
Abstract
The development of a highly effective vaccine against the human immunodeficiency virus (HIV) will likely be based on rational vaccine design, since traditional vaccine approaches have failed so far. In recent years, an understanding of what type of immune response is protective against infection and/or disease facilitated vaccine design. T cell-based vaccines against HIV have the goal of limiting both transmission and disease progression by inducing broad and functionally relevant T cell responses. In this context, CD4+ T cells play a direct cytotoxic role and are also important for the generation and maintenance of functional CD8+ T and B cell responses. The use of MHC-binding algorithms has allowed the identification of novel CD4+ T cell epitopes that could be used in vaccine design, the so-called epitope-driven vaccine design. Epitope-based vaccines have the ability to focus the immune response on highly antigenic, conserved epitopes that are fully recognized by the target population. We have recently mapped a set of conserved multiple HLA-DR-binding HIV-1 CD4 epitopes and observed interferon (IFN)-γ-producing CD4+ T cells when we tested these peptides in peripheral blood mononuclear cells (PBMCs) from HIV-infected individuals. We then designed multiepitopic DNA vaccines that induced broad and polyfunctional T cell responses in immunized mice. In this review we will focus on alternative strategies to increase the immunogenicity of an epitope-based vaccine against HIV infection.
Introduction
A
The development of an efficacious vaccine would be facilitated by the knowledge of what type of protective immune response is elicited against infection and/or disease. 5 It is widely accepted that an effective prophylactic vaccine against HIV-1 should elicit neutralizing antibodies (Nabs) that, if present at the time of transmission, could block HIV acquisition. Since virus discovery, six efficacy trials have been conducted but only one, the recent RV144 Thai trial, demonstrated some level of protection. In this Phase III trial, researchers observed protection against HIV-1 acquisition among vaccinees in the absence of serum-neutralizing antibodies, with an estimated vaccine efficacy of 31.2%. 6 Preliminary analysis of the protective immune responses revealed that most vaccinated HIV-negative individuals presented predominantly polyfunctional effector CD4+ T cell responses against the V2 region of the envelope protein. 7 –9 Alternative approaches have focused on vaccines that stimulate cell-mediated immune responses (CMI) against conserved HIV-1 proteins in an attempt to limit viral transmission and prevent HIV-1-associated disease progression by controlling viral loads in those individuals who become infected. 10
Here we will focus on the development of a CD4+ T cell epitope anti-HIV-1 vaccine and on the multiple strategies for increasing its immunogenicity.
Role of CD4+ T Cells in HIV Infection
The role of virus-specific CD4+ T cell responses in protective immunity has been highlighted in recent years. During HIV infection, and also in simian immunodeficiency virus (SIV) infection in nonhuman primates, CD4+ T cells can suppress virus replication directly (through the release of perforin and granzymes) or indirectly (through the release of soluble antiviral chemokines), maintain mucosal immunity by producing interleukin (IL)-17, and produce cytokines to help B and CD8+ T cells. 11 In SIV infection, specific CD4+ T cells can contribute directly to effective suppression of the virus by inhibiting SIV replication in macrophages. 12 Depletion of CD4+ T cells in macaques infected with SIV increased disease progression, suggesting that CD4+ T cells are important for disease control. 13
The protective role of HIV-specific CD4+ T cells was initially suggested in the less virulent HIV-2 infection 14 since the majority of infected individuals with lower viral loads presented gag-specific CD4+ T cell responses.
A significant association was observed between the breadth and specificity of HIV-specific CD4+ T cells and low viral load in HIV-infected individuals. 15 The polyfunctionality of HIV-specific CD4+ T cell responses is greater in nonprogressors than in patients with rapid disease progression. Indeed, HIV-specific CD4+ T cells from elite controller individuals are more polyfunctional and produce more interferon (IFN)-γ/IL-2/tumor necrosis factor (TNF)-α in response to HIV peptides when compared to those from progressors. 16,17 More recently, it was demonstrated that HIV-infected individuals with a lower set point viral load, slower disease progression, and better clinical outcome in the absence of antiretroviral therapy show a significant expansion of HIV-specific CD4+ T cell responses. Moreover, these subjects present enhanced CD4+ T cell cytolytic activity and a higher frequency of IFN-γ-producing cells. 18 Those data suggest that the polyfunctional HIV-specific CD4+ T cell responses may contribute to control of disease progression.
The integrity of the CD4+ T central memory compartment is important to provide the help that promotes the persistence of memory and effector CD8+ T cells. 19 Indeed, HIV infection controllers show a preserved CD4+ T central memory compartment, which has been considered a correlate of protection. 16 IL-21 is a cytokine produced mainly by CD4+ T cells, especially T follicular helper cells (Tfh). 20 Expansion of specific CD4+ T cells producing IL-21 correlated with lower viral set points in HIV controllers compared to progressors, 21 suggesting that IL-21+ CD4+ T cells might contribute to the control of viral replication. Of note, the interaction of CD4 Tfh with B cells in the germinal center is essential for somatic mutation and high-affinity neutralizing antibody production. 20 More recently, it was shown that there is a close association of HIV control and HLA-DRB1 alleles, such as DRB1*13 22,23 and DRB1*15:02. 15
CD4+ T cells may play other beneficial roles in HIV infection. Recently, it was shown that reconstitution of HIV-1-specific CD4+ T cell function followed the administration of therapeutic vaccine, the HIV-1 Gp120/Nef/Tat subunit protein, and increased NK cell activity in HIV-1-infected individuals. 24 This collaborative function of CD4+ T cells to promote innate immune response is also an advantage of a CD4+ T cell-based vaccine.
Since HIV-specific CD4 T cells are the main targets of the virus, 25 a major concern is that a vaccine that induces this type of response might fuel infection rather than help control viral replication. However, to date there is no evidence that vaccine-induced CD4+ responses would hamper the control of infection. On the contrary, the majority of RV144 uninfected vaccine recipients presented specific CD4+ T cell responses. 7,9
It is thus clear that a successful HIV vaccine is likely to also induce CD4+ T cell responses. 26
T Cell-Based Vaccine Design—Epitope Selection
Conventional approaches to vaccine development are based on live-attenuated, inactivated, or isolated components from the pathogen. Those approaches allowed the development of many successful vaccines. 27 However, success was not achieved for many pathogens that lack an immunodominant component or for pathogens that cannot be directly used due to safety reasons, such as HIV. 28
The development of techniques that allowed pathogen whole-genome sequencing and bioinformatics tools drastically changed the rationale for vaccine development since it made possible the identification of potential antigenic targets with protective potential. This new approach in vaccine development is known as reverse vaccinology. 27,29
Given that only the few amino acid residues that comprise an epitope, instead of the whole protein, are sufficient to elicit an immune response, efforts are being made to identify/predict these amino acid sequences. Methods and approaches to identify T cell epitopes are critically important for the development of T cell-based vaccines.
30
T cells recognize antigen via binding of T cell receptors (TCR) to self or foreign peptides.
31
This phenomenon is dependent on the previous binding of the antigenic peptides to cell surface glycoproteins, the major histocompatibility complex (MHC) proteins, expressed by an antigen-presenting cell (APC). The human MHC locus (HLA, Human Leukocyte Antigens) encodes three HLA class I molecules (HLA-A, -B, -C) and three HLA class II molecules (HLA-DR, -DQ, -DP). The structures of MHC class I and class II molecules complexed with different peptides were solved by X-ray crystallography studies. CD8+ T cell epitopes bound to MHC class I range from 8 to 11 residues, while CD4+ T cell epitopes bound to HLA class II range from 11 to more than 20 residues in length. Class I and class II MHC molecules are highly polymorphic. To date, more than 9,000 class I and 3,000 class II alleles have been identified according to the HLA allele website (
A conventional approach to identify T cell epitopes is to synthesize many overlapping peptides (usually 15-mer) spanning the full length of the target antigen and test for its immunogenicity using T cell assays. However, this approach is time consuming, expensive (especially for whole proteome analysis), and may not disclose all the longer CD4+ T cell epitopes. On the other hand, bioinformatics/in silico tools can predict which peptides are more likely to contain T cell epitopes, greatly reducing the number of candidate sequences. 32 Usually these algorithms screen protein sequences for peptides from 8 to 30 amino acids predicted to bind one or more MHC alleles. This method has some advantages since it can select the epitopes able to bind one and/or many MHC molecules. A range of bioinformatics algorithms was developed to predict MHC class II epitopes (Table 1). Initially, the methods for T cell prediction epitopes were based on the identification of anchor motifs of the peptide.
SVM, support vector machine; SVR, support vector regression; ANN, artificial neural network.
References for Table 1
1. Sturniolo T, Bono E, Ding J, et al.: Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat Biotechnol 1999;17(6):555–561.
2. Singh H and Raghava GP: ProPred1: Prediction of promiscuous MHC Class-I binding sites. Bioinformatics 2003;19(8):1009–1014.
3. Nielsen M, Lundegaard C, Blicher T, et al.: Quantitative predictions of peptide binding to any HLA-DR molecule of known sequence: NetMHCIIpan. PLoS Comput Biol 2008;4(7):e1000107.
4. Zhang GL, Khan AM, Srinivasan KN, et al.: MULTIPRED: A computational system for prediction of promiscuous HLA binding peptides. Nucleic Acids Res 2005;33(Web Server issue):W172–179.
5. De Groot AS, Jesdale BM, Szu E, et al.: An interactive Web site providing major histocompatibility ligand predictions: Application to HIV research. AIDS Res Hum Retroviruses 1997;13(7):529–531.
6. Reche PA, Glutting JP, and Reinherz EL.: Prediction of MHC class I binding peptides using profile motifs. Hum Immunol 2002;63(9):701–709.
7. Zhang GL, Srinivasan KN, Veeramani A, et al.: PREDBALB/c: A system for the prediction of peptide binding to H2d molecules, a haplotype of the BALB/c mouse. Nucleic Acids Res 2005;33(Web Server issue):W180–183.
8. Rammensee H, Bachmann J, Emmerich NP, et al.: SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999;50(3–4):213–219.
9. Guan P, Doytchinova IA, Zygouri C, and Flower DR: MHCPred: A server for quantitative prediction of peptide-MHC binding. Nucleic Acids Res 2003;31(13):3621–3624.
10. Donnes P and Elofsson A: Prediction of MHC class I binding peptides, using SVMHC. BMC Bioinformatics 2002;3:25.
11. Lata S, Bhasin M, and Raghava GP: Application of machine learning techniques in predicting MHC binders. Methods Mol Biol 2007;409:201–215.
12. Antonets DV and Maksiutov AZ: [TEpredict: Software for T-cell epitope prediction]. Mol Biol (Mosk) 2010;44(1):130–139.
13. Liu W, Meng X, Xu Q, et al.: Quantitative prediction of mouse class I MHC peptide binding affinity using support vector machine regression (SVR) models. BMC Bioinformatics 2006;7:182.
14. Nielsen M, Lundegaard C, and Lund O: Prediction of MHC class II binding affinity using SMM-align, a novel stabilization matrix alignment method. BMC Bioinformatics 2007;8:238.
Other methods were based in quantitative matrices (QM) considering the score of each amino acid in a certain position. Some methods developed are machine-learning techniques such as artificial neural networks (ANN), virtual matrices, support vector machine (SVM), MHC peptide threading, three-dimensional quantitative structure and activity relations (3DQSAR), and molecular modeling. 33 All those methods are able to select epitopes and have some advantages and disadvantages. Indeed, a comparative study using different MHC class II epitope prediction programs showed that the majority was able to identify the nonamers from a given protein with high sensitivity. 34
Considering the diversity of HLA types observed in the population, others and ourselves 35 –38 believe that an effective epitope-based vaccine should contain promiscuous epitopes that are able to bind to several HLA class II molecules. To select the epitopes for our anti-HIV vaccine, we used the algorithm TEPITOPE that can predict those epitopes that have the potential ability to bind to one or more of 25 different HLA-DR molecules by using 25 virtual matrices that cover most of the HLA class II peptide binding specificities in the white population. 39,40 TEPITOPE also allows detection of sequences predicted to bind to several HLA-DR molecules simultaneously, making it possible to select promiscuous T cell epitopes. 41 This approach has been successfully used by our group to identify allele-specific and promiscuous T cell epitopes in several pathogen antigens and autoimmune disease targets 37,42 –44 as well as by others. 41,45,46
Using the TEPITOPE algorithm to scan the whole proteome of the HIV-1 B-subtype consensus sequence, we identified 18 sequences (Table 2) predicted to bind significantly to at least two-thirds of the HLA-DR molecules covered by the algorithm representing about 90% of the white population. 35 Binding assays with the nine most prevalent HLA-DR molecules in the general population confirmed the ability of the selected peptides to bind to multiple HLA-DR molecules.
Peripheral blood mononuclear cells from over 90% of HIV-1-infected patients recognized at least one of the promiscuous peptides and most patients recognized multiple peptides, demonstrating that epitopes are presented in vivo during infection. These results encouraged us to construct multiepitopic vaccines encoding the 18 promiscuous CD4+ T cell epitopes (HIVBr18) and test its immunogenicity in mice transgenic to human HLA class II molecules and BALB/c mice. 36,37,47
DNA Vaccines
Despite a large number of clinical studies (107 ongoing studies in the world as of March 2015, see
Use of Adjuvants
Currently, adjuvant selection brings a major breakthrough for the use of DNA vaccines for both prophylactic and therapeutic treatments. Different categories of adjuvants were studied based on their nature, but few have advanced for use in humans, in part because of the difficulty to license them according to the Guideline on Adjuvants in Vaccines for Human Use developed by The European Medicines Agency or by the Federal Drug Agency (FDA).
Alum particles 50 and MF59 emulsions 51 when mixed with plasmids were shown to improve the immunogenicity of DNA vaccines. Toll-like receptor (TLR) agonists have been used as adjuvants with DNA vaccines against SIV. 52,53 Adjuvanting a DNA vaccine with the TLR9 agonist, CpG oligodeoxynucleotides (ODN), at the time of priming enhanced CD8+ T cell responses and control of viral load after SIV challenge in rhesus macaques. 53 However, when an SIV DNA vaccine was administered together with the TLR3 agonist poly(ICLC), in rhesus macaques, no improvement in immunogenicity was observed. 54
Nonclassical adjuvants, also called “genetic adjuvants,” are plasmids that after injection drive the production of cytokines, chemokines, and costimulatory molecules. They can be incorporated in the same (bicistronic system) or in another plasmid (admixed formulation). 55 To increase the immunogenicity of DNA immunization using HIV and/or SIV antigens, genes encoding several cytokines 56 –59 were used.
Granulocyte-macrophage colony-stimulating factor (GM-CSF) associated with DNA vaccines recruits dendritic cells to the site of vaccination and increases antigen presentation. 60,61 Also, it is able to potentiate specific T cells, driving the development of Th1 immune responses. 62 This property to potentiate vaccine-induced immune responses was demonstrated in studies with different animal models, such as mice, 63,64 nonhuman primates, 65,66 and also in human clinical trials for antitumor and antiviral vaccines. 67,68
Our group evaluated the combination of pGM-CSF (GM-CSF encoding plasmid) with the HIVBr18 DNA vaccine in mice. Coadministration of 100 μg of each plasmid by the intramuscular route resulted in enhancement of the magnitude of the cellular immune response against pooled HIV-1 peptides (data not shown). Moreover, we observed a higher number of polyfunctional T cells (CD3+CD4+IFN-γ+IL-2+TNF-α+) in splenocytes of mice immunized with both plasmids when compared to mice immunized with HIVBr18 alone (Santana et al., unpublished). For HIV, progress has been also made using interleukin-2. In mice, anti-gp120 specific immune response was enhanced when a plasmid encoding the gp120 gene was administered first and then followed by another plasmid encoding a fusion IL-2/Ig (IL-2 fused to the Fc portion of immunoglobulin G; IgG). 69 This study was extended to monkeys using plasmids encoding SIV Gag and HIV-1 Env genes administered with the IL-2/Ig plasmid. No evidence of clinical disease was observed in the group of monkeys that received the combination of plasmids and were subsequently challenged. 70 A clinical trial was then set in place to evaluate an HIV-1 DNA vaccine with the plasmid cytokine adjuvant (IL-2/Ig) in 70 HIV-negative adults. As observed in the mouse study, IL-2/Ig plasmid increased immune responses when administered 2 days after the DNA vaccine. 71
Cytokines such as IL-12 and IL-15 have been effective in enhancing immune responses against HIV/SIV in both murine and nonhuman primate models. 58,59 The coadministration of plasmids encoding IL-12 and IL-15 in macaques increased the specific CD8+ T cell memory populations and their ability to produce cytokines. 72 In humans, three clinical trials were completed using HIV DNA vaccines administered together with plasmids encoding IL-12 (NCT00991354), or IL-12 and IL-15 (NCT00528489 and NCT00775424).
The envelope glycoprotein D (gD) of the human herpes simplex virus (HSV-1) can be used as a molecular adjuvant for DNA vaccines. 73 The immunological effects of gD involve the binding of gD to herpes virus entry mediator (HVEM) and the blockage of a coinhibitory immune mechanism involving the B- and T-lymphocyte attenuator (BTLA) cell receptor. 74,75 Blockade of the HVEM-BTLA pathway during vaccine-induced immune responses was associated with increased immune responses, particularly in antiviral and anticancer vaccines. 75 –77 We designed a bicistronic plasmid driving the expression of HSV gD and HIVBr18 to immunize BALB/c mice. In preliminary ELISpot assays, we observed that mice immunized with the bicistronic plasmid presented a similar T cell immune response against pooled HIV-1 peptides when compared to mice immunized with HIVBr18 plasmid alone (data not shown). These data suggest that this mechanism may not be useful for all antigens, given the complexity of costimulatory/coinhibitory molecules during antigen presentation and T cell–APC interaction.
Chemical compounds were also evaluated as adjuvants for DNA vaccines. 78 One of these compounds is bupivacaine, a local anesthetic drug that belongs to the amino amide group that blocks neuron transmission. Bupivacaine is a myotoxin that when injected destroys myofiber cells leading to the clearance of cell debris and proliferation of myoblasts. 79 In addition, the recruitment of inflammatory cells to the site of bupivacaine injection may allow the transfection of immune cells. 80 Pretreatment of muscle with 0.25–0.5% bupivacaine prior to DNA injection increases gene expression by 80-fold, resulting in an enhancement of the immune responses. 79 Also, the complex formed with bupivacaine protects DNA from nuclease degradation, and intramuscular immunization with this formulation results in higher immune responses against the encoded antigen. 81
Recently, we evaluated the concomitant use of bupivacaine to improve the immunopotency of the DNA vaccine HIVBr18. 82 We observed that administration of a DNA vaccine encoding HIV CD4+ T cell epitopes together with bupivacaine at the time of immunization increases the overall magnitude of the specific immune response (Fig. 1). In addition, 90 days after the last dose, higher numbers of IFN-γ secreting cells and T cell proliferation were observed in splenocytes from mice immunized with HIVBr18 plus bupivacaine when compared to mice immunized with HIVBr18 only (data not shown). Such an ability to promote long-term immunological memory is a critical feature of an effective vaccine.

Coadministration of HIVBr18 and bupivacaine enhances specific T cell responses. Proliferative T cell responses were assessed by carboxyfluorescein N-succinimidyl ester (CFSE) dilution assay. Splenocytes from immunized mice were labeled with CFSE and cultured in the presence of pooled HIV-1 peptides or medium only. CFSE dilution on gated CD3+CD4+
Use of Viral Vectors
A better understanding of viral biology in recent years facilitated the use of viruses as vaccine vectors. A wide range of viruses (poxvirus, adenovirus, adeno-associated virus, yellow fever virus, cytomegalovirus, etc.) has been extensively studied for their ability to express exogenous antigens and induce specific immune responses. 83,84
Adenovirus (Ad) is one of the most exploited since its safety is well-documented. 85 Adenoviruses display several features such as high levels of transgene expression, no integration in the host genome, physical and genetic stability, the ability to infect dendritic cells, and enhanced expression of costimulatory molecules that culminate in greater cytokine/chemokine production.
Several clinical trials showed that vaccines based on replication-defective adenovirus serotype 5 (Ad5) are safe and highly immunogenic, 86,87 even though the Ad5-vectored trivalent HIV vaccine tested in the STEP trial showed no protection. 88 Further analysis of the STEP trial indicated that male volunteers who received the vaccine candidate had a statistically significant elevated risk of acquiring HIV if they were uncircumcised and had preexisting immunity to the Ad5 vector. Another trial, HVTN 505, was originally designed to enroll circumcised, HIV-uninfected individuals with no preexisting immunity to adenovirus serotype 5 (Ad5). The investigational HIV vaccine regimen involved a series of three immunizations, beginning with a DNA-based vaccine, followed by an Ad5 vector-based vaccine candidate. The evolution of the HVTN 505 trial was driven by the outcomes of two other efficacy trials: the STEP trial and the RV144 trial that tested a canarypox vector-based candidate in combination with a protein boost. The latter study energized the AIDS vaccine field 2 years later, when it yielded the first evidence of vaccine-induced protection against HIV. But just like the vaccine candidate tested in the STEP trial, the regimen used in the HVTN 505 trial reduced neither the rate of HIV-1 acquisition nor the viral load set point. 40
One challenge with adenovirus vectors, especially Ad5, is the high prevalence of preexisting immunity against the vector in some areas, since this is the commonly circulating strain of the cold virus. 89 This phenomenon is being extensively examined and this obstacle can be overcome by the development of novel vectors based on less prevalent Ad serotypes (Ad26, Ad35) or nonhuman primate adenovirus (Ad68). 90 –92
To test the immunogenicity of an adenovirus expressing our multiepitopic gene, we generated an Ad5HIVBr18 and compared the induced cellular immune response with the DNA counterpart (HIVBr18). We observed that immunization with Ad5HIVBr18 induced stronger T cell responses than the DNA vaccine HIVBr18 without impairing the breadth of the immune responses. Ad5HIVBr18-immunized mice displayed significantly higher proliferative (Fig. 2), as well as polyfunctional CD4+ and CD8+ T cell responses when compared to HIVBr18. Most of these responses were highest among animals immunized with Ad5HIVBr18 by the subcutaneous route. 47 The same immunization protocol induced the highest T cell immune responses even at 28 weeks after the last immunization, indicating that it was also able to improve the longevity of the vaccine-induced immune responses. An additional advantage of immunization with Ad5HIVBr18 as compared to HIVBr18 was the reduced number of required doses (two vs. three doses, respectively).
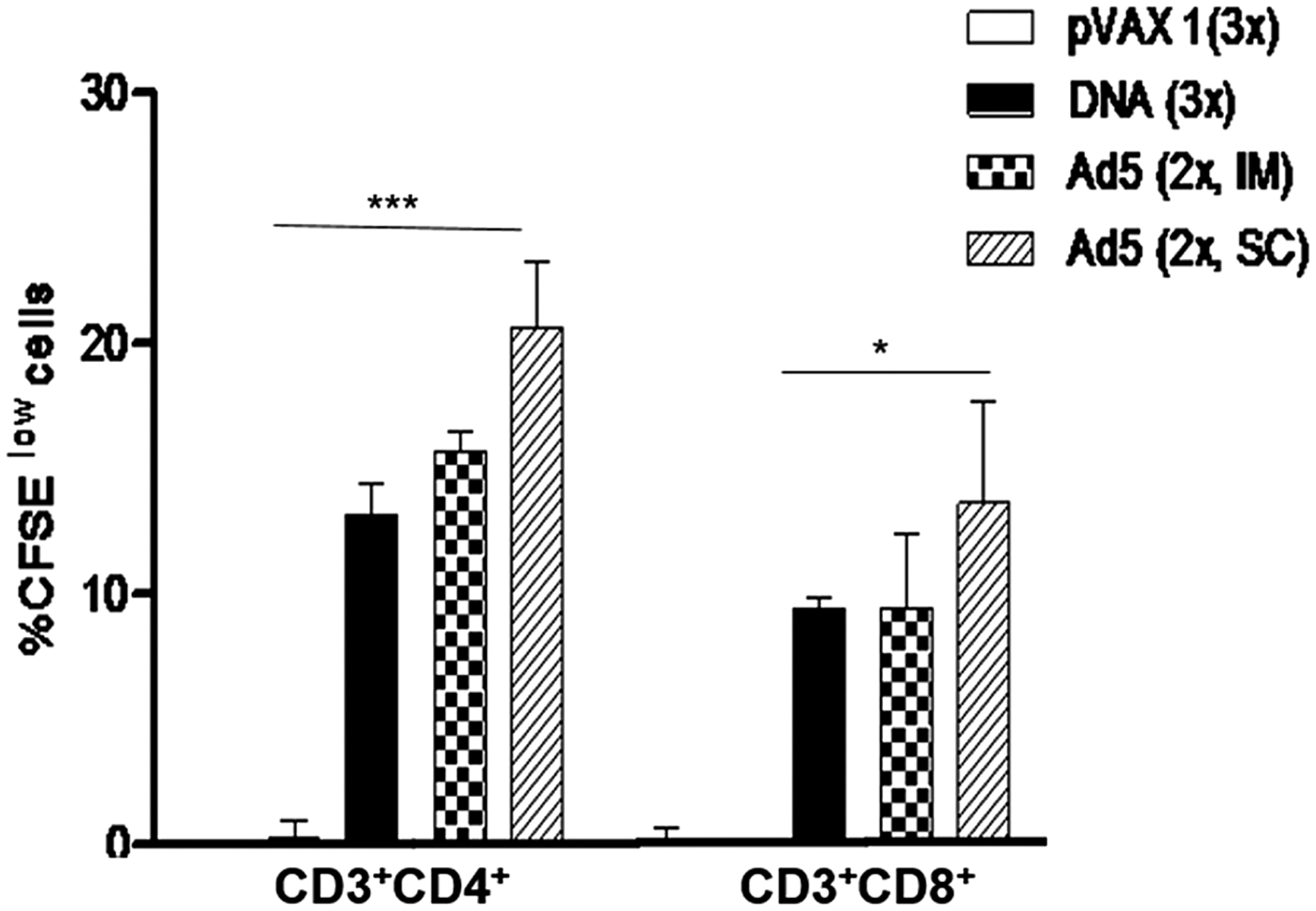
Immunization with an adenovirus encoding the multiepitope gene (Ad5-HIVBr18) elicits higher proliferative responses when compared to other regimens. BALB/c mice were immunized intramuscularly (IM) with the DNA vaccine HIVBr18 (three doses), or recombinant adenovirus (Ad5) by the intramuscular (IM) or subcutaneous (SC) route, or received a heterologous prime-boost regimen based on a DNA prime (IM), Ad5 boost (IM or SC). Control groups received pVAX1 plasmid. Two weeks after the last immunization splenocytes from each immunized group were labeled with CFSE and cultured in the presence of pooled HIV-1 peptides. Cells were analyzed by flow cytometry and CFSE dilution on gated CD3+CD4+ or CD3+CD8+ cells was used as readout for antigen-specific proliferation. Only significant differences with the DNA immunized group are depicted (*p<0.05 and ***p<0.001).
Another promising viral vector for vaccine purposes is cytomegalovirus (CMV), which has been used as a persistent viral vector to deliver SIV genes. Immunization with a rhesus CMV proved to be highly successful to induce virologic control against a highly pathogenic mucosal SIV challenge. 93,94
Adeno-associated virus (AAV) has being widely used for gene therapy applications against disease such as cystic fibrosis, muscular dystrophy, and hemophilia B.
95
More recently, recombinant AAV (r-AAV) has emerged as a vector to delivery HIV genes.
96
–99
Antibody gene transfer mediated by rAAV, also known as vectored immunoprophylaxis (VIP), has been successfully used to produce neutralizing antibodies in mice,
100
monkeys,
101
and also in humanized mice. Such antibodies were able to protect against intravenous as well as mucosal HIV transmission.
102
A Phase I clinical trial of an AAV vector encoding the HIV neutralizing antibody PG9 started and according to clinicaltrials.gov is still recruiting (
Increasing the Number of Epitopes to Expand the Breadth of T Cell Immune Responses
The failure of multiple HIV-1 vaccines has been partly attributed to either the low breadth, as observed in the STEP trial, 103 of induced T cell immune responses or to the inability to elicit cross-clade immunity. 93 Strategies to cope with viral genetic diversity, due to high rates of mutation, and the diversity of viral strains, is considered a potential alternative to induce broad T cell responses. The use of artificially designed M-group consensus HIV-1 sequences and mosaic proteins has been used to achieve this goal. 104 In fact, it was proposed that a vaccine composed of viral segments conserved across all M-group subtypes would limit HIV-1 replication by focusing the CD8+ T cell immune response on regions that are crucial for viral viability. 105 In the same context, other studies showed that immunization of either mice or macaques with M-group consensus envelope elicits broader cross-clade neutralizing antibodies and T cell responses when compared to immunogens based on single HIV-1 subtypes. 33,106,107
To increase the immunogenicity of our vaccine approach and ensure that it would provide CD4+ T cell immune responses toward the global HIV-1 diversity, we developed a DNA vaccine (HIVBr27) encoding an increased number of viral peptides, highly promiscuous and conserved among all M-group subtypes. 108 Immunization of BALB/c mice elicited IFN-γ secretion against 11 out of the 27 vaccine-encoded peptides (Fig. 3). We observed CD4+ and CD8+ T cell proliferation against eight and six peptides, respectively (Fig. 4A and B) and cross-clade T cell responses against several HIV-1 peptide variants (Fig. 5A–C). Overall, our data demonstrated that a DNA vaccine encoding an increased number of peptides derived from the HIV-1 M-group consensus sequence was able to improve the immunogenicity of our approach and provide cross-clade immunity.

Immunization with HIVBr27 elicits broad T cell responses in BALB/c mice. BALB/c mice were immunized intramuscularly three times with 100 μg of HIVBr27 or with the empty vector pVAX. Two weeks after the last immunization, pooled spleen cells from six mice per group were cultured in the presence of individual HIV-1 peptides (5 μM) and the frequency of specific IFN-γ secreting T cells was evaluated. The dotted line represents cutoff value. Data are shown as mean±SD of three independent experiments for ELISpot assays.

Immunization with HIVBr27 elicits CD4+ and CD8+ T cell proliferation against multiple HIV-1 peptides. BALB/c mice were immunized intramuscularly three times with 100 μg of HIVBr27 or with the empty vector pVAX. Two weeks after the last immunization, pooled spleen cells from six mice per group were labeled with CFSE (1.25 μM) and cultured for 5 days in the presence of individual HIV-1 peptides (5 μM). After staining with fluorochrome-labeled anti-CD3, -CD4, and -CD8 monoclonal antibodies, cells were analyzed by flow cytometry. CFSE dilution on gated CD3+CD4+

HIVBr27 immunization provides cross-clade immunity. Two weeks after the last immunization with HIVBr27 or empty pVAX1, pooled spleen cells from six BALB/c mice were cultured in the presence of HIV-1 M-group consensus peptides (5 μM) (white bars) or their variants (colored bars) from diverse HIV-1 subtypes. The frequency of IFN-γ-secreting cells was assessed by ELISpot assay
Concluding Remarks
The development of an efficacious anti-HIV vaccine remains a scientific hurdle. Several lines of evidence suggest that a T cell-based vaccine can limit both transmission and disease progression by inducing broad and functionally relevant T cell responses. While the potential role of CD4+ responses in HIV/AIDS vaccines has been acknowledged recently, the mapping of CD4+ T cell epitopes as vaccine candidates has not been extensively explored. In view of the results from HIV vaccine clinical trials that employed whole protein as an immunization concept, we believe that in silico identification of CD4+ T cell epitopes is an alternative for rational vaccine design.
Our group identified a set of conserved and promiscuous CD4+ T cell epitopes from HIV and evaluated its immunogenicity in mice. Since a caveat that can hamper the advance of a vaccine candidate to a clinical trial is the induction of potent immunity in humans, we tested several strategies to increase the immunogenicity of an epitope-based vaccine against HIV infection. We observed that the use of chemical compounds, genetic adjuvants, and viral vectors, and increasing the number of epitopes influence specific immune responses. In addition, this multiepitope-based vaccine concept can be used as a stand alone or as a source of cognate help for whole protein immunization (unpublished observations).
Footnotes
Acknowledgments
This work was supported by the São Paulo State Research Funding Agency (FAPESP) and Brazilian National Research Council (CNPq).
Author Disclosure Statement
No competing financial interests exist.