncogene-alpha (GRO-α), which is a novel finding of this study (Table 3). GRO-α secretion recruits neutrophils, 83 supporting our findings of reduced tissue leukocytes among participants with BV, and may support the idea that BV suppresses the host's ability to fend off additional pathogens.
Previous studies have shown that the anti-HIV activity of normal vaginal fluid was dependent upon the composition of cationic polypeptides, and that the anti-HIV activity was reduced or lost when these components were selectively removed from collected vaginal fluid. 24 Using multivariate regression, we found that the combination of MIP-3α, IL-8, and IL-1β concentrations was most predictive of the anti-HIV activity of the genital tract secretions.
In vitro models demonstrated that normal human vaginal fluid confers intrinsic anti-HIV capacity against both R5 and X4 strains of HIV, 24 but there are no longitudinal data on differential in vivo anti-HIV activity of the cervicovaginal secretions, based on vaginal flora. Most studies examining in vivo anti-HIV and antibacterial activity examine healthy women, 24,29,34,84,85 and exclude women with common reproductive tract infections, such as BV.
Previous groups found that the successful treatment of BV normalizes cervicovaginal immune and inflammatory markers and restores the antibacterial properties of the cervicovaginal fluid. 51,86,87 Our data are in accordance with a previous study of women with BV that showed that the inherent anti-E. coli capacity of the cervicovaginal secretions was enhanced after antimicrobial treatment 51 ; however, among our cohort, there was a high recurrence rate, with anti-E. coli activity diminishing back to baseline levels at the final visit.
The strengths of our study were that all enrolled women had normal menstrual cycles of 21–35 days, no concomitant STIs, normal cervical cytology, were not using any hormonal birth control, and were all nonsmokers, as some of these factors could potentially modify study endpoints. 56,88,89
The limitations of this study include the fact that adherence to antimicrobial regimens was based on self-report. We assume that women took the provided medication, as they were all seeking relief from bothersome vaginal symptoms, but recognize that self-report of pill taking is open to recall and social desirability reporting bias. Only 14 of 33 women had at least one normal Nugent score during either of the two follow-up visits (contributing 18 samples). This is consistent with the high recurrence rates of BV despite adequate antimicrobial therapy. 5 It would have been ideal to be able to follow women with the type of detailed genital sampling we performed for longer periods of time, but this was not feasible in this study. Because many women with BV have frequent recurrences, 5 we were not able to follow this cohort of 33 women for several months, to capture resolution of BV in the majority of women, and therefore we elected to also include a control cohort of healthy, racially matched women who were enrolled in a separate study with similar exclusion criteria.
Although all women enrolled in this study had normal menstrual cycles and were not taking exogenous hormones, we were unable to time the visits to the menstrual cycle. All participants presented seeking treatment for BV and we thought they would not want to delay treatment. Importantly, the vaginal flora, especially among women with non-Lactobacilli dominant biomes, 30 fluctuates rapidly and we wanted to obtain genital samples and initiate treatment when a diagnosis of BV or abnormal vaginal flora was confirmed.
In summary, we demonstrated that the inherent anti-HIV activity of the cervicovaginal secretions is significantly reduced in the presence of abnormal vaginal flora. CVLs obtained in the presence of abnormal flora were found to have significantly higher levels of IL-1β and ICAM-1 and significantly lower levels of SLPI and GRO-α. Multivariate regression analysis found that the soluble mediators IL-1β, IL-8, and MIP-3α significantly predicted the anti-HIV activity of the CVL, among this group of women. Finally, we found that women with BV had significantly higher numbers of CCR5+ immune cells, a known receptor for HIV-1 mucosal infection in women. These data are important because they provide insight into the biological mechanisms behind the observational data that women with BV, a highly prevalent, recurrent, and often asymptomatic infection, are at greater risk of HIV-1 acquisition.
Footnotes
Acknowledgments
The authors acknowledge Maurizio Macaluso, MD, DPH and Jennifer Legardy-Williams, MPH, CCRP for their essential contributions to the design of this study.
This work was funded by an intraagency agreement between the Centers for Disease Control and Prevention (CDC), United States Aid and International Development (USAID), and CONRAD (GPO-A-00-08-00005-00). The views expressed by the authors do not necessarily reflect those of the funding agency or CONRAD.
Author Disclosure Statement
No competing financial interests exist.