
Abstract
Acquired immune deficiency syndrome (AIDS) is mainly caused by infection with human immunodeficiency virus-1 (HIV-1) and still poses a global threat for which we lack a protective or therapeutic vaccine. Dendritic cells (DCs) play a major role in the onset of HIV infection, providing one of the primary sites of HIV replication, and also act as viral reservoirs in vivo. Previous studies have shown that baculovirus (BV) induces strong host immune responses against infections and malignancies. In this study, we infected human monocyte-derived DCs with recombinant BV (AcCAG-gag) and showed that AcCAG-gag-infected human DCs underwent maturation and produced interferon alpha and other proinflammatory cytokines accompanied by increases in the mRNA and protein expression levels of APOBEC3 (A3A, A3F, and A3G), proteins associated with the inhibition of HIV-1 replication. Surprisingly, HIV-1 inhibition is also observed in human DCs infected with a wild-type BV, as determined by the production of inflammatory cytokines, the expression of A3, and a reduction in the p24 level. Our findings outline the mechanism underlying the inhibition of HIV-1 in BV-infected human DCs and pave the way for the use of BV as an effective tool for immunotherapy against HIV-1.
Introduction
H
Dendritic cells (DCs) are the professional antigen-presenting cells of the immune system responsible for the initiation and regulation of immune responses against various antigens via the activation of T cells, natural killer cells, and B cells. 2 –5 Moreover, DCs are also involved in the transmission and pathogenesis of HIV-1, which can preferentially infect and replicate in DCs. 6 HIV-infected DCs account for the transfer of HIV-1 to CD4+ T cells and also act as viral reservoirs for long-term transmission in vivo. 7,8 Although DCs are susceptible to HIV-1, viral replication is generally less productive in DCs than in CD4+ T cells, presumably because of the presence in DCs of cellular restriction factors, such as apolipoprotein B mRNA-editing catalytic polypeptide (APOBEC3), cyclophilin A, and SAMHD1, which exhibit direct antiviral activity.
Among these cellular antiviral factors, the APOBEC3 proteins exhibit the most powerful antiretroviral activity by deaminating retroviral cytidine to uridine residues during reverse transcription, which causes hypermutation in the nascent retroviral DNA and restricts viral replication through increased degradation of the DNA and the generation of nonfunctional proviral genomes. 9 Furthermore, APOBEC3 can disrupt the propagation of HIV through direct binding with viral RNA, interfering with the DNA strand transfer during reverse transcription, physically blocking reverse transcriptase, and obstructing integration into the host cell genome. 10 –13 In addition to the function of virion-packaged APOBEC3 proteins to inhibit HIV-1, there is increasing evidence to indicate that cellular A3 proteins in resting CD4+ T cells, monocytes, and mature DCs can also restrict incoming virus particles. 14,15 Among the APOBEC3 family members, APOBEC3G (A3G) and APOBEC3F (A3F) are reported to be the potent inhibitors of HIV-1 replication in monocyte-derived DCs. 16 Recent studies have shown that APOBEC3A (A3A) is also involved in the suppression of HIV-1 replication. 17,18 Peng et al. showed that the silencing of A3A renders monocytes vulnerable to HIV-1, which associates A3A with anti-HIV activity. 19 Treatment with interferon alpha (IFN-α) significantly enhances the expression of APOBEC3 at the transcriptional level and promotes anti-HIV-1 activity. 20,21
The baculovirus Autographa californica nuclear polyhedrosis virus (AcNPV) is an enveloped insect virus carrying a circular double-stranded DNA genome of 130 kb. Viral vectors derived from this baculovirus are commonly used as tools for efficient gene transfer and recombinant protein expression. Several research groups have reported that recombinant baculovirus containing an appropriate eukaryotic promoter is able to infect and express recombinant proteins efficiently in a wide variety of mammalian cells and animal models. 22 –26 Moreover, this baculovirus has reportedly been used for the transfer of protective genes in solid organs during transplantation procedures and gene therapy against cancers. 27,28 The advantageous characteristics that prompt its use in therapeutic applications include its low cytotoxicity in mammalian cells, even at a high multiplicity of infection (MOI), its inability to replicate in mammalian cells, and the absence of preexisting antibodies against baculovirus because it is not infectious to humans. The prevalence of antiviral immunity against human viruses may deter the use of these viral vectors in gene therapy due to the possibility of undesired rejection responses.
We have previously reported that the indicated baculovirus induces strong host immune responses against infections and malignant tumors by activating both innate and adapted immunity. 29 –34 The baculovirus can penetrate into mammalian cells via envelope glycoprotein (gp64)-mediated endocytosis. The mannose receptor, which is primarily expressed on DCs, recognizes mannose present in gp64, is involved in the process of endocytosis, and plays a major role in host defense to induce innate immunity. The internalization of baculovirus DNA into endosomes occurs via the membrane fusion of viral envelope glycoproteins and clathrin-mediated endocytosis and/or macropinocytosis. 35 –41
In the present study, we examined the effect of the recombinant baculovirus AcCAG-gag (rBV) and wild-type AcNPV (wBV) on the activation of human monocyte-derived DCs and their efficacy to inhibit HIV-1 replication. rBV-infected DCs (rBV-DCs) show phenotypic and functional maturation, present substantially upregulated expression of cellular APOBEC3, and allow the inhibition of HIV-1 replication. The inhibition of HIV-1 replication was also observed in wBV-infected human monocyte-derived DCs (wBV-DCs), as determined by the production of inflammatory cytokines, the expression of A3, and a reduction in the p24 level.
Materials and Methods
Cell culture and reagents
Spodoptera frugiperda (Sf9) insect cells were cultured at 27°C in Sf-900 II culture medium (Invitrogen, Carlsbad, CA). Human monocyte-derived DCs were prepared from peripheral blood mononuclear cells (PBMCs; purchased from KAC Co., Ltd., Kyoto, Japan). PBMCs were isolated by density gradient centrifugation using Ficoll-Paque PLUS (GE Healthcare UK Ltd., Buckinghamshire, UK) and dish-adhered monocytes were cultured for 6 days in DC culture medium consisting of RPMI 1640 (Sigma Aldrich, St. Louis, MO) supplemented with 2 mM
Preparation of baculoviruses and R5-HIV-1
Wild-type baculovirus (wBV) was purchased from BD Biosciences (San Diego, CA) and propagated in Sf-9 cells in Sf-900 II culture medium. AcCAG-gag (rBV) encoding the HIV-1-gag gene was produced and propagated in Sf9 cells as previously described. 42,43 The virus titer was determined by plaque assay. The M-tropic R5-HIV-1JR-CSF strain was generated by the transfection of HEK-293T cells with plasmid DNA (pJR-CSF). The culture supernatant was collected 48 h after transfection and filtered through 0.45-μm filters to remove any residual cellular contamination. The viral p24Gag content was measured using a Lumipulse automated chemiluminescent enzyme immunoassay (CLEIA, FUJIREBIO, Tokyo, Japan) analyzer.
Baculovirus infection of human DCs and analysis
Human DCs were seeded at a density of 106 cells/well in a six-well plate. The human DCs were infected with wBV or rBV at an MOI of 100 for 4 h at 37°C. The cells were then washed and cultured for 20 h at 37°C. For the phenotypic analysis, MDDCs were washed, stained with FITC-conjugated antihuman CD1a and PE-conjugated antihuman CD80, CD86, HLA-ABC, or HLA-DR (eBioscience, San Diego, CA), and analyzed with a FACSCalibur flow cytometer using CellQuest software. The cytokines secreted in the culture supernatants were analyzed using ELISA kits (BD Biosciences).
Western blot analysis
The human DCs were infected with wBV or rBV at an MOI of 100 and cultured for 24 h at 37°C, and in some cases the human DCs were cultured for 72 h. HEK-293T cells were transfected with the baculovirus transfer vector pAcCAG-gag using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's recommendations and used as a control. The HIV virion fraction was prepared from the culture supernatants of HIV-infected human DCs. The supernatants were centrifuged for 20 min at 3,000 rpm to remove cell debris and then for 2 h at 19,000 rpm in a Beckman NVT-100 rotor to pellet the virus particles. The cells and viral pellets were lysed in lysis buffer [1% Nonidet P-40, 50 mM Tris-HCl (pH 7.5), and 1 mM EDTA]. The lysates were then resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and blotted onto a PVDF membrane (Roche). The expression of HIV-1 gag in rBV-DCs was detected using mouse monoclonal anti-gag antibody (AUSTRAL Biologicals, San Romon, CA). For the detection of APOBEC3, rabbit polyclonal anti-A3A (ABGENT, San Diego, CA) and anti-A3G (ProSci Inc., Poway, CA) antibodies and goat polyclonal anti-A3F (Santa Cruz Biotechnology, Dallas, TX) antibody were used at a 1:1,000 dilution as the primary antibody, and antimouse or antirabbit IgG antibody conjugated with horseradish peroxidase (1:5,000; GE Healthcare Bio-Sciences) was used as the secondary antibody. The proteins were visualized using the ECL plus detection system (Amersham Pharmacia Biotech Inc., Piscataway, NJ), and β-actin was used as the lysate control.
Immunofluorescence staining and confocal microscopy
Control human DCs, wBV-human DCs, and rBV-human DCs were washed with phosphate-buffered saline (PBS) and fixed with 2% paraformaldehyde for 5 min. The human DCs were then permeabilized with 0.1% Triton X-100 in PBS for 10 min and blocked with 3% bovine serum albumin (BSA) for 30 min. For HIV-1 gag expression, the cells were incubated for 1 h with mouse monoclonal antigag antibody (AUSTRAL Biologicals), and for A3 expression, the cells were incubated with rabbit polyclonal anti-A3A (ABGENT) or anti-A3G antibody (ProSci Inc.) or with goat polyclonal anti-A3F (Santa Cruz Biotechnology) antibody at 1:500 dilution and then with FITC-conjugated antirabbit IgG or antigoat IgG antibody (Santa Cruz Biotechnology), respectively. The nuclei were stained with DAPI (Sigma Aldrich), and the protein expression was visualized by confocal microscopy using a Zeiss (LSM-510, V-2.5) laser imaging confocal microscope.
Reverse transcription-PCR analysis
The total RNA from control human DCs, wBV-human DCs, or rBV-human DCs was extracted using a GenElute Mammalian Total RNA Miniprep Kit (Sigma Aldrich) according to the manufacturer's instructions. cDNA was synthesized using ReverTra Ace-α (TOYOBO, Osaka, Japan). The PCRs for A3A, A3F, A3G, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were performed by TaKaRa Ex Taq Hot Start Version (TaKaRa, Saga, Japan) using the following primers: forward primers, 5′-TTCTTTGCAGTTGGACCCGG-3′ for A3A, 5′-TACGCAAAGCCTATGGTCGG-3′ for A3F, 5′-TCATCTAGTCCATCCCAGGG-3′ for A3G, and 5′-CCTTGGAGAAGGCGGGG-3′ for GAPDH; and reverse primers, 5′-CTCATCTAGTCCATCCCAGG-3′ for A3A, 5′-GCTCCAAGATGTGTACCAGG-3′ for A3F, 5′-TTACCTGCTTCACCTCCTGG-3′ for A3G, and 5′-CAAAGTTGTCATGGATGACC-3′ for GAPDH. The PCR conditions consisted of an initial denaturation step (94°C for 3 min) followed by 30 cycles of denaturation (94°C for 30 s), annealing (64°C for 30 s), and extension (72°C for 12 s).
HIV-1 infection of human DCs and p24 assay
The human DCs (5×105) were infected with wBV or rBV at an MOI of 100 and cultured for 24 h at 37°C. The control human DCs, wBV-human DCs, and rBV-human DCs were then infected with 100 ng of p24Gag content of HIV-1JR-CSF for 2 h, washed with PBS, and cultured at 37°C. One-half of the culture supernatants was collected for the measurement of p24Gag at the indicated time points and replaced with fresh medium. To monitor the amount of virus, the levels of HIV-1 p24Gag antigen were determined using CLEIA analyzer.
Statistical analysis
All of the data are presented as the mean±SD. The statistical analyses were performed using the Student's t test. p values less than 0.05 were considered statistically significant.
Results
Expression of HIV-1 gag in human DCs infected with AcCAG-gag
We constructed an rBV transfer vector expressing the gag gene of HIV-1 under the control of the CAG promoter (pAcCAG-gag) and produced rBV and AcCAG-gag as described in Materials and Methods. The CAG promoter is used in a wide variety of mammalian cell lines and exhibits stronger protein expression. The human DCs were infected with wBV or rBV at an MOI of 100 and cultured for 4 h at 37°C. At 24 h postinfection, HIV-1 gag expression was detected by Western blot analysis of human DC lysates (Fig. 1A) and by confocal microscopy (Fig. 1B). Three days after the infection of human DCs with rBV, gag expression became undetectable by Western blot analysis. According to this result, we suggest that the infection of human DCs with AcCAG-gag induces the transient expression of gag after 24 h and that this expression fades after approximately 72 h.

Expression of HIV-1 gag protein in recombinant baculovirus AcCAG-gag (rBV)-infected human dendritic cells (DCs). Human DCs were infected with rBV or wild-type AcNPV (wBV) at a multiplicity of infection (MOI) of 100. Uninfected human DCs were used as a control. HIV-1 gag expression was observed after 24 h.
AcNPV and AcCAG-gag induces human DC activation and cytokine production
Previous studies have shown that baculovirus induces the phenotypical and functional maturation of human immature DCs, and the baculovirus-matured DCs displayed a high T cell-stimulatory capacity. 44,45 We also reported that BV induces the functional maturation of human monocyte-derived DCs and the activation of human NK cells via BHDCs. 34 In the present study, we investigated whether the wBV and rBV infection of human DCs can induce their activation. The human DCs were infected with wBV or rBV at an MOI of 100 and cultured for 4 h at 37°C. After 24 h, the human DCs were analyzed for the expression of CD80, CD86, MHC I, and MHC II using a FACSCalibur flow cytometer, and the cytokine levels in the culture supernatant were measured by ELISA. The wBV- and rBV-infected human DCs showed the upregulation of the activation markers CD80 and CD86 as well as the human DC maturation markers MHC I and MHC II compared with the levels observed in the control human DCs (Fig. 2A). The AcCAG-gag- and AcNPV-infected human DCs produced the proinflammatory cytokines IL-12p70, IL-15, and interferons, namely IFN-α and IFN-γ, at greater levels than the control human DCs (Fig. 2B). These results indicate that both wBV and rBV strongly induce the activation and maturation of human DCs and the production of cytokines.

wBV and AcCAG-gag induce the activation of human DCs and the production of proinflammatory cytokines. Human DCs grown in a six-well plate were infected with wBV or rBV at an MOI of 100 for 24 h. Control human DCs were left uninfected.
AcNPV and AcCAG-gag upregulates APOBEC3 expression in human DCs
We then investigated whether infection with AcCAG-gag and AcNPV can upregulate the expression of A3A, A3F, and A3G in human DCs. Human DCs were infected with wBV or rBV at an MOI of 100 for 4 h at 37°C. The cells were then washed and cultured for 20 h at 37°C. The mRNA levels of A3A, A3F, and A3G were assessed by reverse transcription-PCR using respective primers, and the protein expression was examined by Western blot analysis of cell lysates. Both the mRNA (Fig. 3A) and protein expression levels (Fig. 3B) of A3A, A3F, and A3G were increased in the rBV-human DCs and wBV-human DCs compared with those observed in the control human DCs. Furthermore, we analyzed the expression of A3A, A3F, and A3G in rBV-human DCs and wBV-human DCs by immunofluorescence microscopy. As shown in Fig. 3C, the upregulation of A3A, A3F, and A3G was observed in the rBV-human DCs and wBV-human DCs compared with the control human DCs.

wBV and AcCAG-gag upregulate the expression of A3A, A3F, and A3G in human DCs.
Inhibition of HIV-1 replication in AcNPV and AcCAG-gag-infected human DCs
To further explore the potential role of AcNPV and AcCAG-gag in antiviral efficacy, human DCs were infected with 100 ng of p24Gag content of HIV-1JR-CSF and HIV-1gag content in the cell lysates and supernatants for the indicated time points. The level of HIV-1gag in the cell lysate was decreased after 3 days in wBV-human DCs and rBV-human DCs compared with the control human DCs (Figs. 4A and B). The inhibition of HIV-1 replication in wBV-human DCs and rBV-human DCs was further confirmed by CLEIA of the production of p24Gag in the culture supernatants. The result showed an abrogation of progeny virion production in wBV-human DCs and rBV-human DCs at 3 days postinfection. The HIV-1 p24Gag levels in wBV-human DCs and rBV-human DCs were reduced compared with the levels in the control human DCs (Fig. 4C). To assess the kinetics of viral replication, the control human DCs, wBV-human DCs, and rBV-human DCs were infected with HIV-1JR-CSF and cultured for 7 consecutive days. Virus replication was then monitored by CLEIA of the production of p24Gag in the culture supernatant. On day 7, the p24Gag levels in the culture supernatant of wBV-human DCs and rBV-human DCs were reduced in comparison with the levels observed in the control human DCs (Fig. 4D).
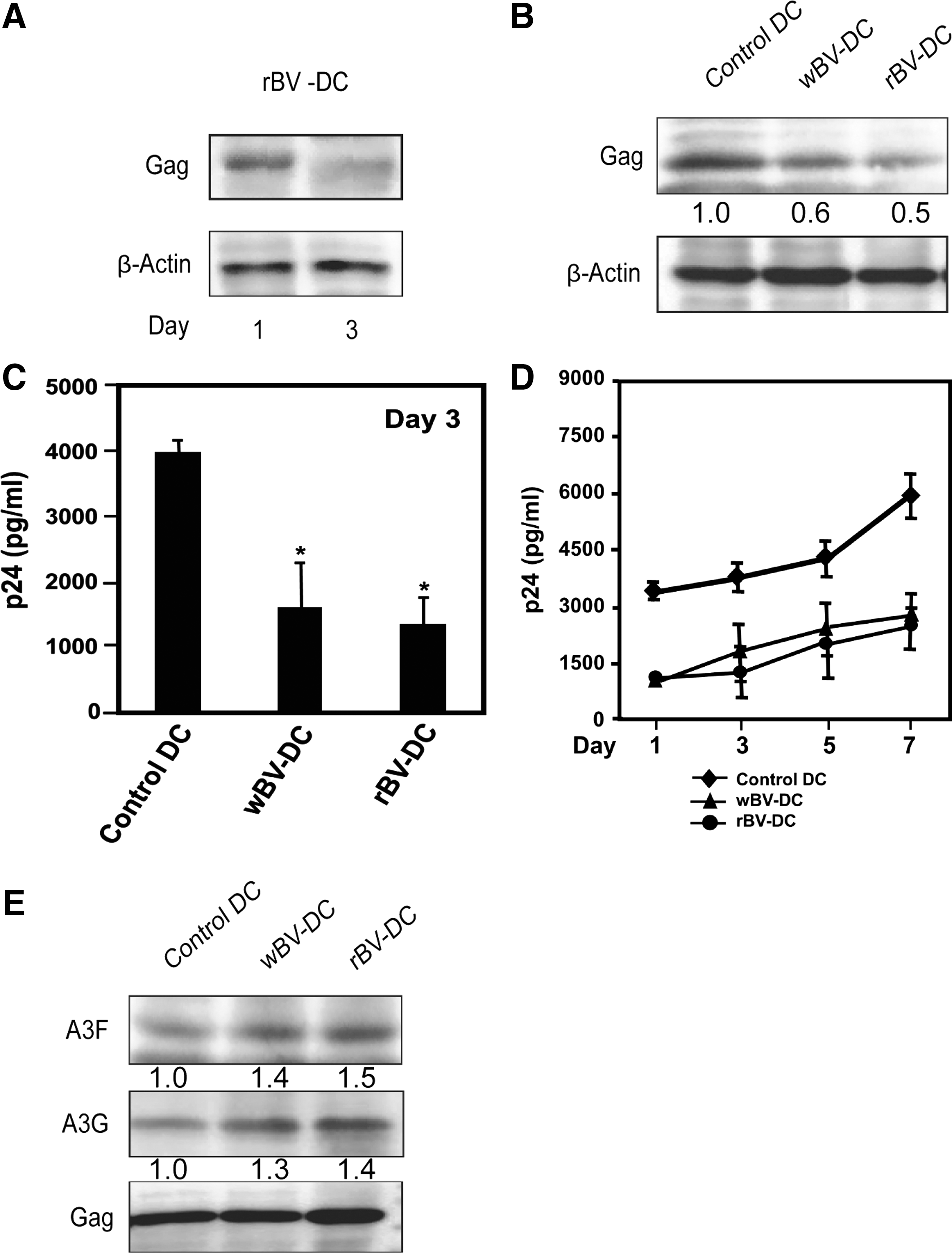
wBV and AcCAG-gag inhibit HIV-1 replication in human DCs and increase APOBEC3 incorporation into nascent virions released from wBV-human DCs and rBV-human DCs.
Increased incorporation of APOBEC3 into nascent virions released from baculovirus-infected human DCs
We next examined the incorporation of A3A, A3F, and A3G into nascent HIV virions released from baculovirus-infected-human DCs by Western blot analysis. The activation of human DCs by wBV or rBV enhances A3F and A3G incorporation into HIV-1 particles compared with the control human DCs. However, the incorporation of A3A in HIV-1 particles could not be detected (Fig. 4E).
In conclusion, our data show that both AcNPV and AcCAG-gag efficiently induce the activation and maturation of human DCs to produce proinflammatory cytokines, including IFN-α, and to upregulate the expression of A3A, A3F, and A3G, leading to the suppression of HIV-1 replication. Notably, wild-type baculovirus-infected human DCs inhibit HIV-1 replication via APOBEC3 expression.
Discussion
DCs play a critical role in the onset of HIV infection, providing one of the primary sites of HIV replication in vivo before the virus is transmitted to CD4+ T cells and disseminated. HIV-infected DCs also act as viral reservoirs for long-term HIV transmission in vivo. 6 –8 To gain an effective control of HIV-1 replication, the initial viral replication in DCs should be contained. Therefore, the identification of strategies to trigger the initiation of immune responses that inhibit DC-endorsed HIV replication represents a potential strategy to limit HIV infection.
In the present study, the immune efficacy of baculovirus on the activation of human DCs and the inhibition of HIV-1 replication was analyzed. We produced AcCAG-gag (rBV) encoding the HIV-1 gag gene under the control of the eukaryotic CAG promoter. rBV induces the phenotypical and functional maturation of human DCs accompanied by the secretion of proinflammatory cytokines, such as IL-12p70, IL-15, IFN-α, and IFN-γ (Fig. 2). Moreover, the wBV (AcNPV) also induces the phenotypical and functional maturation of human DCs accompanied by the secretion of proinflammatory cytokines, such as IL-12p70, IL-15, IFN-α, and IFN-γ (Fig. 2). Although the exact mechanism underlying the baculovirus-mediated activation of human DCs remains unclear, the possible phenomena may involve the association of envelope gp64 with mannose receptor or some Toll-like receptors primarily expressed on human DCs or the signaling events triggered by viral entry.
The results show that AcCAG-gag transiently expresses gag in human DCs (Figs. 1 and 4A) and that rBV upregulates the expression of cellular A3A, A3F, and A3G (Fig. 3) and restricts HIV-1 replication (Fig. 4B–D). We also found that wBV upregulates the expression of cellular A3A, A3F, and A3G (Fig. 3) and restricts HIV-1 replication (Fig. 4B–D). Our results suggest that the inhibition of HIV-1 in wBV-human DCs and rBV-human DCs is related to increases in the expression of cellular APOBEC3 and the incorporation of APOBEC3 into newly formed viral particles. Although the control human DCs are expected to release replication-competent nascent virions into the culture supernatants, the wBV-human DCs and rBV-human DCs released replication-defective APOBEC3-incorporated virions. To prove this finding, we examined the content of APOBEC3 in newly released HIV virions in human DC culture supernatants and found that the virions released from wBV-human DCs and rBV-human DCs contained high contents of A3F and A3G, although the incorporation of A3A could not be detected (Fig. 4E).
We previously showed that HIV-1 gag-virus-like particles induce the activation of human DCs and inhibit HIV-1 replication via IFN-α production and APOBEC3 expression. 46 Buonaguro et al. also reported that HIV-1 Pr55gag virus-like particles (HIV-VLPs), produced in a baculovirus expression system, induced maturation and activation of MDDCs. Furthermore, HIV-VLP-loaded MDDCs are able to induce a primary and secondary response in autologous human CD4+ T cells in an ex vivo system. 47 Several reports have also indicated that inactivated viral particles and purified viral proteins are also capable of inducing interferons. 48,49 In our study, the expression of gag in rBV-DCs is suggested to further enhance the production of cytokines, particularly IFN-α, which then increases the expression of cellular APOBEC3. Facciabene et al. showed that A3G in the cytoplasm interacts efficiently with HIV-1 gag, resulting in the transport of most of the cytoplasmic APOBEC3G to the cell membrane during viral assembly and thereby leading to increased A3G incorporation into budding nascent viral particles. 50
We previously reported that baculovirus stimulates murine DCs and IFN production both in vitro and in vivo. 29 –34 The baculovirus itself can elicit both innate and adaptive immune responses, and these responses may be harnessed therapeutically to induce host immunity against viruses. To date, little information is available regarding the induction of immune responses against HIV-1 in human DCs. In this study, we examined the antiviral effects of baculovirus against HIV-1 in human DCs. The results concerning rBV and wBV show that both can induce the activation and maturation of human DCs to produce proinflammatory cytokines, including IFN-α, and to upregulate the expression of A3A, A3F, and A3G, leading to the suppression of HIV-1 replication. Recently, Abe et al. demonstrated that baculovirus-infected DCs and macrophages produce proinflammatory cytokines and type I IFN via Toll-like receptor 9 (TLR9)-dependent and TLR9-independent pathways. 51 –53 The production of these proinflammatory cytokines, particularly IFN-α, may enhance the expression of cellular APOBEC3 and allow inhibition of HIV-1 replication. More recently, we showed that baculovirus induces the functional maturation of human monocyte-derived DCs and the activation of human NK cells via baculovirus-infected human DCs. 34
The functional studies of NK cells have recently suggested that NK cells can contribute to the control of HIV-1 infection through the recognition of virally infected cells by both activating and inhibitory killer immunoglobulin-like receptors (KIRs). 54 –56 The activation of human NK cells via baculovirus-infected human DCs may mediate antiviral activity in HIV-1-infected humans. In the present study, the immune efficacy of rBV was found to be slightly better than that of wBV, which is likely due to the transient expression of gag in rBV-human DCs. Recent studies in mice have shown that wild-type baculovirus induces a strong innate immune response, whereas recombinant baculovirus is able to elicit strong antigen-specific T cell responses. 29,48 Further experiments are needed to verify a role for the inhibition of HIV-1 replication by BV through small interfering RNA-mediated knockdown of APOBEC3 genes.
This study presents the first evidence supporting the feasibility of using baculovirus to activate human DCs and to induce immunity against HIV-1. This efficacy of baculovirus provides further evidence supporting the use of baculovirus as a vector for immunization against HIV-1. The use of baculovirus in clinical applications in human remains far from resolved, and some obstacles, such as safety concerns and identification of the optimal dose, have to be overcome.
In conclusion, our findings provide the novel observation that AcNPV and AcCAG-gag have the ability to induce the immune activation of human DCs and substantially upregulate APOBEC3 expression to exhibit potent antiviral activity. It is noteworthy that wild-type baculovirus-infected human DCs inhibit HIV-1 replication via APOBEC3 expression. Our present findings underscore the potential for using the baculovirus to combat HIV-1 and highlight the possibility that the baculovirus may be harnessed therapeutically in clinical practice.
Footnotes
Acknowledgments
We thank Drs. Ryuichi Sugiyama and Hitoshi Suzuki for the excellent technical assistance provided in the confocal microscopy studies and Ms. Mari Fujisaki for the helpful work she contributed to this research study. We wish to thank Profs. N. Miyano-Kurosaki and K. Hashimoto for their participation in helpful discussions. This work was supported in part by a Grant-in-Aid for High Technology Research (No. 09309011) from the Ministry of Education, Science, Sports, and Culture of Japan, by a Grant-in-Aid for AIDS research from the Ministry of Health, Labor and Welfare of Japan, by a grant from the Supporting Program for Creating University Ventures from the Japan Science and Technology Agency, Japan, and by a grant from the Research and Development Program for New Bio-industry Initiatives from the Ministry of Agriculture, Forestry and Fisheries of Japan.
Author Disclosure Statement
The authors declare no conflicts of interest.