
Abstract
HIV-1 infection results in immunological abnormalities of natural killer (NK) cells such as disturbed distribution of NK cell subsets and downmodulation of activating and upregulation of inhibitory receptors thereby diminishing NK cell killing capacity and cytokine secretion. Antiretroviral treatment (ART) is known to restore phenotype and functions of NK cells. However, the effects of ART on NK cell terminal differentiation, activation, and disturbed distribution have not been studied yet longitudinally. Here, we analyzed the effects of ART on these parameters of peripheral blood NK cells in a longitudinal as well as in a cross-sectional study. We observed that expanded CD56−CD16+ NK cell frequency is inversely correlated with the frequency of CD56dimCD16+ NK cells in treatment-naive HIV-1 patients. Loss of CD56dimCD16+ and expansion of CD56−CD16+ NK cells again restore to the levels of healthy controls after ART. Enhanced immune activation of different NK cell subsets is partially restored after ART. Terminal differentiation of CD56dimCD16+ NK cells is enhanced after ART as measured by CD57 expression. Frequencies of CD57+CD56dimCD16+ NK cells are directly correlated with the frequencies of total NK cells suggesting that an increase in the frequencies of CD57+CD56dimCD16+ NK cells is reflected by increased frequencies of total NK cells after ART. Taken together these data demonstrate that ART has an effect on the immune restoration of NK cells and is enhanced in the terminal differentiation of CD56dimCD16+ NK cells, which is associated with increased frequencies of total NK cells after ART.
Introduction
N
In progressive HIV-1 infection, the innate and adaptive immune systems are activated due to viral proteins and microbial product translocation from the gut-associated lymphoid tissue (GALT) into the circulation. 11 The activation leads to production of proinflammatory cytokines, increased cell turnover, activation-induced cell death, and cell exhaustion. 11,12 This persistent immune activation plays a central role in driving HIV immunopathogenesis and progression toward AIDS. 13
HIV infection has deleterious effects on phenotype and functions of NK cells. 1,14 Progressive HIV-1 infection is associated with decline in cytotoxic CD56dimCD16+ NK cells with an expansion of the dysfunctional CD56−CD16+ NK cell subset. 15,16 Our group has shown an association between chronic HIV infection and a significant loss of CCR7 on CD56brightCD16+– NK cells and a significant decline of CD56dimCD16+ NK cells, which are either CD57− or CD57dim. 17,18 In addition, we and others demonstrated an expansion of terminally differentiated CD56−CD16+ NK cells in HIV-infected patients, 19,20 a possible effect of an increased turnover of NK cells induced by HIV viremia. 21 Notably, frequencies of highly functional CD8+ NK cells correlate with HIV-1 disease progression. 22 Previously it was considered that absolute counts of NK cells decline during HIV-1 infection. 23,24 However, recent discoveries made over the past years elucidated that HIV-1 viremia is related to a pathological reallocation rather than absolute loss in counts of peripheral blood NK cells. 20
Here we demonstrate in a longitudinal and cross-sectional study that disturbed NK cell subset distributions restore to the normal level as immune activation of NK cells decreases after 1 year of antiretroviral treatment (ART). However, the level of immune activation does not directly correlate with the expansion or restoration of the CD56−CD16+ NK cell subset. CD56dimCD16+ and CD56−CD16+ NK cells show elevated terminal differentiation reflected in the increase of NK cell frequencies after ART as measured by CD57 expression. These data reveal that immune activation of NK cells normalizes while terminal differentiation increases after successful reduction of viremia during ART.
Materials and Methods
Study subjects
A cohort of 21 HIV-1-infected treatment-naive subjects with CD4+ helper T cell counts below 500/μl were followed longitudinally at our institution. From these patients we obtained peripheral blood samples before and after 1 year of ART. For the cross-sectional study we obtained peripheral blood from 35 treatment-naive and 25 treated HIV-seropositive subjects on ART at least for 1 year and 15 healthy individuals in the HIV outpatient clinic at Medizinische Hochschule Hannover (MHH). A summary of the clinical data of the studied groups is shown in Table 1. All study subjects gave written, informed consent prior to their participation. The study was approved by the local ethics committee (Votum der Ethikkommission der MHH No. 3150).
All values are provided as medians and the interquartile ranges are indicated in parentheses.
NK, natural killer.
Isolation of mononuclear cells and plasma HIV-1 RNA analysis
Peripheral blood mononuclear cells (PBMCs) were isolated from fresh blood as described previously. 18,22 Aliquots of 107 PBMCs each were cryopreserved in heat-inactivated fetal calf serum (FCS) supplemented with 10% dimethyl sulfoxide (DMSO) (Merck). Plasma HIV-1 RNA levels were determined using the VERSANT-HIV-1 RNA Assay, version 3.0 (bDNA, Bayer Diagnostics, Berkeley, CA).
Phenotypic analysis of NK cells by flow cytometry
Monoclonal antibodies used in this study were anti-CD19 PerCP, anti-CD14 PerCP, anti-CD16 APC-H7, anti-CD57 FITC (BD Biosciences), anti-CD69 PE (Invitrogen), anti-CD56 PE-Cy7, and anti-CD3 ECD from Beckman Coulter. Via Probe PerCP (BD Biosciences) was used as a viability probe to exclude the dead cells. Staining and flow cytometric analyses were performed as described before. 22,25,26
Statistical analysis
GraphPad Prism (version 5.0) software was used for statistical evaluation of data. The paired t-test was used to compare patient's data before and after ART. Pearson's analysis was performed to find the correlation between NK cell subsets. For the cross-sectional study, one-way analysis of variance (ANOVA) followed by Tukey test was used. p values of less than 0.05 were considered significant.
Results
Disturbed NK cell subsets are restored after ART
We analyzed different subsets of peripheral blood NK cells in a cohort of HIV-1-seropositive patients, longitudinally as well as in a cross-sectional study. To define different subsets of NK cells, we excluded T cells, B cells, monocytes, and dead cells from the analysis and identified CD56brightCD16+/–, CD56dimCD16+, and CD56−CD16+ NK cells (Fig. 1A). During HIV-1 infection NK cell subset distributions are disturbed.
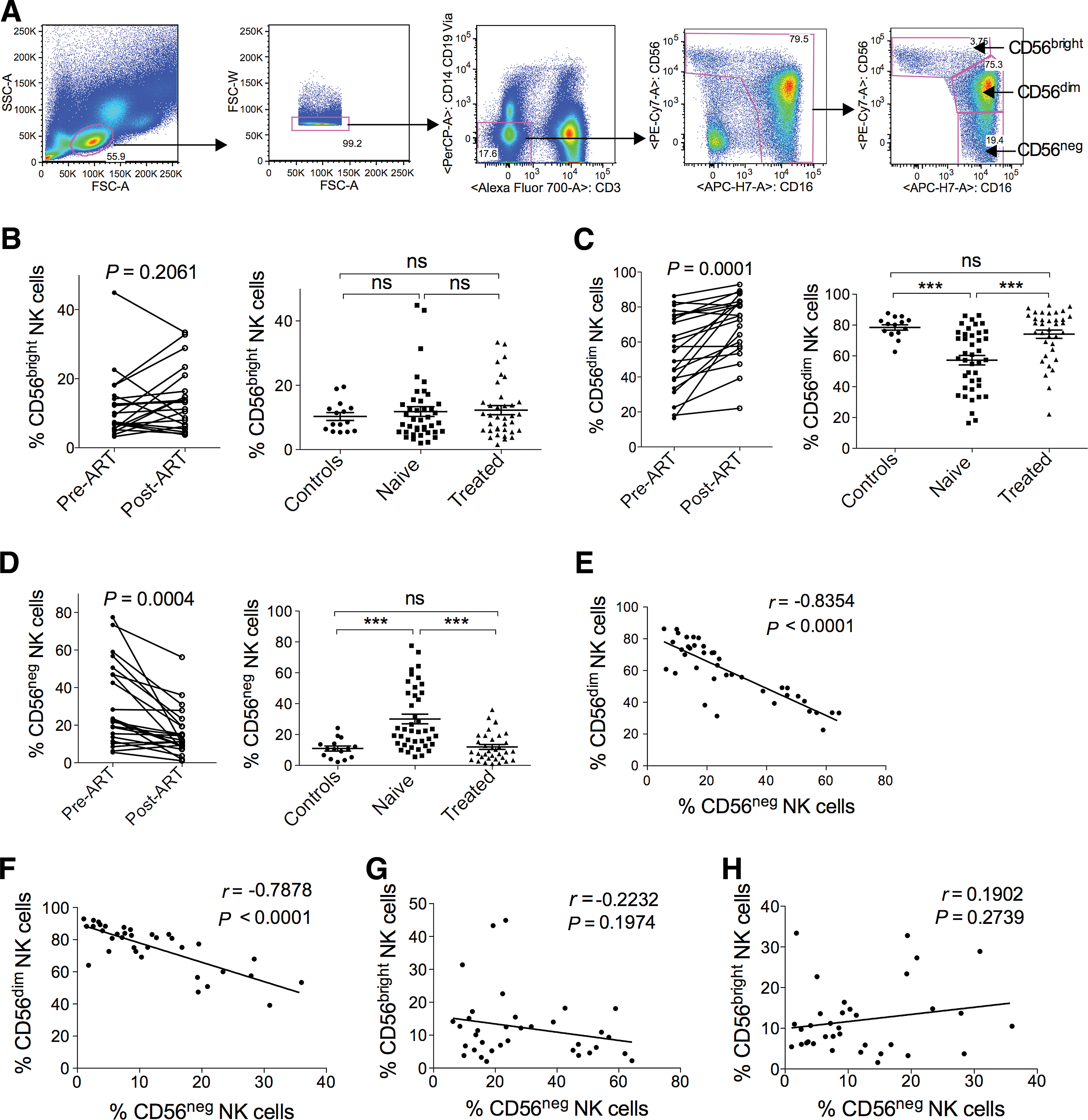
Longitudinal and cross-sectional analysis of different subsets of natural killer (NK) cells.
We wanted to determine whether ART has an effect on the altered distribution of different peripheral blood NK cell subsets. Therefore, we first examined the frequency of CD56brightCD16+/– NK cells, which exert a more immune regulatory function. They remain stable before and after 1 year of ART in the longitudinal study as well as in the cross-sectional study (Fig. 1B). The frequency of cytotoxic CD56dimCD16+ NK cells is reduced (Fig. 1C) while CD56−CD16+ NK cells expand in HIV-1 treatment-naive patients (Fig. 1D). After 1 year of ART, CD56dimCD16+ as well as CD56−CD16+ NK cells are almost restored to the levels of healthy controls (Fig. 1C and D). This outcome is also supported by our longitudinal study (Fig. 1C and D), which corroborates the finding by Brunetta et al.
27
In addition, we also analyzed the absolute numbers of CD56brightCD16+/–, CD56dimCD16+, and CD56−CD16+ NK cells in HIV patients before and after 1 year of ART. No significant alterations were found in terms of the absolute numbers (Supplementary Fig. S1A and B; Supplementary Data are available online at
The finding that CD56dimCD16+ NK cells decrease while CD56−CD16+ NK cells expand makes it necessary to rule out whether the expanded CD56−CD16+ NK cells correlate with the CD56dimCD16+ NK cells in HIV-1 treatment-naive patients and in ART patients. To this point, we performed Pearson's correlation analysis between the frequencies/absolute numbers of CD56dimCD16+ and CD56−CD16+ NK cells in HIV-1 treatment-naive patients as well as in ART patients. We found a strong negative correlation between the frequencies of CD56dimCD16+ and CD56−CD16+ NK cells in both HIV-1 treatment-naive and in ART patients (Fig. 1E and F). The absolute cell counts of the two subsets also showed the trend of inverse correlation but did not reach the level of significance in both HIV-1 treatments-naive and in ART patients (Supplementary Fig. S1D and E).
In addition, we also performed Pearson's correlation analysis between the frequencies/absolute numbers of CD56brightCD16+/– and CD56−CD16+ NK cells in HIV-1 treatment-naive patients as well as in ART patients. Notably, we did not find any significant correlation between the frequencies/absolute numbers of CD56brightCD16+/– and CD56−CD16+ NK cells in both HIV-1 treatment-naive and in ART patients (Fig. 1G and H and Supplementary Fig. S1F and G), suggesting redistribution of CD56dimCD16+ and CD56−CD16+ NK cells within the NK cell compartment during HIV-1 infection. This led us to further analyze CD56dimCD16+ and CD56−CD16+ NK cells with respect to HIV treatment.
Immune activation of NK cells after ART
Chronic immune activation is the hallmark of HIV-1 infection leading to premature aging and apoptosis of immune cells. 28 Therefore we analyzed the early activation marker CD69 on different NK cell subsets cross-sectionally as well as longitudinally (Fig. 2A and C). We found that CD56dimCD16+ and CD56−CD16+ NK cells from treatment-naive HIV-1 patients exhibit higher immune activation compared to healthy controls (Fig. 2B). After 1 year of ART with suppressed viral load there was a substantial decrease in activation of the CD56dimCD16+ NK cell subset, however not to the levels of healthy controls (Fig. 2B). In contrast, immune activation of the CD56−CD16+ NK cell subset decreased to the level of healthy controls in the cross-sectional study. This observation was also supported by our pretreatment and posttreatment analysis of CD69 on CD56dimCD16+ and CD56−CD16+ NK cells (Fig. 2B and D). Similar to CD56dimCD16+ and CD56−CD16+ NK cell subsets, CD56brightCD16+/– NK cells also show a higher activation state in treatment-naive patients compared to healthy controls, which is again restored after ART (data not shown).

Activation of NK cell subsets before and after treatment.
Terminal differentiation of peripheral blood NK cells after ART
For defining terminal differentiation we measured CD57, which has been reported to be a marker for exhaustion and senescence of CD8+ T and NK cells. 17,29 Recently, CD57 has been used to define terminal NK cell differentiation. 30 In contrast to CD56dimCD16+ and CD56−CD16+ NK cells, CD56brightCD16+/– NK cells do not express CD57. 17 Increased proportions of CD57+CD8+ T cells have been observed in some infections such as human parvovirus, measles, and pulmonary tuberculosis. 31 –33 Increased frequencies of CD57+CD16+ cells were first reported in HCMV-infected individuals. 34
Here we examined the terminal differentiation of different NK cell subsets before and after 1 year of ART in a cross-sectional and in the longitudinal study. Representative flow cytometry plots display the frequency of CD57 on CD56dimCD16+ NK cells (Fig. 3A) as well as on CD56−CD16+ NK cells (Fig. 3C). The frequencies of CD57 expressing CD56dimCD16+ NK cells remained similar in treatment-naive HIV-1 patients compared to healthy controls (Fig. 3B). In contrast, CD56−CD16+ NK cells from treatment-naive HIV patients showed an increased frequency of CD57-expressing cells compared to healthy controls in the cross-sectional study (Fig. 3D). After 1 year of ART both CD56dimCD16+ and CD56−CD16+ NK cells revealed an elevated CD57 frequency, although on CD56−CD16+ NK cells the CD57 frequency did not reach the level of significance in the longitudinal study (Fig. 3B and D). When we analyzed the frequencies of CD57+CD56dimCD16+ and CD57+CD56negCD16+ NK cells before and after 1 year of ART, we observed that some of the patients presented increased frequencies of CD57+CD56dimCD16+ cells while others had decreased frequencies of CD57+CD56dimCD16+ NK cells (Fig. 3B and D). These led to a further study of CD57 expression in individual patients before and after treatment.
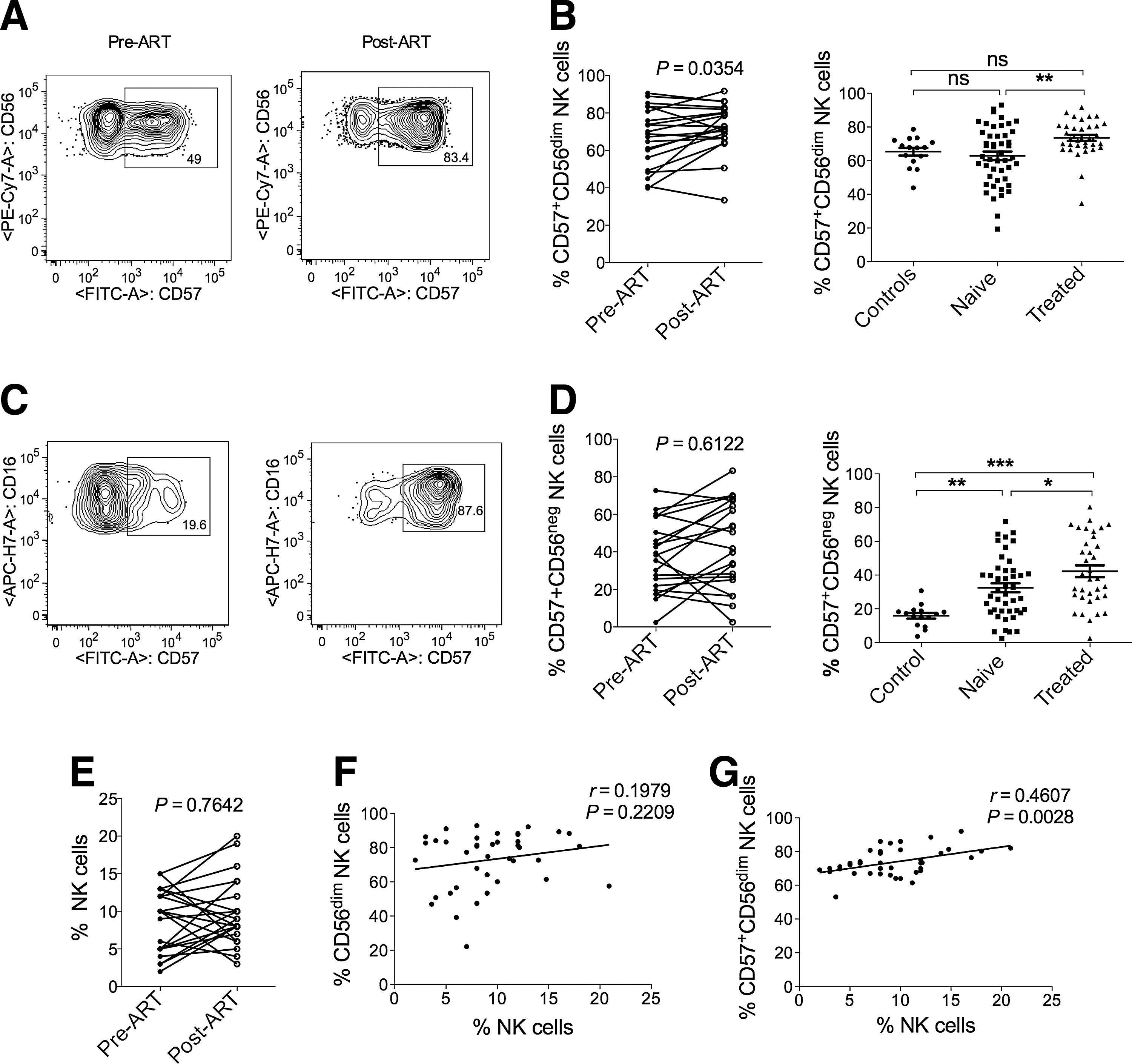
Terminal differentiation of NK cell subsets before and after treatment.
Increased frequencies of CD57 expressing CD56dimCD16+ NK cells is associated with higher NK cell frequencies after ART
As shown in Fig. 3B, some of the patients exhibited increased while others had decreased frequencies of CD57+CD56dimCD16+ NK cells, which was also true for the frequency of total NK cells (Fig. 3E). Therefore, we wanted to examine whether the increase in CD56dimCD16+ NK cells or in CD57+CD56dimCD16+ NK cells is associated with total NK cell frequencies after ART. To this end, we performed Pearson's correlation analysis of NK cell frequencies with the frequencies of CD56dimCD16+ (Fig. 3F) as well as with the frequencies of CD57+CD56dimCD16+ NK cells (Fig. 3G). We found no correlation between the frequencies of CD56dimCD16+ NK cells and NK cells. However, the frequencies of CD57+CD56dimCD16+ and NK cells correlated significantly (Fig. 3G), suggesting that the increase in CD57+CD56dimCD16+ NK cells is associated with a higher frequency of NK cells after ART.
Discussion
Previous studies reported that throughout the course of HIV disease progression a number of phenotypic and functional alterations occur in the NK cell compartment. 35 However, the total NK cell frequencies as well as absolute numbers remain stable in HIV-1-infected individuals compared to healthy controls, suggesting reallocation of NK cells from one compartment to another during HIV infection. After effective ART, these phenotypic and functional changes again are fully or partially restored similar to healthy controls. 36 Nevertheless, some of the phenotypic markers had a different outcome after ART. 35 Although terminal differentiation and activation of NK cell have already been reported in HIV-infected individuals, 11,13,17 they have not yet been studied under ART. Here we describe the effect of ART on terminal differentiation as well as on activation of NK cell subsets longitudinally as well as cross-sectionally.
Our data revealed that the frequencies and absolute numbers of CD56brightCD16+/– NK cells remain stable in treatment-naive patients compared to healthy controls as well to those treated with ART. This is in contrast to chronic HCV infection where CD56brightCD16+– NK cells expand in treatment-naive patients compared to healthy controls. 37 We also demonstrate overall similar absolute CD56dimCD16+ NK cells counts in post-ART HIV patients compared to pre-ART patients in the longitudinal study. Thus, the unrestored absolute numbers of CD56dimCD16+ NK cells in the cohort of post-ART HIV patients could represent a reflection of the low absolute NK cell counts. Furthermore, we could show a decrease in the frequencies or absolute numbers of CD56−CD16+ NK cells with restored frequencies of CD56dimCD16+ NK cells after ART (Fig. 1F and Supplementary Fig. S1C).
Notably, frequencies or absolute numbers of CD56brightCD16+/– NK cells do not show a significant correlation with the frequencies or absolute numbers of CD56−CD16+ NK cells. However, frequencies of CD56dimCD16+ and CD56−CD16+ NK cells correlate significantly in both treatment-naive and in ART patients. This suggests that HIV-1 viremia is associated with a pathological redistribution rather than an absolute decrease in the numbers of circulating NK cells and CD56−CD16+ NK cells might emerge from the CD56dimCD16+ NK cell compartment after CD56 downregulation; however, there is no clear evidence for this. The absolute numbers of CD56dimCD16+ and CD56−CD16+ NK cells do not show any significant correlation, possibly due to higher or lower initial absolute NK cell numbers that reflect lower or higher absolute CD56dimCD16+ and CD56−CD16+ NK cell numbers. Unfortunately, due to lack of clinical data of the healthy controls we were not able to compare absolute numbers of different NK cell subsets in the cross-sectional study.
Immune activation may be speculated to be one of the major players in driving the expansion of CD56−CD16+ NK cells. However, there was no direct correlation between expansions of CD56−CD16+ NK cells and the immune activation, suggesting a bystander effect of immune activation on CD56−CD16+ NK cell expansion.
Currently, systemic immune activation is considered to be the main driving force of CD4+T cell decline and development of acquired immune deficiency syndrome (AIDS) in HIV-1-infected individuals. 11,38 We observed immune activation of NK cells declining after ART, although it does not reach the level of healthy controls, suggesting that immune restoration may require more time. Nevertheless, the level of immune activation of CD56−CD16+ NK cells restored faster (Fig. 2C and D) than CD56dimCD16+ NK cells (Fig. 2A and B), which is also supported by our longitudinal study. The residual immune activation may persist due to residual ongoing viral replication.
CD57 expression defines a functionally stable NK cell subpopulation that increases with age and is consistently associated with a better outcome in cancer and autoimmune diseases. 39 We observed that CD56dimCD16+ and CD56−CD16+ NK cells show elevated CD57 expression after ART, suggesting a more differentiated phenotype. The longitudinal study data of CD57 on CD56−CD16+ cells did not reach the significance level perhaps due to low patient numbers. Increased frequencies of CD57+CD16+ NK cells were first reported in the individuals infected with human cytomegalovirus (HCMV), which drive the expansion of NKG2C+ NK cells that have preferentially acquired CD57. 40 Expansion of NK cells is also reported during hantavirus infection, which is restricted to the NKG2C+ subset coexpressing CD57 and KIRs. 41
In contrast, significantly reduced numbers of CD57+ NK cells were also reported in some autoimmune diseases such as alopecia areata, atopic dermatitis, and Sjögren's syndrome. 39 However, our cohorts did not show any manifestations of HCMV or hantavirus coinfection at the time of participation in the study. When we divided our cohorts into two groups based on the frequencies of CD57+CD56dimCD16+ NK cells after ART, we found those patients with increased frequencies of CD57+CD56dimCD16+ NK cells also presented increased total NK cell frequencies after ART. The correlation between the frequencies of CD57+CD56dimCD16+ NK cells and total NK cells suggests an association of terminal differentiation with the total NK cells after treatment. Expression of CD57 on NK cells may be associated with age, duration of the treatment, or possibly better immune recovery. However, we did not find significant differences in age, treatment duration, or CD4 counts between the two patient groups.
In summary, immune activation and disturbed distribution of NK cells are restored to normal levels after ART, but there is an increased terminal differentiation of CD56dimCD16+ NK cells reflected in the overall increase in NK cell frequencies after ART.
Footnotes
Acknowledgments
The authors acknowledge Margot Zielinska-Skowronek for collecting the PBMCs. F.A. is supported by the MD/PhD program of the Hannover Biomedical Research School (HBRS) and DZIF TTU 04.802. R.E.S. is supported by Grant IND 06/20, NEAT, and DZIF TTU 04.802. D.M.T. and R.J. are supported by SFB738/A5.
F.A. and R.E.S. conceived the study and designed the experiments. F.A., D.M.T., and N.M. performed the experiments. F.A. and D.M.T. analyzed the data. F.A., R.J., and R.E.S. prepared the manuscript. All authors read and commented on the manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.