
Abstract
The CCR5 coreceptor amino terminus and extracellular (ECL) loops 1 and 2 have been implicated in HIV-1 infections, with species differences in these regions inhibiting zoonoses. Interactions of gp120 with CD4 and CCR5 reduce constraints on metastable envelope subunit gp41, enabling gp41 conformational changes needed for infection. We previously selected HIV-1JRCSF variants that efficiently use CCR5(Δ18) with a deleted amino terminus or CCR5(HHMH) with ECL2 from an NIH/Swiss mouse. Unexpectedly, the adaptive gp120 mutations were nearly identical, suggesting that they function by weakening gp120's grip on gp41 and/or by increasing interactions with ECL1. To analyze this and further wean HIV-1 from human CCR5, we selected variants using CCR5(HMMH) with murine ECL1 and 2 sequences. HIV-1JRCSF mutations adaptive for CCR5(Δ18) and CCR5(HHMH) were generally maladaptive for CCR5(HMMH), whereas the converse was true for CCR5(HMMH) adaptations. The HIV-1JRCSF variant adapted to CCR5(HMMH) also weakly used intact NIH/Swiss mouse CCR5. Our results strongly suggest that HIV-1JRCSF makes functionally critical contacts with human ECL1 and that adaptation to murine ECL1 requires multiple mutations in the crown of gp120's V3 loop.
HIV-1
CCR5's amino terminus (Nt) and extracellular loop (ECL) 1 and 2 regions contribute to coreceptor activity. 20 –31 Affinities of sCD4-gp120 complexes for CCR5 are weakened by Nt and ECL2 mutations. 22,24 –26,32 –37 Tyrosine sulfates in Nt enhance infection and sCD4-gp120 binding, 26,34,35,38 and tyrosine sulfated Nt peptide binds to the base of gp120 V3. 33,37 Additionally, antibodies to ECL2 block entry. 36,39 –41 Studies of chimeric human CCR5s with substitutions from murine CCR5 or other chemokine receptors also suggest involvement of Nt and ECL1 and 2. 21,23,24,27,30,42 African green monkeys (AGMs) have been endemically infected by SIVAGM at high prevalence for millennia and their CCR5s contain many polymorphisms at functionally important sites in Nt, ECL1, and ECL2. 27,43,44
Damaging mutations in CCR5 can be overcome by adaptive mutations in HIV-1JRCSF gp120 centered in V3. 14 –16,44,45 Surprisingly, as described previously and summarized below, mutations adaptive for CCR5(Δ18) with a deleted Nt or CCR5(HHMH) with ECL2 from NIH/Swiss mice were overlapping, with S298N and F313L in V3 and elimination of an N-glycan at N403 (by substitutions N403K,S or T405N,A) in V4 being common. 45 These common mutations increased syncytia formation and susceptibilities to sCD4 inactivation and reduced the activation energy barrier that restricts gp41 refolding, thereby enabling weak coreceptors to function efficiently. 45 Conceivably, these common mutations might strengthen gp120 interactions with ECL1, thereby compensating for reduced reliance on Nt and ECL2.
A major goal of our investigation has been to wean HIV-1JRCSF from dependency on human CCR5 by adapting it in incremental stages for utilization of NIH/Swiss mouse CCR5. In addition, this approach provides evidence concerning the interactions of specific gp120 amino acids with sites in CCR5. To investigate these issues, we used previous methods. 14 –17,27,44 –46 We made CCR5(HMMH) by substituting the MscI–BsaBI fragment from the cDNA of NIH/Swiss mouse CCR5 for the corresponding human sequence (Fig. 1A), and used the HI-J clone of HeLa-CD4 cells to derive subclones with distinct cell surface CCR5(HMMH) amounts measured using monoclonal antibody 3A9 to human Nt. 47
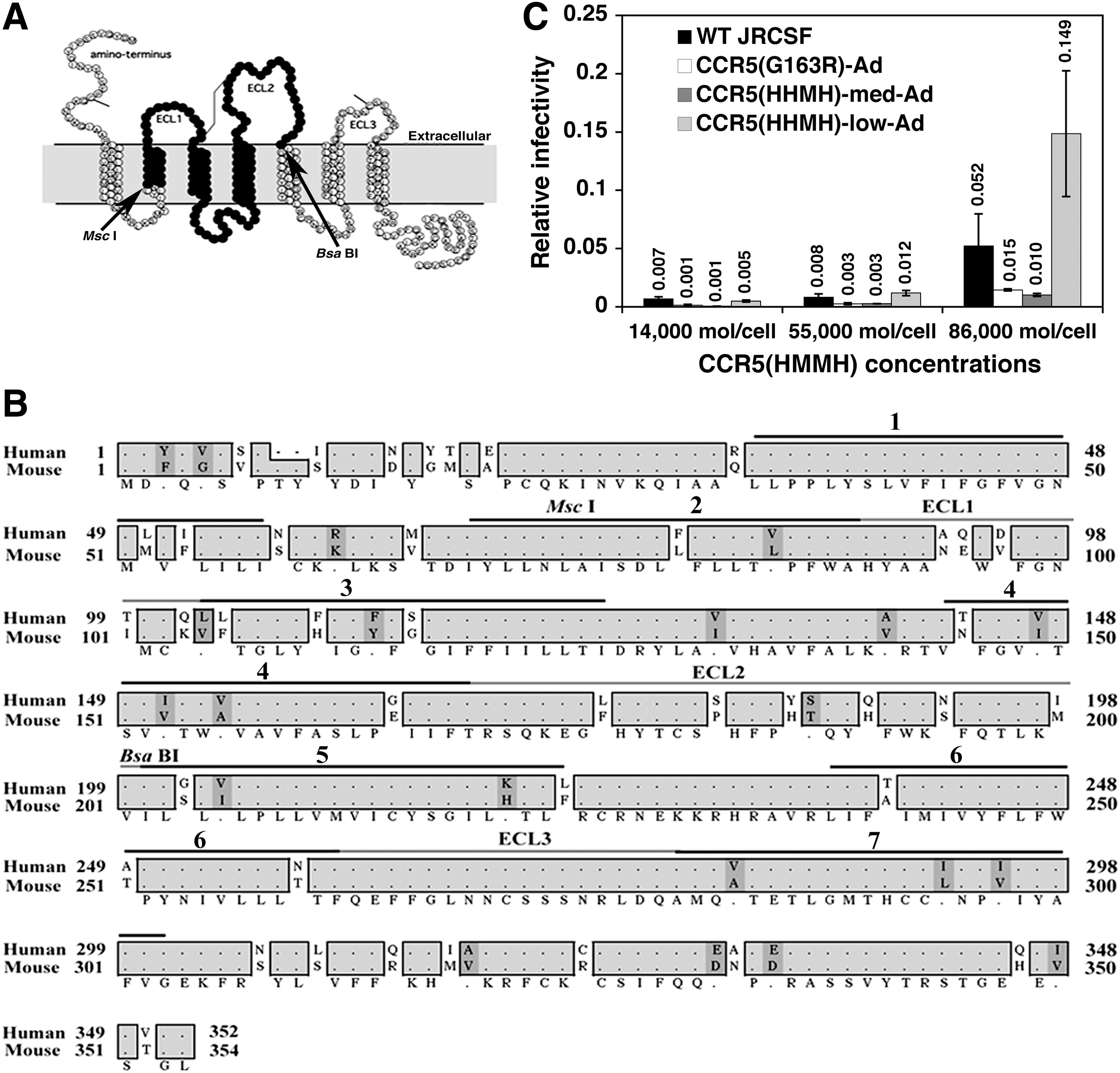
Construction and coreceptor activity of CCR5(HMMH).
Human and mouse CCR5 sequences differ in Nt, ECL1, and ECL2, but are identical in ECL3 except for conservative N-to-T and V-to-A substitutions at the TM6-ECL3 and ECL3-TM7 junctions (Fig. 1B). We initially assumed that HIV-1JRCSF variants adapted to CCR5(HHMH) with murine ECL2 and CCR5(G163R) that disrupts gp120 binding and occurs at the TM4/ECL2 junction 36 would be partially adapted to CCR5(HMMH). However, these variants could not use CCR5(HMMH) at low or medium concentrations although they weakly used a high concentration (e.g., ∼86,000/cell) (Fig. 1C), confirming that HIV-1s use coreceptors in a concentration-dependent manner. 16,44,45 JC.53 cells, which contain a supersaturating number (∼130,000/cell) of human CCR5 and are infected efficiently by all HIV-1 variants, were used to normalize titers of our virus preparations. These results suggested that HIV-1JRCSF mutations adaptive for CCR5(G163R) and CCR5(HHMH) were not partially adapted to use CCR5(HMMH) and might be maladaptive for use of murine ECL1.
Isolation of adapted variants occurs optimally when the initial virus replicates weakly in the cells used for selection. Accordingly, we used the HeLa-CD4/CCR5(HMMH) clone with ∼86,000 coreceptors/cell and the HIV-1JRCSF(G163R)-adapted virus that infects these cells ∼1.5% as efficiently as JC.53 cells (see Fig. 1C). After five cell passages a virus emerged (variant A) that we used to select variant B that replicates efficiently in cells with ∼10,000 CCR5(HMMH)/cell. This selection was very difficult and required 15 cell passages. Figure 2A shows infectivity assays of the A and B variants and several other HIV-1JRCSF variants previously adapted to other mutant CCR5s. Interestingly, viruses adapted to CCR5s containing human ECL1 used a high concentration of CCR5(HMMH) inefficiently and were unable to use low concentrations. Conversely, viruses adapted to CCR5(HMMH) cannot use CCR5(HHMH) or CCR5(Y14N) containing human ECL1. Importantly, none of the viruses uses CXCR4 (Fig. 2A). Figure 2B lists the previously isolated viruses that we employed in this investigation and identifies the specific adaptive env mutations that they contained.

Characterization of HIV-1JRCSF variants adapted for use of CCR5(HMMH).
Interestingly, variant B also efficiently used CCR5(HMMM), which differs from mouse CCR5 only in Nt, 27 and weakly used intact murine CCR5 in HEK293T cells that had been cotransfected with expression vectors for CD4 and CCR5s (Fig. 2C). In contrast, other tested viruses lacked this capability. The A and B variants were also less syncytium inducing in JC.53 cells than the highly fusogenic variant with S298N, F313L, and N403S that was adapted for efficient use of CCR5(HHMH) (Fig. 2D). In contrast to the latter virus, the A and B variants were also unable to infect HeLa-CD4/CCR5(Δ18) cells in the presence of the tyrosine sulfated Nt peptide (Fig. 2E). This suggests that CCR5(HMMH) adaptations did not enhance viral reliance on human Nt. Although gp120 adaptations to damaged CCR5 Nt and ECL2 substantially overlap (see Fig. 2B) and neither of these regions is essential, HIV-1 infection evidently requires specific interactions with ECL1.
The gp120-gp41 sequences of env cDNA clones from adapted viruses A and B are summarized in Table 1. Importantly, mutations adaptive for CCR5s with human ECL1 but damaged Nt or ECL2 were absent in CCR5(HMMH)-adapted variants except for the N-glycan loss mutation N403D in V4 in a proportion of A and B gp120s. The V3 mutation F313I in the initially employed CCR5(G163R)-adapted virus also reverted during adaptation to CCR5(HMMH). The A and B viruses both contain two viral populations, all having the I307L V3 substitution. The highly adapted B viruses also contain I305V plus G317E in V3 or R311G in V3 plus N403D in V4. These mutations suggest that loss of positive charge (R311G) or gain of negative charge (G317E) in V3 is adaptive for mouse ECL1 and maladaptive for human ECL1. Unlike our previously isolated N-glycan loss mutations (N403K,S or T405N,A), N403D also adds a negative charge. These charge differences may be important because murine ECL1 is positively charged (Fig. 1B) and because electrostatic interactions strongly influence coreceptor choice. 18,48,49
Entire envelope genes, encoding gp120/gp41, were obtained by PCR of genomic DNA harvested from infected cells and individual clones were then sequenced.
We have used HIV-1JRCSF numbering. The corresponding HIV-1HXBc2 numbering for the adapted envelope mutations is 219T, 307V, 309L, 314G, 320E, 411D, and 532T.
Envelope clones isolated from variant A virus-infected HeLa-CD4 cells expressing large amounts of CCR5(HMMH).
Envelope clones isolated from variant B virus-infected HeLa-CD4 cells expressing low amounts of CCR5(HMMH).
These V3 mutations cluster in the structures of sCD4-gp120 monomers 11 and gp120-gp41 trimers 50 (Fig. 3). Importantly, the V3 substitutions controlling ECL1 usage occur in gp120's V3 crown. The conservative V3 substitutions I305V and I307L occur in a conserved hydrophobic cluster that influences the orientation and packing of V3. Mutation R311G alters the highly conserved GPGR consensus sequence in the crown tip of V3. 51 The N403D N-glycan loss mutation is situated near positive charges in the V3 base of the unliganded trimer structure. Although some B variant clones contained mutations in C2 (A217T) and in the gp41 fusion peptide (A524T), they occur in regions not known to interact with CCR5 and were not present in a coherent pattern in the B clones. Consequently, we presume they may have been caused by genetic drift during the prolonged selection process. In contrast, the other A and B virus mutations were located specifically in gp120 sites previously shown to interact with CCR5. 15,16,45
Locations of the CCR5(HMMH)-adaptive mutations in the monomeric and trimeric crystal structures of gp120.
These experiments suggest that the gp120 mutations adaptive for both CCR5(HHMH) and CCR5(Δ18) (i.e., S298N, F313L, and N-glycan loss at N403) (Fig. 2B) make the virus more reliant on human ECL1 and consequently less tolerant of mutations in this region. Previous mutagenic studies also strongly imply that CCR5 ECL1 plays an important role in HIV-1JRCSF entry. 27 Thus, the gp120 V3 loop mutations that were adaptive for CCR5(HHMH), CCR5(Δ18), and CCR5(G163R), which contain human ECL1, were maladaptive for use of murine ECL1. Conversely, the A and B CCR5(HMMH) HIV-1JRCSF variants adapted for murine ECL1 were maladapted for use of human ECL1.
Whereas gp120 mutations adaptive for CCR5s with damaged Nt or ECL2 are substantially identical, implying that gp120 interactions with these regions cooperate in a common process, we conclude that gp120 interactions with CCR5 ECL1 have a functionally different role that is essential for infection. We propose that binding steps involving Nt and ECL2 precede the ECL1-dependent entry process that involves its interactions with the gp120 V3 crown. This interpretation is concordant with previous evidence of Cormier et al. 37
Footnotes
Acknowledgments
This research was supported by NIH grant CA67358 from the National Cancer Institute. We thank S. Kozak and L. Schwanemann for technical assistance. Author contributions: E.J.P. and D.K. planned the overall project, discussed experimental results, and wrote the article. E.J.P. planned and performed all experiments and collaborated with J.P.D. in constructing the CCR5(HMMH) chimeric coreceptor and generating cell clones.
Author Disclosure Statement
No competing financial interests exist.