
Abstract
The p21/CDKN1A protein has been described in vitro as well as in a small subset of patients as a restriction factor for HIV infection. We evaluated p21/CDKN1A mRNA expression on CD4+ T cells from HIV-infected individuals with two outcomes (18 elite controllers and 28 viremic progressors). Our results show broad interindividual variation in this factor, which is unrelated to the patient's phenotype. Considering the gene's genetic surroundings in chromosome 6, such as HLA genotype and single nucleotide polymorphisms (SNPs), there was a positive association with carrying HLA-B2705 alleles and the rs733590 SNP. Thus, this natural variation of p21/CDKN1A alone does not appear to be a prognostic indicator of effective viral control in vivo and other factors must be considered.
Introduction
T
A recent report showed that HIV infection alone increased expression of p21, indicating an additional effect on HIV induction. 8 In vitro studies showed that p21 gene expression was upregulated by the HIV Vpr to stall the cell cycle for viral benefit. 9,10 All these studies support the hypothesis of a dual role for p21, which depending on the immune status of the patient and homeostasis of T cell subsets would have either a negative or positive effect on the course of infection, as seen in other pathologies. 11
The genetic environment may influence the gene's expression. In other diseases, the presence of some single nucleotide polymorphisms (SNPs) influences the expression of p21. 12,13 These SNPs either have a direct effect on p21 expression—some of them located within the gene promoter and coding regions—or act as tagging-SNPs for another involved gene. In addition, the p21/CDKN1A gene is within close proximity of the MHC class I genes on chromosome 6, which is a region with a strong linkage disequilibrium and a high incidence of polymorphisms. This situation introduces more complexity into the equation and the contradictory results obtained so far warrant more studies to obtain a clearer picture of the overall HIV-infected population. Therefore, we aimed to evaluate p21/CDKN1A expression and the patient's genetic background, which might influence its expression in two groups of HIV-infected patients (elite controllers and patients with active HIV replication).
Materials and Methods
Patients and sample processing
We evaluated 18 HIV EC (HIV-infected patients who control viremia without receiving antiretroviral treatment) and 28 HIV viremic patients (VP) (patients with a detectable viral load) from the Hospital Carlos III cohort. The ethical committee of Hospital Carlos III approved the study and all patients signed the informed consent document. The criteria for EC selection included HIV-1 infection with sustained undetectable plasma viral load for several years in the absence of antiretroviral treatment (only sporadic blips of viremia <500 copies/ml were allowed). VPs were selected based on continuous detectable viremia and a clear CD4+ T cell decline; samples from these later patients were obtained prior to antiretroviral treatment. The CD4+ T cells were positively isolated from peripheral blood mononuclear cells (PBMCs) using specific antibody-coated magnetic beads (Dynabeads CD4, Invitrogen). These cells were processed to extract large size RNA (>200 nt) and genomic DNA using AllPrep RNA/DNA by Qiagen.
Analysis of CDKN1A mRNA expression
Reverse transcription was performed on the RNA fraction (approx. 1 μg RNA) using random primers and the AMV-RT Access kit from Promega. The conditions were 95°C for 5 min before adding AMV-RT, followed by incubation at 25°C for 10 min, 40°C for 30 min, 48°C for 30 min, and a final inactivation step of 80°C for 2 min. cDNA was amplified by qPCR using the CDKN1A TaqMan Gene Expression Assay from Life Technologies. Relative levels of mRNA were calculated by the ΔΔC t method using β-actin as the reference constitutive gene. All measurements were normalized to the mean value of two healthy donors.
Genotyping
SNP typing was performed on DNA samples using the TaqMan SNP Genotyping Assays for rs733590, rs2395655, and rs1801270 (Life Technologies). HLA class I-B typing was done by INNO-LIPA HLA-B update Plus (Fugiribio) and PCR-SSO using Luminex technology (Gen-Probe).
Statistical analysis
Continuous variables were expressed as median and interquartile range and categorical variables as counts and percentages. Given the sample size and the nonnormal distribution of most variables, nonparametric tests were used in most of the analyses. The 0.05 p-value cut-off was used for significance. The deviation of the observed frequency of haplotypes (D′) and the Spearman correlation were calculated and the chi-squared test was performed for linkage disequilibrium (LD) and haplotype analysis using Haploview 4.2 software. 14
Results
When we analyzed the two groups of patients with different phenotypic characteristics (Table 1), we found that p21/CDKN1A mRNA levels were heterogeneous among individuals within both groups, with overall differences ranging from 0.06-fold to 20.32-fold change with respect to a reference sample. For the VP group mRNA fold changes ranged from 0.06 to 20.32 and for the EC patients differences went from 0.22 to 10.45. Overall, the p21/CDKN1A levels in EC participants were slightly increased but not statistically significant (p = 0.083) (Fig. 1). If we removed the outlier in the VP group, this difference reached significance (p = 0.046); however, except for a few patients, the differences with the VP group were moderate (EC patients had a median of 1.53 and IQR = 0.95–2.26, while VP patients had a median of 0.72 and IQR = 0.43–1.73) and both groups had an important overlap in expression levels. The levels of p21/CDKN1A and the patients' CD4+ and CD8+ T cell counts showed no association (Spearman Rho = –0.19, p = 0.902 and Spearman Rho = 0.002, p = 0.992; respectively), nor did the CD4+/CD8+ ratios (Spearman Rho = −0.161, p = 0.379). In addition, no correlation was found between viral load and p21/CDKN1A expression (Spearman Rho = 0.29, p-value = 0.846).
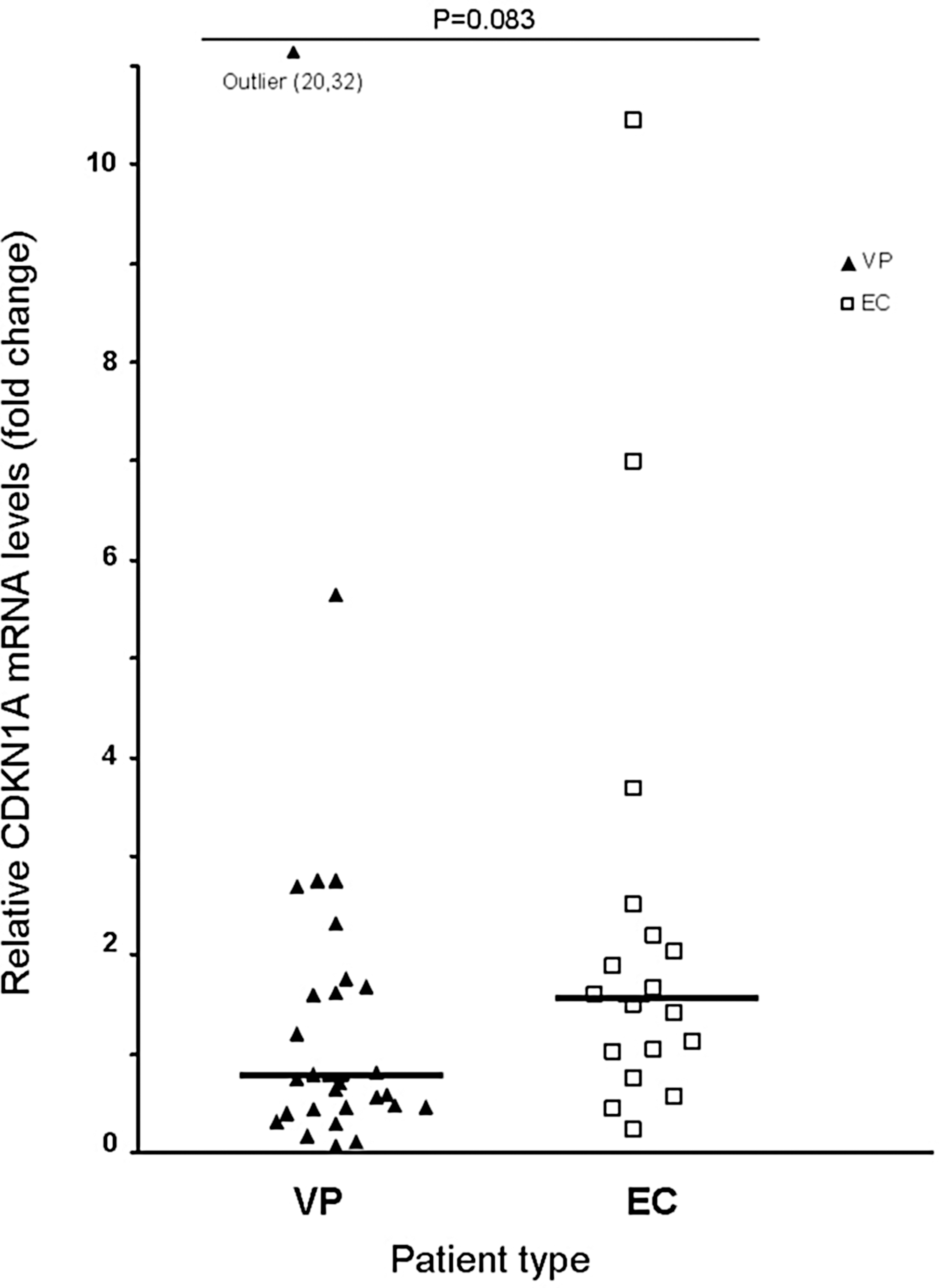
Relative CDKN1A mRNA levels in CD4+ T cells from VP (▴) and EC (□) patients. Horizontal bars indicate median values. p values were calculated using the Mann–Whitney or Kruskal–Wallis tests, as appropriate.
p-values <0.05, Fisher's exact or Mann–Whitney tests, as appropriate.
Medians (IQR) or N (%) for each variable are indicated.
HLA-typing of all patients showed that EC patients carried more protective major histocompatibility complex (MHC) class I alleles: HLA-B*2705 and HLA-B*5701 than VP [EC: 10 (55%) vs. VP: 5 (18%), p = 0.005]. In the EC patients, the difference in protective HLA-B alleles was skewed toward the HLA-B*2705 genotype. The p21/CDKN1A mRNA expression showed a positive association with carrying protective HLA-B alleles [protective versus nonprotective, medians (IQR): 1.92 (1.09–4.7) vs. 0.72 (0.43–1.57), p = 0.002]. This association seems driven by the presence of the HLA-B*2705 allele [medians (IQR): 2.67 (1.14–6.65) vs. 0.74 (0.44–1.7), p = 0.007, respectively] more than the HLA-B*5701 allele [medians (IQR): 1.59 (0.55–2.5) vs. 0.8 (0.45–1.75, p = 0.468] (Fig. 2a and b). We did not observe differences in p21 mRNA expression between EC and VP when all patients carrying a protective HLA-B alleles were excluded from the analysis [medians (IQR): 1.02 (0.44–1.48) and 0.64 (0.41–1.64), p = 0.559, respectively].
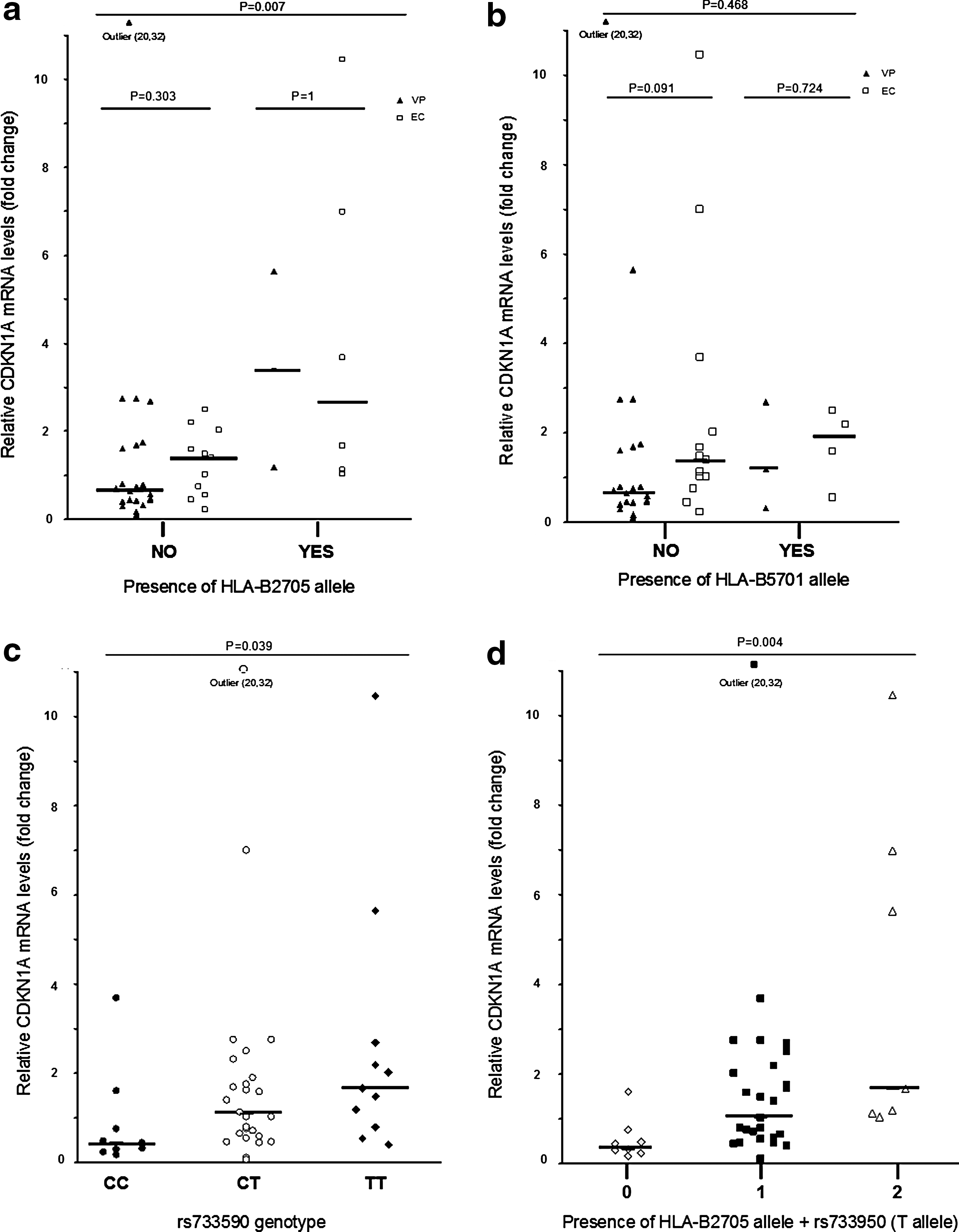
Relative CDKN1A mRNA levels in CD4+ T cells from
We also determined three regulatory polymorphisms related to p21/CDKN1A expression. None of these polymorphisms was in linkage disequilibrium with the others in our pooled population. Only the rs733590 SNP was associated with p21/CDKN1A expression. Carriers of the major T allele, either in heterozygosis or homozygosis, showed higher levels of mRNA (Fig. 2c). The medians (IQR) of p21/CDKN1A expression were 0.43 (0.88–1.13), 1.08 (0.57–1.99), and 1.66 (0.8–2.69) for CC, CT, and TT genotypes, respectively. We did not detect differences in this SNP genotype frequency between EC and VP patients, and when we separately tested the SNP in each group, the same trend in p21 expression was observed.
In the analysis of the combined effect of HLA-B and rs733590 genotypes, individuals who carried both HLA-B*2705 and rs733590 (T) alleles had a greater expression of p21/CDKN1A mRNA than individuals carrying only one or neither (Fig. 2d). Exploring this association further, a two-way ANOVA for the analysis of rank variance of p21 showed a certain interaction effect of HLA-B2705 on the rs733590 T allele by modifying p21/CDKN1A expression, but the analysis did not reach statistical significance [F(1, 38) = 2.41, p = 0.129]. Moreover, despite being in close proximity within the same chromosome, both genotypes were not in LD, indicating independent inheritance. This was also confirmed by the analysis of the rs733590 major T allele frequency in patients carrying the HLA-B*2705 allele, which did not differ from the frequency observed in the general European population (0.62 vs. 0.6, respectively). However, patients not carrying the HLA-B*2705 allele showed a higher frequency of the rs733590 minor C allele compared to the general European population (0.59 vs. 0.4, respectively).
Discussion
In this study, we observed a natural variation in mRNA expression levels of p21/CDKN1A between individuals, independently of disease stage, and although we detected a slight overall increase of expression in the elite controller group, differences between the groups did not meet conventional levels of statistical significance. This diversity in expression has also been seen in healthy donors in other studies. 12,15 This result would indicate a more complex relationship of this restriction factor with HIV infection, which is in accordance with other studies. Due to its complex set of biological functions and various regulatory pathways, the p21/CDKN1A protein has been shown to exert dual positive and negative activities toward the cell cycle in a context-dependent manner, including intrinsic and extrinsic signals. 16 In HIV infection, p21/CDKN1A could be either exploited by the host as an antiviral defense or by HIV itself to counteract this defense. 17 Other groups have also found similar levels of p21/CDKN1A in elite controllers and patients in various HIV disease stages or in activated HIV-infected PBMCs from various patients. 2,15,18 It has been speculated that the mere presence of viral antigens triggers the expression of p21/CDKN1A 19 ; however, we did not find any correlation with viral load.
HIV causes abnormal T cell homeostasis and p21/CDKN1A seems to be involved in its regulation, especially in the control of activated/memory T cell expansion and tolerance. 20 Along these lines, recent reports have identified the action of p21 on several host proteins such as RNR2 and SAMHD1 and its effect on restricting HIV by modulating the cell status. 5,6,21 The only parameter we had available from our cohort to test T cell status was the CD4+/CD8+ T cell ratio—which some studies have linked with T cell perturbations and immune activation 22,23 —and although we did not find any correlation, we do not dismiss the observation that the p21/CDKN1A levels observed in our patients likely reflect the fact that its expression is driven by immune activation and the T cell status of each patient.
Since the p21 gene is localized in the 6p21.2 region of chromosome 6 and is close to the HLA genes, we also explored a possible relationship with some of the protective HLA-B alleles that are overrepresented in the EC population, such as HLA-B*5701 and HLA-B*2705. We found differences in p21/CDKN1A expression only in patients carrying the HLA-B*2705 allele. We cannot rule out a possible effect of the immune cellular response in these differences since the HLA-B*5701 numbers are small. On the other hand, this association observed with HLA-B*2705 and not with the most protective HLA-B*5701 allele—as seen in other studies 19 —may suggest a certain influence of the genetic background. We also studied some selected SNPs located within the p21 promoter region that could alter its expression and observed an association with the rs733590 SNP. 12
The presence of the T allele is associated with increased levels of p21 mRNA, which is in agreement with the results obtained by Korthagen's group in a white population. 12 The rs733590 does not seem to be genetically associated with the HLA-B*2705, but we cannot rule out a possible cofounding effect of one on the other. The univariate analysis of variance suggested some effect, but the numbers are too small to be conclusive. In this sense, the fact that patients without the HLA-B*2705 allele showed a higher frequency of the minor C rs733590 allele, which is associated with a lower p21/CDKN1A expression in our study, supports this possible interaction and could act as a tag-SNP linked to a functional SNP in another part of the gene or chromosomal region.
In conclusion, the natural variation of p21/CDKN1A, considered independently of other cellular and immunologic patterns, does not appear to be a prognostic indicator of effective viral control in vivo. The equation must include additional factors.
Footnotes
Acknowledgments
We thank Juliette Siegfried for manuscript revision. This work was supported by grants from the Dirección General de Farmacia, Ministerio de Sanidad, Spain DG11/205 (B.R.), Instituto de Salud Carlos III-Subdirección general de evaluación y fomento de la investigación (PN I+D+I 2008–2011), Ministerio de Economia y Competitividad, Spain and ERDF: PI12/00850 (B.R.), and CA12/00333 (A.P.).
Author Disclosure Statement
No competing financial interests exist.