
Abstract
Mucosal tissues are the predominant sites for genital HIV-1 transmission. We investigated the mechanisms by which broadly neutralizing antibodies (bNAbs) inhibit HIV-1 replication in a coculture model including primary mucosal dendritic cells (DCs), such as Langerhans cells, interstitial dendritic cells, and CD4+ T lymphocytes. We show that bNAbs efficiently prevent HIV-1 infection by inhibiting HIV-1 transmission to CD4+ T lymphocytes. This inhibition of cell-to-cell transmission was observed with equal potency as the inhibition of cell-free infection of primary CD4+ T lymphocytes. In addition, a decrease in HIV-1 replication in DCs and the induction of DC maturation were detected. This additional inhibition was Fc mediated as it was blocked by the use of specific anti-FcγR monoclonal Abs. The DC maturation by bNAbs during HIV transmission may contribute to mucosal protection. Therefore, multiple antibody-mediated inhibitory functions should be combined for the improvement of future preventive/therapeutic strategies to cure HIV.
G
We used a physiologically relevant model of HIV-1 transmission including various primary DCs to mimic early mucosal HIV-1 infection, as previously reported. 15,16 Primary LCs and iDCs were generated from CD34+ stem cells as previously described 15 ; monocyte-derived dendritic cells (MoDCs) were isolated and differentiated from human peripheral blood mononuclear cells (PBMCs) of healthy blood donors by positive selection using CD14 MicroBeads selection kits and autoMACS (Miltenyi Biotec), respectively. 16 The primary DCs were incubated for 2 h with 50–200 ng/ml of primary HIV-1BaL isolate (NIH, Bethesda, MD). After extensive washing to remove unbound free viral particles, autologous, PHA (2 μg/ml)/IL-2 (0.1 μg/ml)-activated CD4+ T cells, purified by positive selection using CD4 MicroBeads after CD14+ purification, and anti-HIV-1 bNAbs VRC01 (directed against HIV Env gp120, kindly provided by Dr. John R. Mascola, NIH, Bethesda, MD) or 4E10 (against Env gp41, obtained from Polymun Scientific, Austria) were added to the HIV-1-loaded DCs. The percentages of infected cells and CD83+/CD86+ maturation markers were determined by flow cytometry after 72 h, and IFN-α multisubtype secretion (PBL Interferon Source, Piscataway, NJ) was quantified by ELISA (Fig. 1A).
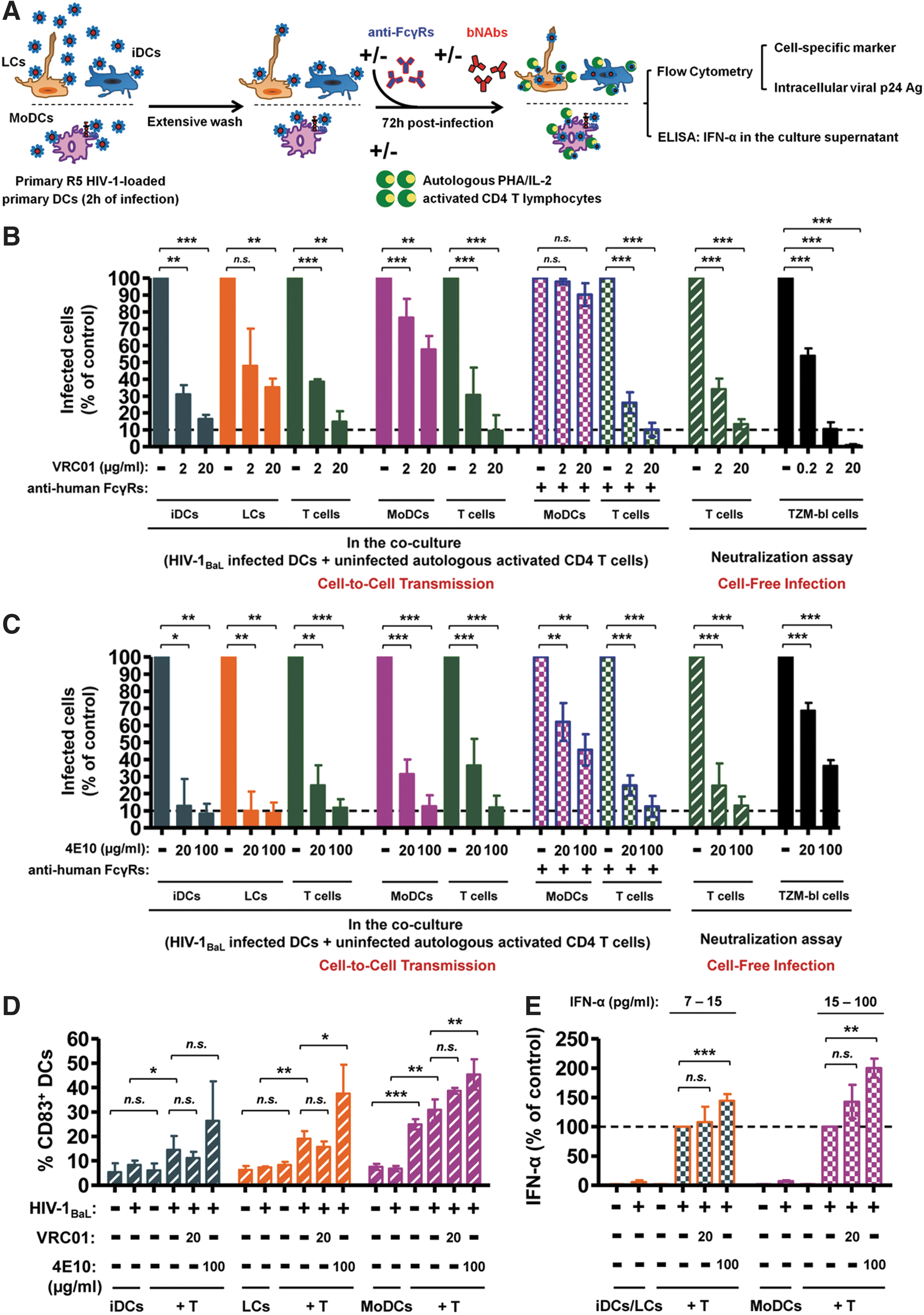
Inhibition of HIV-1 cell-free and cell-to-cell transmission by HIV-1-specific broadly neutralizing antibodies.
We assessed the inhibitory activity of bNAbs VRC01 and 4E10 using various inhibitory assays. VRC01 at a concentration of 20 and 2 μg/ml and 4E10 at a concentration of 100 and 20 μg/ml efficiently inhibited the transfer of PBMC-derived HIV-1BaL primary isolate from DCs to autologous activated CD4+ T cells (Fig. 1B and C, green columns). Interestingly, VRC01 and 4E10 inhibited with a similar efficiency the infection of CD4+ T cells from the same donor by cell-free HIV-1BaL in a conventional neutralization assay when normalized to display equivalent percentages of infected T cells (Fig. 1B and C, hatched green column). A similar bNAb-mediated inhibition was also observed with HIV-1BaL or a transmitted/founder (T/F) primary isolate HIV-1Bx11 either transferred from pDCs or directly infecting CD4+ T cells. 17
Noteworthy, using the TZM-bl assay (NIH, Bethesda, MD), 18 a 10-fold more efficient inhibition of the cell-free HIV-1BaL infection was measured for bNAb VRC01 whereas 4E10 performed poorly against this primary isolate on TZM-bl cells (Fig. 1B and C, black column). The surprisingly poor inhibition of bNAb 4E10 on the infection of TZM-bl cells by primary isolates was previously documented. 19 These results emphasize the complexity of bNAb inhibition, which depends on the target cells, the virus, and the host cells used to propagate it. This makes a direct comparison between inhibitions of cell-free versus cell-to-cell HIV-1 transmission difficult. 20,21 Further clarification of the role of bNAbs in the physiologically relevant mucosal tissues will be crucial for future vaccine and intervention strategies to prevent transmission.
DCs have been described as partially restricting HIV-1 replication and DC maturation further decreases their infection rate. 16 Early after cross-talk with autologous lymphocytes, the susceptibility of HIV-1 replication in immature DCs increases. 15,22 Surprisingly, we observed that VRC01 and 4E10 decreased DC infection, although they were added 2 h after HIV-1BaL (Fig. 1B and C). LCs, iDCs, or MoDCs express various Fcγ receptors (FcγRs) on their surfaces, 15,16 while CD4+ T cells or the TZM-bl cells lack these receptors. 23
To investigate the involvement of FcγRs on the mechanism of inhibition of DC infection by bNAbs, we blocked these FcγRs by addition of 10 μg/ml of monoclonal Abs directed against human FcγRI, FcγRII, and FcγRIII (BD Pharmingen) 16 to HIV-1BaL-loaded MoDCs concomitantly with bNAbs (Fig. 1B and C). We found that the blockade of FcγRs completely abrogated the anti-HIV-1 activity of VRC01 on MoDCs (Fig. 1B), indicating that the inhibition of DC infection by VRC01 involves FcγRs present on the surface of these cells. Moreover, the blocking FcγRs significantly impaired HIV-1 inhibitory activity of 4E10 on MoDCs (p = 0.0057 at 100 μg/ml and p = 0.0218 at 20 μg/ml of 4E10) (Fig. 1C). This impairment of 4E10 inhibitory activity is only partial, most likely because of the postfusion inhibitory potential of 4E10. 24,25 Indeed, we previously showed that about 80% of HIV-1 particles had not yet fused 2 h after addition of virus to immature MoDCs, 16 suggesting that the virus is still accessible to the postfusion inhibitory activity of 4E10. Of note, no changes in the inhibition of HIV-1 transfer to autologous activated CD4+ T cells that lack FcγRs were observed in the presence of antihuman FcγRs blocking Abs (Fig. 1B and C).
DC-T cell coculture stimulates HIV-1 replication in DCs and this stimulation was previously attributed to DC–T cell cross-talk. 22,26 Interestingly, the addition of antihuman FcγRs blocking Abs themselves reduced by 50% the stimulation of HIV-1 replication observed in cocultured MoDCs (data not shown). This reduction of HIV-1 replication in DCs may be due to the hindrance of DC–T cell cross-talk by FcγRs blocking Abs. Additional investigations will be necessary to define the role of Fc receptor-mediated adhesion, signal transduction pathways and its effect on immunological synapse formation between DCs and T cells. Taken together, these results fully corroborate the key role of FcγRs in bNAb-mediated antiviral inhibition of DC infection and of DC-to-T cell HIV-1 transmission in vitro 15,16 and possibly in vivo. 27,28
As triggering of FcγRs induces DC activation, we further assessed whether HIV-1-specific bNAbs activate DCs in our cell-to-cell HIV transmission assay. The expression of the DC maturation marker CD83 and costimulation marker CD86 was measured on the surface of HIV-1-infected DCs. We found that the expression of CD83 and CD8617 was increased in the presence of HIV-1BaL and HIV-1BX11 17 when DCs were cocultured with CD4+ T lymphocytes, and that these maturation markers were further increased by the addition of bNAb 4E10 (Fig. 1D). This maturation process could contribute to the impairment of HIV-1 replication in DCs as mature DCs are less susceptible to R5 virus replication. 16,29 DC maturation was enhanced for 4E10 at 100 μg/ml (p < 0.05) (Fig. 1D). This increased maturation is partially reverted after FcγRs blockade (data not shown) demonstrating that the binding of 4E10 to FcγRs on DCs was partially responsible for their maturation. Noteworthy, bNAb 4E10 was particularly efficacious and displayed high inhibitory activity on DC infection in our coculture model (Fig. 1C). Additional studies will be necessary to determine if this high inhibitory activity of 4E10 on DCs is due to other functions such as its polyspecific properties or its capacity to bind to phospholipids. 30,31 In contrast, bNAb VRC01 at 20 μg/ml did not induce significant DC maturation, suggesting the involvement of additional mechanisms of Fc-mediated inhibition.
In addition to DC activation markers, we measured interferon (IFN)-α secretion in the supernatants by ELISA (Fig. 1E). We found that IFN-α secretion by infected DCs was significantly increased in the presence of autologous CD4+ T cells and this effect was maintained or even enhanced in the presence of bNAbs VRC01 or 4E10, respectively (Fig. 1E); this production was donor dependent. IFN-α is known to inhibit early stages of the HIV-1 replication cycle. 12,32,33 A synergistic effect between IFN and antibodies was described for enterovirus 70 and adenovirus. 34 Whether such synergistic activity occurs in HIV has to be further analyzed. Altogether, the findings indicate that the increased DC maturation and persistence of IFN-α production in the presence of bNAbs may participate in the overall protection of cell-to-cell HIV-1 transmission.
In conclusion, we demonstrate for the first time that bNAbs not only inhibit cell-to-cell HIV-1 transmission, but also participate in the overall inhibition by decreasing HIV-1 replication in mucosal DCs and enhancing DC maturation and IFN-α production. These additional inhibitory functions involving the Fc domain of the antibody open a new area of research on Fc domain engineering (including an increase in serum half-life or modification of the affinity of the Fc region for various Fc receptors). Therefore, at the mucosal portal of HIV-1 entry, a complex network of virus–cell and cell–cell interactions induces multifunctional antibody inhibitory activities. These multiple antibody activities should be taken into consideration for the design of vaccine strategies to prevent early virus dissemination at mucosal frontlines, to contribute to protection against sexual transmission of HIV-1, and to create the next generation of antibody-based therapeutic agents. 35 –37
Footnotes
Acknowledgments
We thank Dr. John R. Mascola (NIH, Bethesda) and Polymun who kindly provided bNAb VRC01 and 4E10, respectively. We thank our financial support from the ANRS (French National Agency for Research on AIDS and Viral hepatitis), VRI (Vaccine Research Institute), EuroNeut41 (FP7-HELTH-2007-A-201038), Sidaction, Dormeur Investment Service Ltd., and Fonds de Dotation Pierre Berge. B.S. was supported by a French fellowship from ANRS.
B.S. and C.M. conceived the study, designed the experiments, and analyzed the data. B.S., M.P., C.D., and J.P. performed the experiments; L.M.M., G.L., S.S., and T.D. contributed to reagents and materials; and B.S. and C.M. wrote the article. All authors read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.