
Abstract
MicroRNAs (miRNAs) participate in host innate immunity against HIV-1 infection. We examined the impact of HIV-1 infection on viral restriction miRNAs in plasma of HIV-1–infected subjects. HIV-1–infected subjects had significantly lower plasma levels of HIV-1 restriction miRNAs (miRs-29a, −29b, −125b, −223, −198, and −382) than control subjects. Further in vitro studies showed that HIV-1 infection of macrophages suppressed production of the extracellular miRs-29b, −125b, and −223. These data demonstrate the compelling evidence that HIV-1 infection impairs host innate immunity by inhibiting antiviral miRNAs, which provide a possible mechanism for HIV-1 persistence in the host.
M
Depending on locations and presence, miRNAs can be generally classified as cellular or circulating miRNAs. Accumulating evidence indicates the presence of cell-free miRNAs that are secreted by the cells to circulation in vivo. 8 A number of studies demonstrated that specific miRNAs could be secreted by various tissues, such as the heart, adipose tissue, liver, and aorta, and released into circulation. 9 These circulating miRNAs have been identified in a broad range of human body fluids, such as serum, plasma, saliva, tears, breast milk, and urine, and are believed to play a crucial role in mediating cell-to-cell communications. 9 A recent study demonstrated that miRNAs packed in cancer cell–secreted lipid vesicles can be transferred to surrounding immune cells and bind to Toll-like receptors (TLR7 in mice and TLR8 in humans) and thus to induce cytokine secretion and prometastatic inflammatory responses in immune cells. 8 Moreover, due to their high stability and ease of measurement, use of circulating miRNAs as potentially useful markers to monitor host immune and disease status has been extensively investigated for a variety of human diseases, such as cancer and viral infections. 10,11 Despite several previous studies that investigated the dynamics of the cellular miRNAs in HIV-1 disease, there is little information regarding the impact of HIV-1 infection on plasma circulating miRNAs. 12 We thus examined several key HIV-1 restriction miRNAs in the plasma of HIV-1–infected individuals and control subjects.
HIV-1–infected individuals and uninfected healthy subjects were recruited through the clinic at the Wuhan Centers for Disease Prevention and Control (WHCDC) in China. Informed consent was obtained from the study subjects. Institutional review boards of the WHCDC and Temple University approved this study. Subjects were excluded if they had a chronic systemic illness (cardiac, renal, pulmonary, hepatic, endocrine, metabolic, or autoimmune disorders) or other medical disorders. Demographic and clinical characteristics of these study subjects are summarized in Table 1. Whole blood was collected from study subjects using EDTA as anticoagulant. Separation of plasma was accomplished by centrifugation at 2000 rpm for 15 min. The plasma was stored at −80°C until use. Total RNA was isolated from 200 μl plasma using miRNeasy Mini Kit (QIAGEN, Valencia, CA). Synthetic Cel-miR-39 was added to the samples as an internal control according to the instruction of manufacturer. RNA (1 μg) was reverse transcribed using miScript Reverse Transcription Kit (QIAGEN). Real-time polymerase chain reaction (PCR) for the quantification of a subset of miRNAs (miRs-29a, −29b, −198, −28, −125b, −150, −223, and −382) was carried out using miScript Primer Assays and miScript SYBR Green PCR Kit (QIAGEN). Among the eight miRNAs tested, the levels of miRs-125b, −223, −382, −198, −29a, and −29b were significantly lower in HIV-1–infected subjects than in the control subjects (Fig. 1A–F). In contrast, no significant differences for miRs-28 and −150 were observed between the HIV-1–infected individuals and control subjects (Fig. 1G–H).

Effect of HIV-1 infection on circulating HIV-1 restriction miRNA expression. Plasma specimens were collected from control and HIV-1–infected subjects. Total RNA was extracted from plasma and subjected to the real-time reverse transcriptase–polymerase chain reaction (RT-PCR) to quantify circulating miRNA-125b
To validate these in vivo observations, we examined the in vitro effect of HIV-1 infection on the expression of macrophage-derived extracellular miRNAs. Macrophages are a major player in anti–HIV-1 immunity, while they also serve as a major target of HIV-1 infection. Macrophages express a number of the HIV-1 restriction miRNAs. 13 Purified human monocytes obtained from the Human Immunology Core at the University of Pennsylvania were plated on 48-well plates (106 cells/well) in complete Dulbecco's modified Eagle's medium (DMEM) with 10% fetal calf serum. Monocytes were differentiated into macrophages during the in vitro cultures (5–7 days). HIV-1 R5 Bal strain was obtained from the AIDS Research and Reference Reagent Program (National Institutes of Health, Bethesda, MD). Macrophages were infected with cell-free HIV-1 Bal strain for 2 h at 37°C. The cells were then washed three times with DMEM to remove extracellular free viruses, and fresh medium was added to the culture. The final wash was tested for HIV-1 reverse transcriptase (RT) activity and shown to be free of residual viruses. The cells were incubated for 12 days, and culture supernatant was harvested for HIV-1 RT activity assay to confirm the viral infection and for miRNA extraction. As shown in Figure 2, three HIV-1 restriction miRNAs (miRs-125b, −223, and −29b) were significantly lower in supernatant of HIV-1–infected macrophages than in uninfected cell cultures.
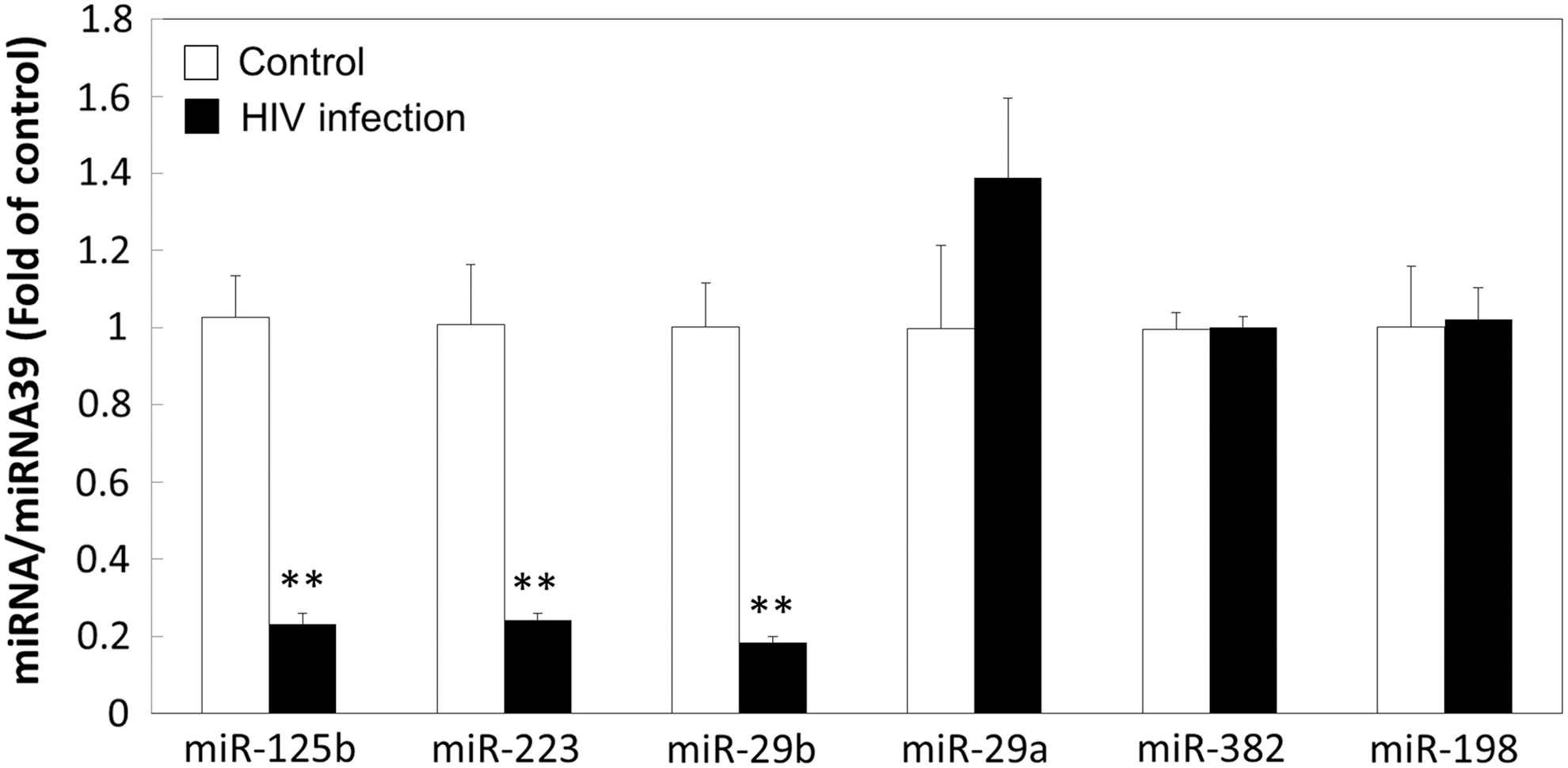
Effect of HIV-1 infection on cellular HIV-1 restriction miRNA expression by macrophage. Monocytes were isolated from peripheral blood mononuclear cells (PBMCs) of healthy subjects and cultured 7 days for differentiation into macrophages. Macrophages were then infected by HIV-1 Bal, and noninfected macrophages were included as a control. Fresh media were added into culture at day 9 postinfection, and supernatant was collected 3 days later for miRNA extraction. RNAs were subjected to the real-time RT-PCR for the levels of miRs-125b, −29b, −223, −29a, −382, and −198. Cel-miR-39 was used as a spiked-in control. The data are expressed as the miRNA levels relative (fold) to the control (without HIV-1 infection, which is defined as 1). The results shown are the mean ± SD of triplicate wells, representing three independent experiments (**p < .01).
In the present study, we selected eight well-documented miRNAs (miRs-28, −125b, −150, −223, −382, −29a, −29b, and −198) that have the ability to inhibit HIV-1 replication by targeting either viral genome or cellular factors. We found that six of these HIV-1 restriction miRNAs (miRs-125b, −223, −382, −29a, −29b, and −198) were significantly decreased in plasma from HIV-1–infected subjects. Consistent with the in vivo observations, our in vitro experiments showed that HIV-1 infection of macrophages inhibits the production of extracellular miRs-29b, −125b, and −223. Houzet et al. 11 demonstrated that of the 63 miRNAs that were perturbed by HIV-1 infection, 59 miRNAs were decreased in peripheral blood mononuclear cells (PBMCs) of HIV-1–infected individuals compared to those of uninfected healthy donors. In particular, they found that expression of miRs-29a and −29b was significantly downregulated in the PBMCs of HIV-1–infected subjects, and several T-cell–specific miRNAs, including miR-223, were decreased in HIV-1–seropositive individuals. 11 Additionally, Witwer et al. 14 examined expression of cellular miR-125b and miR-29 family in PBMCs of HIV-1 elite controllers or suppressors (ES) as well as of viremic patients and found that both miR-125b and the members of miR-29 family (miRs-29a, −29b, and −29c) were downregulated in these subjects.
The mechanisms of HIV-1 infection–mediated downregulation of the viral restriction miRNAs remain to be determined. It was reported that HIV-1 Tat protein could inhibit several key proteins (Dicer and Drosha), which are essential for miRNA biogenesis. 6 Triboulet et al. 15 demonstrated the knockdown of Dicer- and Drosha-enhanced HIV-1 infection of T cells, implying the potential role of these proteins in modulating HIV-1–related miRNAs. However, it is unclear why some of the HIV-1 restriction miRNAs were not affected by HIV-1 infection. In our in vitro experiments, only three of the six restriction miRNAs (miRs-125b, −223, and −29b) were inhibited by HIV-1 infection of macrophages (Fig. 2). In addition, we observed that the plasma levels of miRs-28 and −150 were not affected in HIV-1–infected patients (Fig. 1G, H). A recent study by Munshi et al. 16 demonstrated that the levels of circulating miR-150 were increased in plasma of HIV-1–infected subjects, even though its cellular levels in PBMCs were decreased by HIV-1 infection. These findings indicate that the levels of miRNAs are differentially regulated in different immune compartments.
In summary, we present the compelling in vivo and in vitro evidence that HIV-1 infection could inhibit the plasma and extracellular levels of the viral restriction miRNAs. These findings highlight the importance to further investigate the role of circulating miRNAs in the immunopathogenesis of HIV-1 disease. Understanding whether HIV-1 infection modulates the host miRNAs will not only shed lights on the mechanisms of HIV-1 persistence in the host but also reveal the likelihood that HIV-1 restriction miRNAs can be used as useful markers to monitor immune status and disease progression during HIV-1 infection.
Footnotes
Acknowledgments
This study was supported by the National Institutes of Health (DA012815 and DA022177 to W.Z.H.) and in part by the National Natural Science Foundation of China (81271334 to W.Z.H. and 81301428 to L.Z.).
Author Disclosure Statement
No competing financial interests exist.