
Abstract
Epithelial cells in human cervical and colonic mucosa do not express HIV receptor. However, HIV transmission occurs across the unbreached epithelia by an unknown mechanism. In this study, the effect of HIV exposure on tight junction (TJ) and cytokine production in ectocervical and colon mucosal epithelia in tissue biopsies was investigated in an organ culture model. After HIV exposure, the distribution patterns and quantities of epithelial TJ and adherens proteins were evaluated by immunofluorescence staining followed by confocal microscopy. Cytokine mRNA in the mucosal epithelia was also evaluated by real-time reverse transcription–polymerase chain reaction (RT-PCR). HIV transmission was evaluated by measuring p24 production in culture supernatant. Our results showed there were no significant changes in the distribution and quantities of epithelial TJ/adherens junction (AJ) proteins after exposure to HIV. However, higher levels of CXCL10 and CXCL11 mRNA expression were detected in HIV-exposed ectocervical epithelia. In case of colon mucosa, higher levels of CXCL10 and IL-6 mRNA expression were detected in HIV-exposed colon mucosa. Our study suggests that HIV induces cytokine production in epithelial cells, which may facilitate HIV transmission by recruiting HIV target cells in the submucosal region. Furthermore, HIV transmission may not occur through epithelial TJ/AJ disruption.
Introduction
W
We and others have shown that viral particles are less likely to be transcytosed through the outer layers of squamous epithelium. 3,4 Regardless of the mechanism of HIV transmission through epithelial layer, the virus passes through these protective layers and infects the underlying CD4+ T cells. The exact mechanism facilitating HIV transmission through the mucosal epithelia is still not well understood.
Epithelial cells are joined by intercellular junctions containing distinctive combinations of specialized molecules such as tight junction (TJ), adherens junction (AJ), and desmosomes. 5 TJ is an intercellular junctional structure that facilitates cell–cell adhesion and plays an important role in epithelial cell function. 6 TJ comprises various transmembrane proteins such as claudin and occludin, and also cytosolic proteins such as the zona occludens (ZO) that act as adapter molecules. 5
The AJ, in contrast, comprises epithelial cadherin proteins (E-cadherin) that are linked to the cytoskeleton via vinculin and alpha and beta catenin. 7 TJ forms a barrier that restricts the transport of ions and nonelectrolytes through the extracellular clefts between cells (the “gate” function). It also serves to maintain cell polarity forming a “fence” that restricts the diffusion of proteins and lipids between apical and basolateral surfaces. 8 Na/K/ATPase expressed basolaterally also plays a crucial role in the development of epithelial polarity. 9
The disruption of TJ can lead to microbial translocation and inflammation in the mucosa. 10 Various pathogenic organisms such as rotavirus, astrovirus, and Escherichia coli have developed strategies to disrupt the TJ, leading to pathogenic conditions characterized by increased intestinal permeability. 11,12 TJ disruption and a decrease in transepithelial electrical resistance have been observed in human primary endometrial cells and intestinal cell lines after exposure to HIV. 10 However, very little data are available in the literature studying the effect of HIV on epithelial TJ in human ectocervical and rectal/colonic tissues, which are more indicative of in vivo conditions.
Once HIV crosses the epithelial barrier, the virus establishes infection in small focal “hot spots.” 1,13 Cervical and intestinal epithelial cells express Toll-like receptor (TLR)s 1–6 and TLRs 1–5 and 9, respectively, to recognize both bacterial and viral pathogenic motifs in the lumen. 1 TLR-mediated activation has been shown to induce cytokine and chemokine production, including CXCL10, IL-6, CXCL11, SDF-1, MIP-1α, MIP-1β, IL-8 TNFα, GM-CSF, Type 1 IFNs, and RANTES, which all play important roles in recruiting immune cells and enhancing their functions. 1,10 Various in vitro studies have shown that HIV induces production of cytokines and chemokines such as IL-8, MIP-1β, MCP-1, thymic stromal lymphopoietin (TSLP), TNFα, and IL-6 in genital and intestinal cell lines. 1,14
Brenchley et al. showed that the degree of inflammation within the GI tract correlated with viral replication and further observed that colonic mucosa from HIV-infected patients had significantly higher levels of proinflammatory cytokine expression (e.g., TNFα, IFN-γ, and IL-6) compared with control patients. 15,16 Studies have reported upregulation of TSLP in human cervical epithelial cell lines post-HIV exposure that strongly activates human myeloid dendritic cell (DC), 14 while others have shown accumulation of CD4+ T cells within 6 h post-HIV infection in human cervical tissue. 17 Therefore, epithelial cells may be the first target for HIV to induce local immune activation, which facilitates the establishment of initial HIV infection in the submucosal region.
In the current study, we investigated the effect of HIV exposure on TJ/AJ and cytokine/chemokine production in ectocervical and colon mucosal epithelia in ex vivo human ectocervical and colon tissue explants. Our results show that the distributions of TJ/AJ proteins in the mucosal epithelial region in ectocervical and colon tissues were not significantly changed after exposure to HIV. However, HIV exposure upregulated expression of certain chemokines and cellular factors in the mucosal epithelia. These induced chemokines may facilitate the transmission of HIV by increasing the target cell availability in the epithelial region.
Materials and Methods
Virus culture
PM1 cells, a T-cell line that expresses CD4, CXCR4, and CCR5, were used to propagate R5 tropic HIV BAL. Briefly, PM1 cells were maintained in RPMI 1640 (Mediatech) containing 100 U/ml penicillin, 0.1 mg/ml streptomycin, 20% (v/v) FBS, and 10 mm HEPES. PM1 cells were infected with HIV BAL (TCID50 1 × 107) for 3 h followed by washing to remove excess virus and then cultured for 15 days. Starting from day 3 postinfection, culture supernatant containing HIV was collected every other day and filtered through an Amicon Ultra-15 filter device (Millipore) to remove the soluble cytokines.
The filtered HIV BAL viral stock was then quantified using HIV p24 ELISA (p24 ElISA kit; SAIC-Frederick, Inc.) and its infectivity titer (TCID50/ml) was determined by titration on CD8 depleted peripheral blood mononuclear cell. The control culture supernatant was prepared in the same way except cells that were not infected with HIV. AT-2-inactivated HIV-1 ADA (R5 virus) was a gift from J. D. Lifson, National Cancer Institute, Frederick, MD.
Human colon and ectocervical tissues
Ectocervical tissues were obtained from premenopausal, HIV-negative patients with no history of sexually transmitted diseases who were undergoing hysterectomy at the Magee Women Hospital, University of Pittsburgh Medical Center. The colon tissues were obtained from HIV-negative patients undergoing surgical resection of colon with noninflammatory conditions at the University of Pittsburgh Medical Center. The Institutional Review Board of the University of Pittsburgh approved this study as an exempt study. Informed consent from every individual was waived because this study used tissues that were procured through the Tissue Procurement Facility with only generalized patient information such as age and race of the patient.
Ectocervical organ culture
To study TJ/AJ proteins, the organ culture was performed with ectocervical tissues as previously described. 3 Briefly, ectocervical biopsies (6 mm diameter) were placed into the wells of a 12-well transwell plate with the epithelial layer facing up and the edges were sealed with 3% agarose. Cell-free HIV (TCID50 of 106) or control supernatant was added to the epithelial layer of the tissue in the upper chamber and complete IL-2 media (RPMI 1640 media), 10% heat-inactivated fetal bovine serum, and 10 U/ml of recombinant human interleukin-2 (Roche) were added to the bottom well. Cultures were incubated at 37°C for 24 h in a CO2 incubator.
To evaluate the TJ gate function and also to examine the barrier properties of the TJ/AJ proteins after exposure to HIV, ectocervical biopsies were exposed to HIV or control supernatant in an organ culture for 24 h. The tissues exposed to control supernatant were treated with or without 10 mm EDTA in the final 2 h incubation time. HIV- or control supernatant-exposed tissues (with or without EDTA) were further exposed to FITC-labeled dextran (3 kDa) for 1 h at 37°C in a CO2 incubator.
To study cytokine/chemokine upregulation in ectocervical tissues, the epithelial layers of ectocervical tissues were exposed to either cell-free HIV BAL {(TCID50 of 106, GP120 0.8 nm (0.1 μg/ml), NIH AIDS reagent Catalog No. 11784, aldrithiol-2 (AT-2)-inactivated HIV (71 ng/ml)} or control supernatant in an organ culture. Cultures were incubated at 37°C for 24 h in a CO2 incubator. For all the previous experiments, the tissues after organ culture were frozen down in OCT at −80°C and cryosectioned (7 μm thickness) for subsequent studies.
To study HIV transmission, ectocervical tissues were inoculated with cell-free HIV BAL (TCID50 of 106) and cultured overnight in a CO2 incubator at 37°C. Following the incubation, the biopsies were washed and cultured again for additional 16 days. To monitor virus growth, the HIV p24 production was measured by the ELISA technique in the culture supernatant every 3 days during the culture period.
Colon organ culture
Organ culture using colon tissues was set up as described previously 18 with modifications. Tissues were collected and processed within 2 h of surgery. The tissues were dissected into 6 mm diameter biopsies and inoculated with cell-free HIV BAL (TCID50 of 106) or control supernatant for 6 h in a CO2 incubator at 37°C. After the incubation, the colon biopsies were either frozen down in OCT at −80°C for measuring cytokine mRNA or fixed in SafeFix II (Fisher Scientific) for 2 h, paraffin embedded, and sectioned (7 μm thickness) for immunostaining of TJ/AJ proteins.
To study virus transmission, colon biopsies were exposed to HIV BAL (TCID50 of 106) for 6 h. The biopsies were then washed and cultured for 16 days. Virus production was monitored every 3 days by HIV p24 measurement in the culture supernatant by ELISA.
Histological and immunofluorescence analysis of colon and ectocervical tissues
To examine the morphology of the mucosal epithelia, hematoxylin and eosin (H&E) staining of the colon/ectocervical tissues was performed after exposure to control supernatant or HIV. To study the effect of HIV on TJ/AJ, ectocervical and colon tissues were examined by immunofluorescence microscopy. Colon tissue sections were deparaffinized and antigen retrieval was performed by heating sections in a sodium citrate buffer (005000; Invitrogen) in a microwave, followed by cooling to room temperature. Tissues were washed with phosphate-buffered saline (PBS) followed by permeabilizing with Triton X-100 and blocking with a blocking buffer (PBS with 2% BSA) for 20 min.
After washes, the tissue sections were then treated with one of the following primary antibodies: rabbit polyclonal anti ZO-1, Claudin-1 (Invitrogen), mouse monoclonal anti Claudin-4, E-cadherin (Invitrogen), Na/K/ATPase (05–369; Millipore) at a dilution of 1:50, 1:75, 1:200, and 1:500, respectively, for 60 min. Tissue sections were stained in parallel with isotype control antibodies: negative control rabbit immunoglobulin (x0936; Dako) and mouse IgG1 isotype (556648; BD Pharmingen). Tissues were washed with 0.5% BSA in PBS and incubated with either goat anti-mouse Alexa 488 (Jackson Immunoresearch) or goat anti-rabbit Cy3 (Jackson Immunoresearch) at a dilution of 1:500 or 1:1,000, respectively, for 1 h. Nuclear counterstaining was done with DAPI. All sections were mounted by gelvatol and coverslips were sealed on slides.
In case of ectocervical tissues, sections were fixed with 2% PFA for 20 min and washed with 0.5% BSA in PBS. The tissue sections were then blocked with 2% BSA in PBS, treated with primary antibodies, secondary antibodies, DAPI, and sealed using coverslip as mentioned earlier for the colon tissues. To examine the permeability of dextran, dextran-exposed ectocervical tissue sections were stained with DAPI and sealed using coverslip as mentioned earlier.
Images were taken with the Olympus Fluoview 1000 confocal microscope or Nikon Eclipse E600 microscope using a 20 × or 40 × oil objective. For acquiring images on confocal microscopy, the identical image acquisition setting was used for control and HIV-exposed tissues. Two controls and two HIV-exposed biopsies were obtained from each donor and 5–10 separate, random images were obtained from each biopsy. Therefore, from each donor, there were 10–15 images from control biopsies and 10–15 images from HIV-exposed biopsies. Experiments were repeated in three donors, each time with two control biopsies and two HIV-exposed biopsies.
To study ZO-1, Claudin-4, E-cadherin, Na/K/ATPase protein expression in ectocervical epithelia, equal thickness of nucleated epithelial region from the basolateral membrane was selected as region of interest using NSI element software. Fluorescent intensity of the above proteins in the region of interest was measured using NSI element software maintaining the same threshold setting for an entire data set and the fluorescence intensity was normalized to the number of nuclei in the region. To examine ZO-1 nuclear colocalization in ectocervical epithelia, 30 nuclei (per image) in the epithelial region closer to basolateral membrane were randomly chosen per field and ZO-1 nuclear colocalization was analyzed using ImageJ program by maintaining the same threshold setting for an entire data set.
In case of colon tissues, cross-sectional images were obtained using the Olympus Fluoview 1000 confocal microscope using a 20 × or 40 × oil objective. Using NSI element software, the columnar epithelial cells surrounding the lumen were selected as region of interest. Fluorescent intensity of the ZO-1, Claudin-4, E-cadherin, and Na/K/ATPase proteins in the region of interest was measured using NSI element software as mentioned in ectocervical tissues.
Transmission electron microscopy
Human colon tissues were exposed to control supernatant or HIV for 6 h and fixed in 2.5% glutaraldehyde for 1 h at room temperature. The biopsies were washed with PBS and processed as previously described. 19 Briefly, biopsies were postfixed in aqueous 1% OsO4, 1% K3Fe(CN)6, dehydrated with ethanol series, infiltrated in a 1:1 mixture of propylene oxide:Polybed 812 epon resin (EBS Sciences), and epon for 1–3 h. This was further embedded in molds, cured, and ultrathin (60–80 nm) sections of the vitreous were collected on copper grids. Furthermore, they were stained with lead citrate and 2% uranyl acetate, and transmission electron microscopy (TEM) images were acquired using a JEOL JEM 1011 TEM at 80 kV fitted with a side-mount AMT 2k digital camera.
TaqMan® real-time PCR
The ectocervical and rectal/colonic epithelial layers from controls, HIV-, AT-2-, or GP120-exposed tissues were collected by microdissection followed by extraction of RNA using RNAzol B (TEL-TEST, INC.). The purity of the ectocervical epithelial regions obtained was assessed by the enrichment of cytokeratin 13 (Krt13) mRNA, which is predominately expressed in epithelial cells. The mRNA levels of Krt13 and cytokines/chemokines of CXCL10, CXCL11, IL-6, IL1β, IL-8, IL-10, TNFα, and IFNγ were measured in the microdissected epithelial layers by real-time reverse transcription–polymerase chain reaction (RT-PCR). Krt13 (endogenous control for epithelial cells), human CXCL10, CXCL11, IL-6, IL1β, IL-8, IL-10, TNFα, and IFNγ primers and probes labeled with FAM/MGB were purchased from Life Technology (gene assay ID: IL1β—Hs99999029_m1, IL-6—Hs99999032_m1, IL-8—Hs99999034, IL-10—Hs00961622_m1, TNFα—Hs99999043, IFNγ—Hs00989291_m1, CXCL10—Hs00171042_m1, CXCL11—Hs00171138_m1, Krt13—Hs00999762_m1). Real-time RT-PCR with gene-specific primers/probes was performed as described previously. 20 Briefly, RNA was reverse-transcribed with TaqMan® Reverse Transcription Reagents (Applied Biosystems) following the manufacturer's protocols.
A 25 μl PCR mixture consisting of 20 × TaqMan Pre-Developed Assay Reagents (Applied Biosystems), 5 μl cDNA (20 ng total RNA equivalent), and 2 × TaqMan Universal PCR Master Mix was used in all the reactions. Real-time PCR was carried out using ABI Prism 7000 Sequence Detection System under the following cycling conditions: 50°C for 2 min, 95°C for 10 min, 40 cycles of 95°C for 15 s and 60°C for 1 min. Results were expressed as fold changes relative to control.
Statistical analyses
TJ/AJ quantitation data are presented as mean ± standard error, and the Wilcoxon signed-rank test was used to determine statistical significance (p < .05). For mRNA comparisons between the tissues treated with HIV/GP120 or HIV/AT-2, T-test unequal variance analysis was performed with p value <0.05 for statistical significance.
Results
Effect of HIV exposure on the integrity of the cervical/colon mucosal epithelia
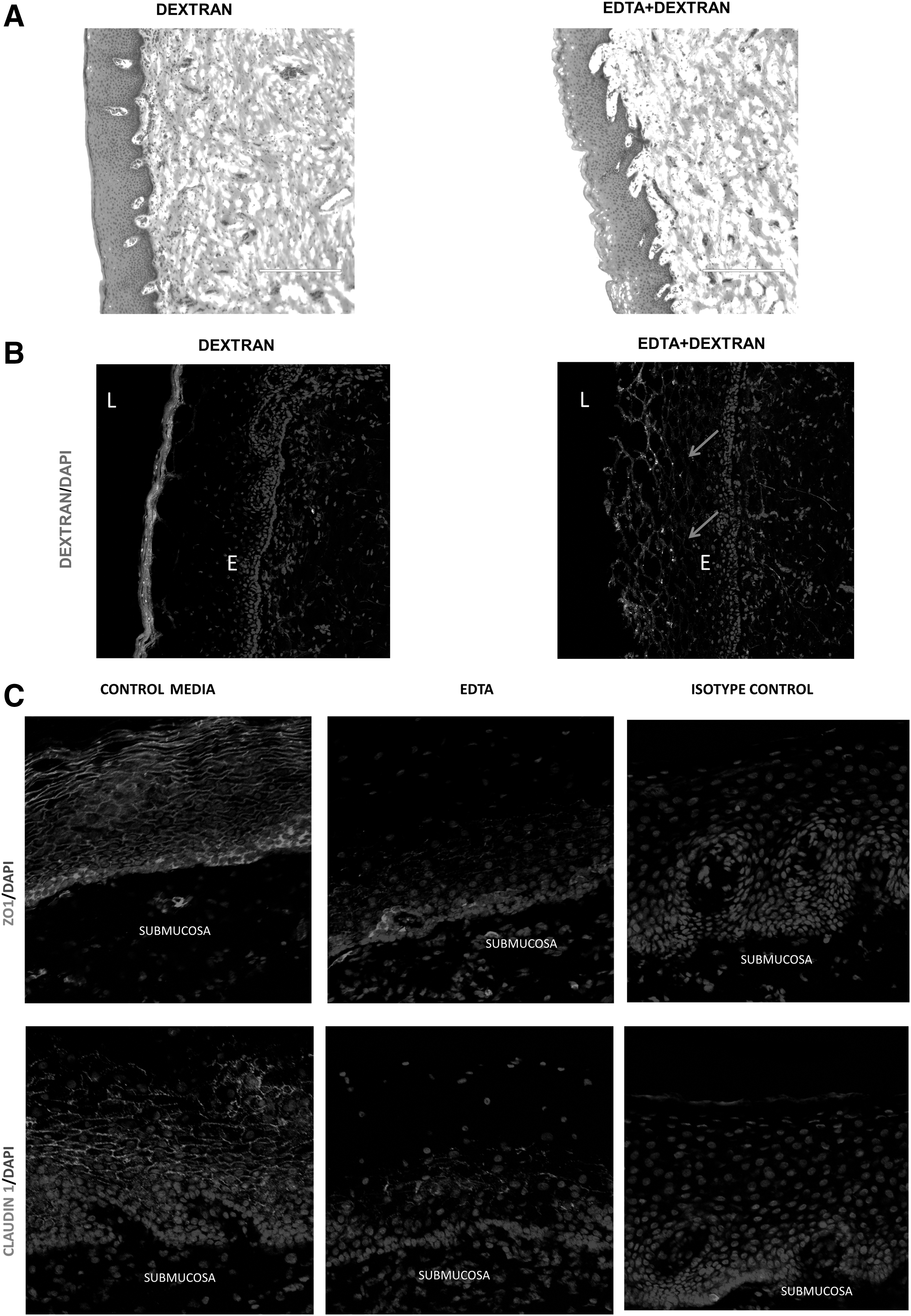
The intactness of apical cell membrane of epithelial cells and the intercellular TJ/AJ proteins forms the basis of barrier function of mucosal epithelia. We first performed a histological examination of the epithelial layer of the ectocervical and colon tissues to determine the integrity of epithelial layer after incubation with and without HIV in organ culture. In the ectocervical tissues, H&E staining showed that the epithelial lining of the ectocervical mucosa remained intact after cultivation for 24 h with and without HIV and was characterized by multilayered stratified squamous epithelial cells and basal layer (Fig. 1A).

Effect of HIV exposure on the integrity of the cervical/colon mucosal epithelia: H&E staining of
Histological examination of colon tissues showed retention of the epithelium and lamina propria cell integrity after 6 h in culture with and without HIV, which was similar to that of healthy colon tissue without cultivation (Fig. 1B). However, when colon explants were cultured for more than 10 h, shedding of the epithelial layer was detected (data not shown).
Characterization of TJ and AJ in ectocervical tissues
Epithelial layers in ectocervical and colonic tissues provide a robust barrier against microorganisms. Various studies have shown the existence of TJ and AJ between epithelial cells in the colon tissue, but very little information is available regarding the structure and molecular composition of ectocervical epithelial junctions between the cells. To determine whether TJ and AJ exist in the multilayer ectocervical epithelium, we examined the profiles of a number of TJ proteins (ZO-1, Claudin-1, Claudin-4), AJ protein (E-cadherin), and TJ gate function in ectocervical epithelium.
TJ regulates the passage of ions and small molecules through the paracellular pathway and serves as a permeability barrier (gate function). To evaluate the gate function, we analyzed the paracellular permeability of ectocervical epithelia to solutes (3 kDa—fluorescent-labeled dextran) in tissues by confocal microscopy. As shown in the Figure 2B, fluorescent-labeled dextran was not detected within the epithelial layer of the tissue, suggesting that the TJs of the ectocervical mucosal epithelium prevented the dextran penetration. In contrast, when ectocervical tissues were exposed to EDTA, an agent known to disrupt TJ/AJ, fluorescent-labeled dextran penetrated into the epithelial layers of the ectocervical tissue (Fig. 2B). Therefore, the epithelial junction in cervical tissue could “gate” the diffusion of dextran between epithelial cells.
Characterization of TJ and AJ in ectocervical tissues:
To study the TJ and AJ proteins, immunofluorescence staining of TJ and AJ proteins was performed on the ectocervical tissue cultured in media for 2 h. Immunofluorescence studies detected well-defined and interconnected ZO-1 and Claudin-1 expression pattern in the perimeter of each epithelial cell (Fig. 2C) in control tissues. However, in EDTA-treated ectocervical tissues, discontinuous/punctate distribution of ZO-1 and Claudin-1 was observed between epithelial cells (Fig. 2C), indicating disruption of TJ/AJ.
Effect of HIV on TJ proteins and AJ proteins in ectocervical/colon tissues
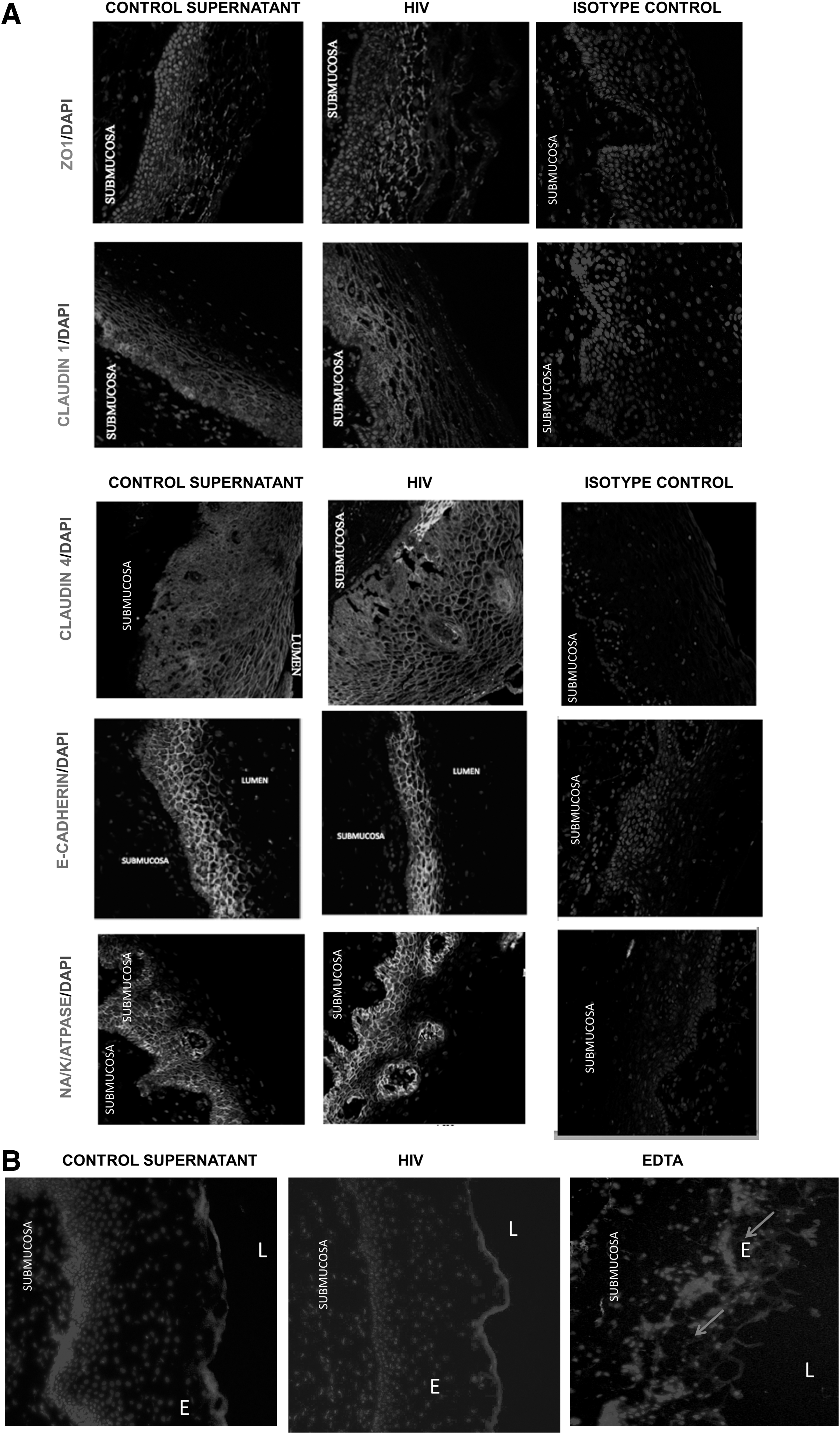
To investigate the effect of HIV exposure on the epithelial TJ/AJ proteins in ectocervical and colon tissues, the tissues were either exposed to HIV or control supernatant. Confocal microscopy was performed and images of mucosal epithelia were captured and quantitated. Control epithelia were characterized by well-defined and interconnected ZO-1, Claudin-1, and E-cadherin staining pattern located at the perimeter of each cell as a ring shape.
Based on the previous study on primary endometrial cells and intestinal cell lines that showed TJ disruption of epithelial cells postexposure to HIV, 10 we expected to notice the following characteristics of TJ/AJ proteins postexposure to HIV: discontinuous distribution pattern around the perimeter of cells, diffused cytoplasmic localization of TJ/AJ proteins, presence of TJ/AJ protein in the apical and basal cell layers, and reduced or complete inhibition of TJ/AJ protein expression. Contradictory to our expectation, epithelial layers of ectocervical/colon tissues exposed to HIV had similar distribution patterns of TJ proteins ZO-1, Claudin-1, Claudin-4, and E-cadherin compared with those in the control tissues (Figs. 3A and 5).
Effect of HIV exposure on TJ proteins and AJ proteins in ectocervical tissues.
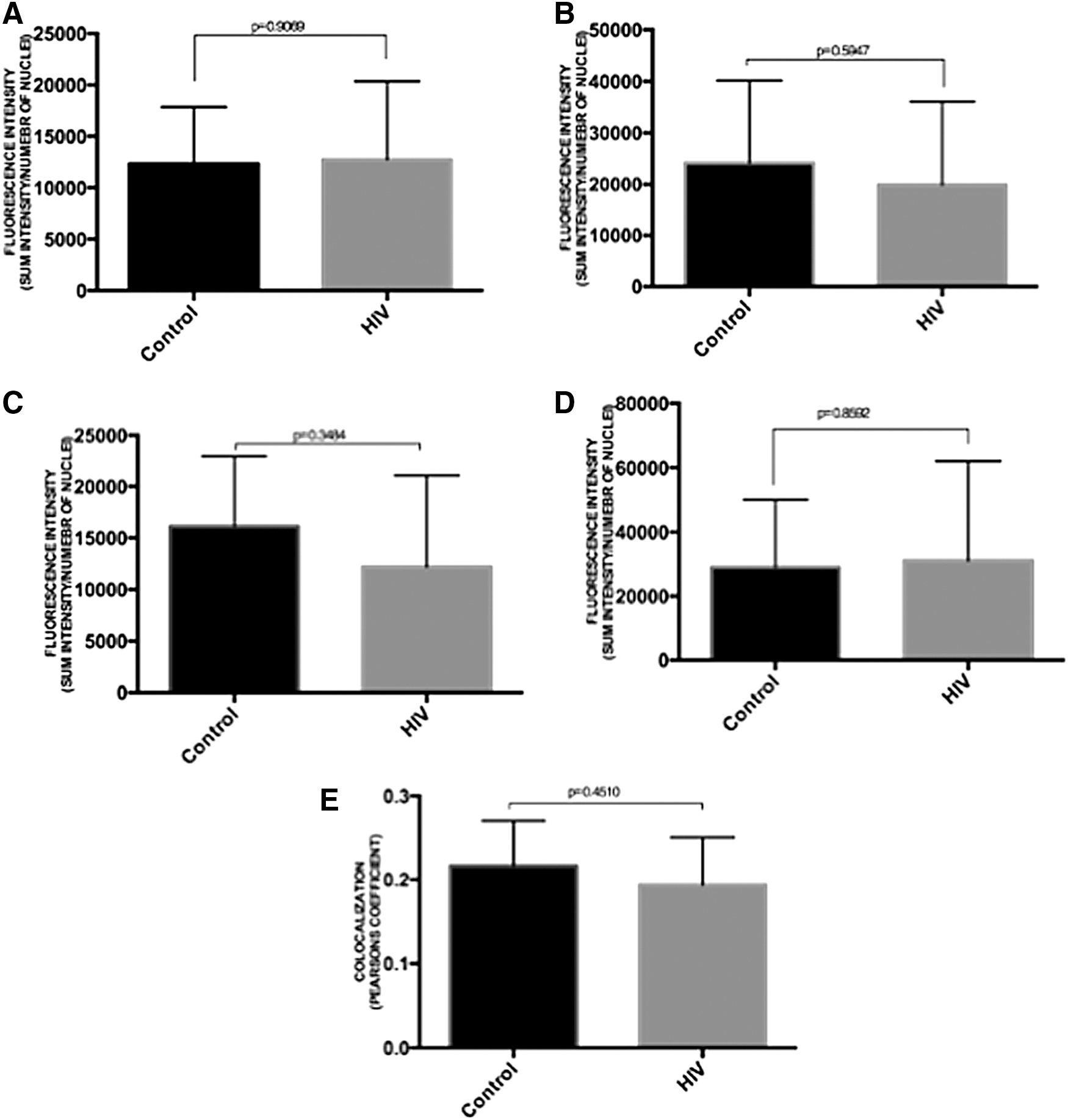
Quantitative analysis revealed that there was no significant reduction in fluorescent intensity of ZO-1, Claudin-4, and E-cadherin proteins in HIV-exposed tissues compared with control tissues, indicating that there was no marked reduction in TJ/AJ protein expression following exposure to HIV (Figs. 4 and 6). Furthermore, a number of studies suggest that ZO-1 nuclear translocation could be occurring when TJ between epithelial cells is disrupted following heat shock, chemical insults, or impaired cell–cell contacts. 21 Quantitative analysis of the presence of ZO-1 in nucleus also revealed that there was no significant increase in ZO-1 nuclear translocation in ectocervical epithelial cells after exposure to HIV (Fig. 4E).
Quantitation of TJ and AJ proteins in ectocervical tissues. Fluorescence intensity of ZO-1
To examine the barrier properties of the TJ/AJ after exposure to HIV, we analyzed the paracellular permeability of ectocervical epithelia to solutes (3 kDa—fluorescent-labeled dextran) in controls and HIV-exposed tissues by microscopy. As shown in the Figure 3B, fluorescent-labeled dextran was detected on the surface of the epithelial layer and not detected within the epithelial layer of the tissue in controls and HIV-exposed cervical tissues. In contrast, the fluorescent-labeled dextran was observed in basal membrane and subepithelial regions in ETDA-treated tissues. These results suggest that the TJs and barrier properties were not disrupted after exposure to HIV for 24 h. Thus, exposure to HIV did not disrupt the distribution of TJ proteins and AJ proteins between the epithelial cells in both colon and ectocervical tissues.
Effect of HIV on NA/K/ATPASE in ectocervical/colon tissues
TJ maintains cell polarity by restricting lateral diffusion of proteins and membrane lipids between the basolateral and apical compartments. Previous studies have suggested that Na/K/ATPase, which is located in the basolateral membrane of the cell, plays an important role in the maintenance of TJs. 9,22 Furthermore, distributions and expression of Na/K/ATPase were altered under pathogenic conditions. 9,23 –28 However, the effect of HIV exposure on the Na/K/ATPase distribution in mucosal epithelia is not known.
To study the effect, the distribution of Na/K/ATPase between apical and basolateral membrane of epithelial cells was assessed in control/HIV-exposed ectocervical/colon tissues. In the control ectocervical/colon tissues, Na/K/ATPase staining was most intense in the basolateral regions. However, such distribution of Na/K/ATPase was not altered after exposure to HIV (Figs. 3 and 5). Furthermore, quantitative analysis of the Na/K/ATPase expression also revealed that there was no significant difference in the levels of Na/K/ATPase expression between HIV-exposed and control tissues (Figs. 4 and 6). Thus, exposure to HIV did not significantly disrupt Na/K/ATPase profiles in the epithelial cells in both colon and ectocervical tissues.
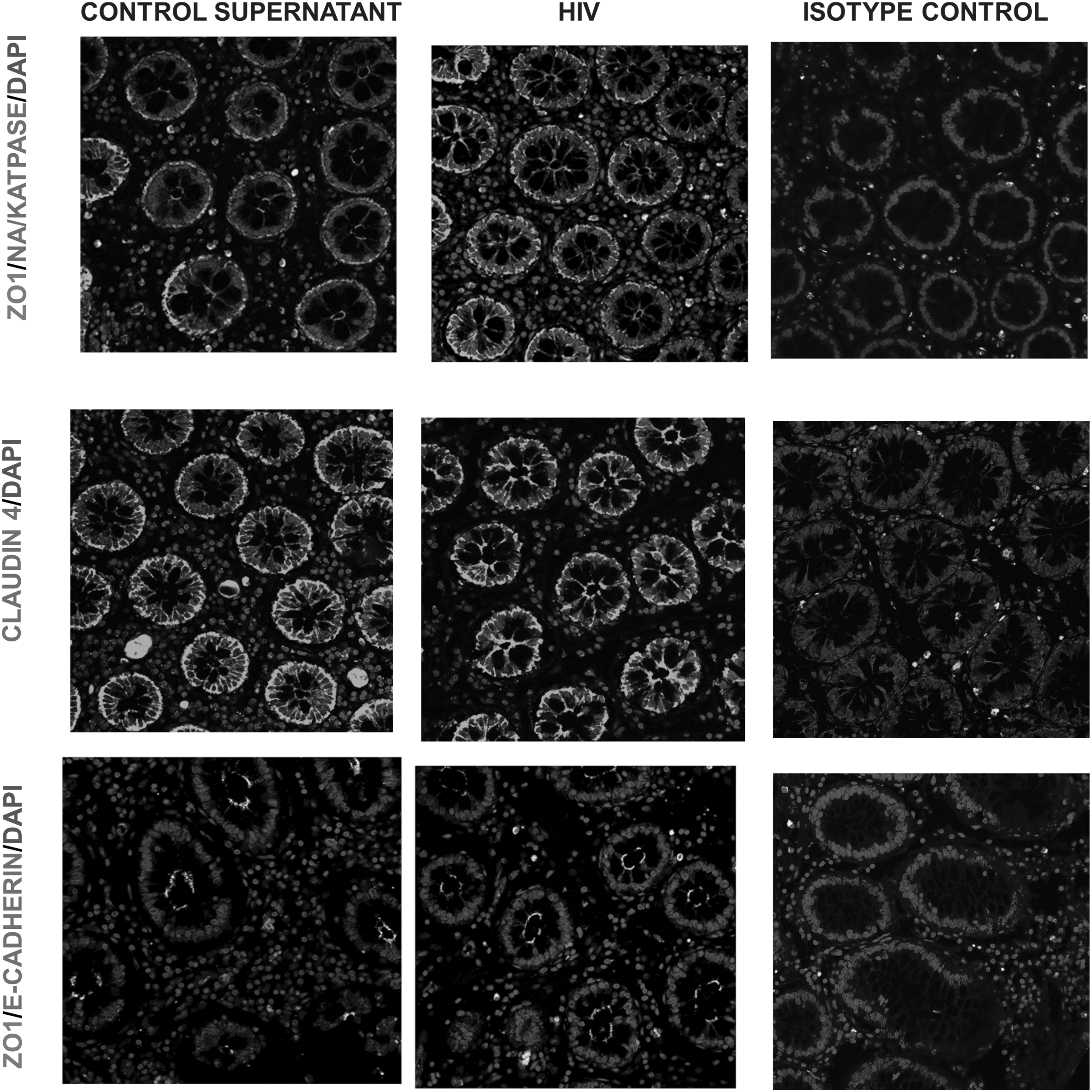
Effect of HIV exposure on TJ proteins and AJ proteins in colon tissues. Human colon tissues were either exposed to HIV (106 infectious viral units) or control supernatant for 6 h. Colon tissue sections were stained for ZO-1, Claudin-4, E-cadherin, or Na/K/ATPase protein. Images were captured by confocal microscope. Original magnification ×40. The images shown are representatives of four independent experiments with different donors. Each donor had two control and two HIV biopsies with 5–10 random images obtained from each biopsy.

Quantitation of TJ and AJ proteins in colon tissues. Fluorescence intensity of ZO-1
HIV transmission through ectocervical and colon tissues
Since we did not observe any effect of HIV exposure on TJ disruption, we investigated whether HIV transmission did occur within 6 h after exposure in colon and 24 h after exposure in ectocervical tissues. For this purpose, we examined entry of HIV into colon tissue within 6 h of HIV exposure by examining tissue using TEM. Following exposure of colonic tissues to HIV for 6 h, virus-like particles inside and between the epithelial cells were observed by TEM.
Morphologically, TEM revealed the presence of an epithelial layer with tightly packed epithelial cells and abundant microvilli in the colon luminal surface exposed to control supernatant or HIV (Fig. 7A, B). Furthermore, apical regions between the colon epithelial cells with and without HIV exposure were characterized by intact epithelial junctions, which help to prevent intercellular passage of luminal contents and maintains cohesion of the epithelium.
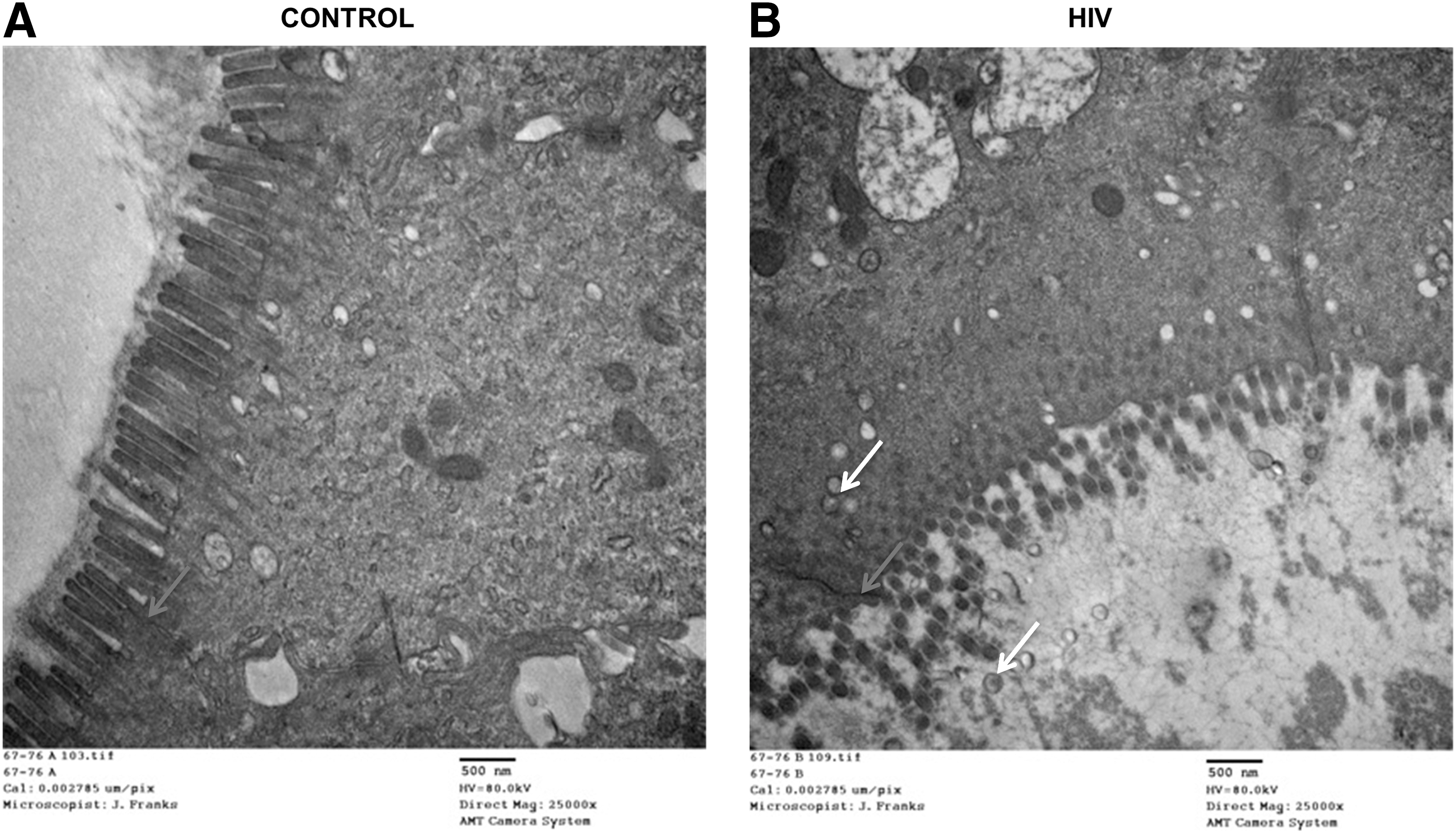
Visualization of epithelial junctions in the human colon tissues by transmission electron microscopy.
To further provide evidence that HIV did enter the ectocervical and colonic tissues within 24 and 6 h of infection, transmission of HIV in these tissues was examined. For this purpose, colon and ectocervical tissues were exposed to HIV for 6 or 24 h, respectively, after which the tissues were washed to remove viral inoculum and cultured in a fresh medium for 16 days. HIV transmission was detected by observing an increase in the level of HIV p24 in the culture supernatant of colon (Fig. 8A) and ectocervical (Fig. 8B) tissues. Viral replication became evident on day 4 postinfection and increased during the entire incubation period in both colon and ectocervical culture. At day 16, postinfection, the average p24 in culture supernatant was 1,160 pg/ml in colon tissue and 13,967 pg/ml in ectocervical tissue.
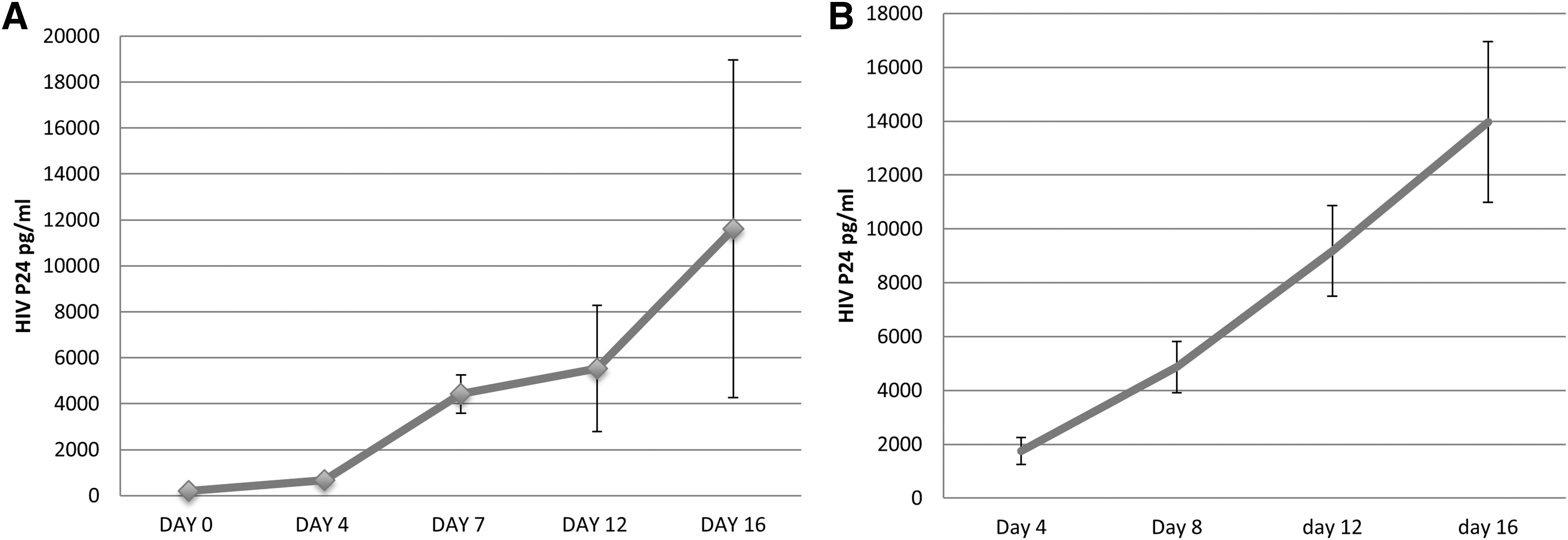
HIV replication in colon/ectocervical tissue after exposure to HIV in vitro.
Cytokine gene expression in epithelial cells following HIV exposure in the ectocervical and colon tissues
Previous study has reported that following HIV infection, there is an upregulation of TSLP that strongly activates human myeloid DC, leading to robust induction of homeostatic proliferation of CD4+ T cells and promoting HIV replication in the activated T cells. 4 Furthermore, studies have shown that accumulation of CD4+ T cells in human cervical tissue was detected within 6 h post-HIV infection. 17 Previous study has also reported an increased production of cytokines such as CXCL1 and CXCL10 in culture supernatant of HIV-exposed ectocervical tissues. 29 In addition, exposure to HIV has shown to induce production of TNF-α, IL-6, IL-8, MCP-1 in T84 intestinal epithelial cell lines and TNF-α, IL-6, MCP-1, IL-10, IL1β in primary endometrial epithelial cells. 10
We examined chemokine and cytokine levels in the epithelial cells of the tissues exposed to HIV since it may facilitate the establishment of local HIV infection after HIV crosses the epithelial barrier. For this purpose, the cytokine expression was analyzed in the ectocervical epithelia. To ensure the purity of microdissected ectocervical epithelia, mRNA level of cytokeratin 13 was measured, which is mainly expressed in epithelial cells and less expressed in submucosa. 29,30
CT values in real-time PCR measurement of cytokeratin was 10-fold less in epithelial regions compared with subepithelial region, which indicated that the epithelial cells were enriched at least by 1,000-fold in the microdissected epithelia compared with subepithelial regions (data not shown). Therefore, the epithelial cells seem to be the main source of observed chemokine expression in our study. As shown in Figure 9A, higher levels of CXCL10 and IL-6 mRNA expression were observed in human colon mucosa exposed to HIV compared with the controls. Similarly, higher levels of CXCL10 and CXCL11 mRNA were also detected in HIV-exposed ectocervical epithelia compared with the controls (Fig. 9B).

Cytokine gene expression in epithelial cells following HIV exposure in the ectocervical and colon tissues.
To determine if infectious HIV was required for such chemokine induction, ectocervical tissues were exposed to infectious HIV, AT-2-inactivated HIV, 30 or HIV-1gp120 followed by microdissection. As shown in Figure 9B, higher levels of CXCL10 and CXCL11 mRNA were only detected in ectocervical epithelia exposed to infectious HIV compared with the control. In contrast, exposure to AT-2-inactivated HIV or gp120 failed to increase these chemokine expressions in the ectocervical epithelia (Fig. 9C, D), indicating that HIV infectivity is required for chemokine induction in ectocervical epithelia.
Discussion
Mucosal surfaces are the primary sites for HIV infection during sexual transmission. In colonic and rectal mucosa, the epithelium is composed of a single layer of columnar cells, whereas ectocervical epithelium is composed of multilayered and stratified squamous epithelial cells, which play key roles in maintaining physical and functional barriers. 10 Although the epithelial layer lacks receptors for HIV entry, HIV can traverse through the epithelial layer and infect the underlying target cells.
Despite the importance of rectal and cervical mucosa in HIV transmission, current knowledge regarding HIV entry across mucosa is limited. 31 The purpose of the present study was to understand the mechanism of HIV transmission across ectocervical/colon mucosa using primary cervical and colonic tissues. Our organ culture model provides the natural tissue architecture, including epithelial layer, submucosa and the resident immune cells, including T cells, Langerhans cells, and macrophages, and allows for the evaluation of cellular factors that may be responsible for HIV transmission across the mucosa. 32
The current study utilizes our previously reported ectocervical tissue-based organ culture model 3 and colon organ culture model described by Dereuddre-Bosquet et al. 18 The existence of TJ in ectocervical epithelium remains controversial, and literature on characterization of epithelial intercellular junctions in human ectocervical tissue is limited. 5 In this study, we first established the existence of epithelial TJ and AJ junctions in normal human ectocervical tissues. The expression of TJ molecules such as ZO-1, Claudin-1, Claudin-4 and AJ molecule such as E-cadherin was detected in the stratified squamous epithelium of the ectocervix. Our results confirm and extend the observation of Blaskewicz et al. that TJ proteins ZO-1, Claudin-4, and AJ protein E-cadherin are indeed present in the ectocervical epithelium.
For colon/rectal tissues, the presence of TJ between the columial epithelial cells is well established. 33,34 Various pathogenic organisms have been shown to traverse through the epithelial barrier at the mucosal surface by disrupting the TJ, characterized by defective TJ barrier and increased intestinal permeability. 11,12 Recent studies have shown that HIV disrupted the epithelial TJ in an intestinal cell line and primary endometrium epithelial cells. 10,35 However, no studies have determined the direct effect of HIV on the epithelium in human ectocervical or colon tissues.
The present study evaluated the effect of HIV on TJ/AJ in cervical and rectal/colon epithelial layers in the context of tissue structure. H&E staining results suggest that the integrity and morphology of the epithelial layers on the cervical/colon tissues were not significantly changed by exposure to HIV in vitro. Furthermore, confocal microscopy of stained TJ and AJ suggests that the TJ and AJ structures in ectocervical/colon mucosal epithelia were not profoundly disrupted by exposure to HIV. Furthermore, we examined HIV transmission in the ectocervical and colon tissues. We observed HIV transmission in ectocervical and colon tissue as early as 24 and 6 h, respectively, following exposure to HIV. This confirms our previous results showing HIV-1 RNA-positive cells in the epithelial–submucosal junction at 6 h postexposure. 17
Our results also agree with previous reports from Dezzutti et al. showing HIV replication in colon tissues after exposure to HIV. 36 Studies using nonhuman primates also suggest that rectal entry of SIV is a very rapid process, and SIV was detected at distal sites very early postinfection. 37 These results indicate that HIV transverses into ectocervical/colon tissue by mechanisms other than TJ/AJ disruption in ectocervical/colon epithelia. Our results go along with findings of other studies that TJ disruption does not occur in colon tissues of humans 38 or rhesus macaques 15 at acute stage of infection.
In response to HIV infection, genital epithelial cells release TSLP that strongly activates human myeloid DC, which leads to robust induction of homeostatic proliferation of CD4+ T cells and promotes HIV replication in these activated T cells. 4 Furthermore, previous studies have shown accumulation of CD4+ T cells within 6 h post-HIV infection in human cervical tissue. 3,17 This suggests that an innate inflammatory reaction of epithelial cells may facilitate downstream amplification of virus in CD4+ T cells at the intraepithelial and submucosal layers. Previous study has also reported an increased production of cytokines such as CXCL1 and CXCL10 in culture supernatant of HIV-exposed ectocervical tissues. 39 However, the cellular source of inflammatory cytokines in HIV-infected patients remains unknown.
To investigate the cytokine/chemokine induction in mucosal epithelia postexposure to HIV, we analyzed the cytokine/chemokine mRNA expression in microdissected mucosal epithelia of the control and HIV-exposed ectocervical/colon tissues, hypothesizing that epithelial cells are the primary source of the chemokines postexposure to HIV. Our study demonstrated that CXCL10 and CXCL11 mRNA expression was significantly higher in the ectocervical epithelia exposed to HIV compared with those exposed to control supernatant and induction of such chemokines required infectious HIV.
Using different postviral infection cells, similar or higher mRNA levels of chemokine/cytokines were reported as fold change in virus-infected cells compared with controls. 40 In case of the colon mucosal layer, higher expression of CXCL10 and IL-6 was induced after exposure to HIV for 6 h. Similar results were observed in a previous study where the level of cytokines/chemokines was increased in intestinal cell lines postexposure to HIV. 10
Thus, results from our study suggest that the epithelial cells might be one of the primary sources of chemokines postexposure to HIV. Since CXCL10 and CXCL11 have been reported to contribute to the recruitment of susceptible T cell, macrophages, and DCs to HIV-infected lymph nodes, 41 the observed HIV-mediated upregulation of chemokines may be responsible for recruitment of target cells toward the epithelial region, resulting in enhanced HIV transmission.
Previous studies have suggested several pathways through which HIV can cross the epithelium, including transcytosis, transmigration, and using unknown receptors for HIV entry. 1,2,42,43 HIV-infected macrophages were shown to transmigrate via fetal oral mucosa 44 without disrupting the epithelial TJs. The results from the present study suggest that HIV transmission through the ectocervical and colon mucosa may occur via unidentified mechanism without TJ/AJ disruption. The presence of sexually transmitted infections (herpes), intravaginal products (some microbicides), vaginal practices (douching), and change in the genital tract microbial environment may cause TJ disruption but not by HIV itself. Further investigation is needed to determine the possible mechanism of HIV transmission via ectocervical and colon mucosa.
In summary, our results show that exposure to HIV does not disrupt TJs and AJs in ectocervical and colonic epithelium. However, HIV exposure at these mucosal surfaces results in chemokine production, which is dependent on HIV infectivity and may play a major role in instigating immune cell recruitment toward the epithelial layer, in turn facilitating HIV infection.
Footnotes
Acknowledgments
We thank Dr. Carolyn Coyne and Dr. Todd Reinhart for useful discussions and Dr. Rajiv Dhir for procuring ectocervical, colon tissues through the Pathology Tissue Bank of the University of Pittsburgh Medical Center. We thank Lori Caruso for editing the article. This project was supported by an NIH R01 grant HD072693 (P.G.), 1 K24HD080539 and 5RO1HD072693 (B.R.), Lifespan/Tufts/Brown Centre for AIDS research 5P30AI042853 (S.C-U., B.R.). S.S. and A.S. were supported by the Fogarty International HIV Research Training Program of the National Institutes of Health (D43TW001038 and D34TW010039).
Author Disclosure Statement
No competing financial interests exist.