
Abstract
Contradictory results have been reported on the impact of duplications/insertions in the HIV-1 gag-p6 late assembly domains [TSG101-binding P(T/S)APP motif and ALIX-binding LYPxnLxxL motif] heterogeneity following therapy failure. However, most studies are limited to small numbers of patients and do not include samples from South Africa, which has the largest number of HIV-1C-infected patients (HIV-1CZA). In this study we compared the gag-p6 variability among HIV-1CZA-infected patients from a South African clinical cohort who experienced antiretroviral therapy (ART) failure (n = 845) with ART-naive HIV-1CZA sequences (n = 706) downloaded from the Los Alamos database. Partial (PTA/PTV/APP) or complete P(T/S)APP duplications were less frequent in HIV-1CZA with ART failure compared to therapy-naive ones (14% vs. 30%; p < 0.001). In contrast, the tetrapeptide PYxE insertion, recently described by us, occurred more frequently (5-fold) in therapy-failure patients (p < 0.001) and was associated with a higher number of reverse transcriptase inhibitor (RTI) mutations (p = 0.04) among patients failing ART.
H
We recently reported intra-HIV-1C-specific differences and a high rate of P(T/S)APP duplications in Indian HIV-1CIN-infected patients failing ART, but not in an HIV-1C-infected German cohort. 11 We also identified a novel tetrapeptide PYxE insertion in the ALIX-binding motif that is highly specific for the HIV-1C strains from Ethiopia (HIV-1CET) with a prevalence in half of the HIV-1CET strains. 11 This insertion is rare (∼1%) in HIV-1C strains from Southern Africa (South Africa, Botswana, Zambia, Malawi, etc.) and India (HIV-1CIN) and is extremely rare in non-C subtypes (∼0.1%). The PYxE insertion was also associated with PI failure in HIV-1CIN strains. However, these studies, including ours, were limited by small numbers of patients and did not include South African HIV-1C sequences (HIV-1CZA), the country with the highest number of HIV-1C infections globally.
Therefore, the main objective of this study was to describe the gag-p6 variability among HIV-1CZA-infected patients from five provinces of South Africa who were enrolled in the national ART program and experienced therapy failure. We compared the data obtained from therapy-naive HIV-1CZA sequences downloaded from the Los Alamos database. In addition, the profile of drug resistance mutations (DRMs) among the failure patients was described as well as their association with duplications in the TSG101-binding P(T/S)APP motif and insertions in the ALIX-binding LYPxnLxxL motif. This is the largest study analyzing the variability of gag-p6 and DRMs so far in HIV-1C and may have implications for the global clinical management of ART patients.
The patients (n = 1026) included were treated as part of the national ART program in South Africa and had samples submitted for genotypic HIV drug resistance testing following ART failure. Samples were received between January and December 2013 from five provinces. The majority of the samples are from Gauteng province (53%; 538/1026), followed by Eastern Cape (33%; 339/1026), Western Cape (9%; 97/1026), KwaZulu-Natal (3%. 33/1026), and Free State (2%; 19/1026) (Fig. 1). Gag-p6 sequences were downloaded (accessed on January 7, 2015) from the Los Alamos database using the “therapy-naive” filter. After excluding multiple sequences from a patient, frameshifts and stop codons, a total of 706 Gag-p6 sequences was included for further analysis.

Distribution of HIV-1C samples used in this study in South Africa.
Genotypic resistance testing (GRT) was performed using a previously described in-house method that provides information for full-length protease (PR: 1–99 amino acids) and partial reverse transcriptase (RT: 1–250 amino acids). 12 Subtyping was performed by COMET-HIV 13 and REGA v3. 14 Acquired DRMs were evaluated using the International AIDS Society list, update 2014, 15 and Stanford HIVDB v7.0.1 (accessed on January 13, 2015). 16 The level of resistance was categorized into four groups based on the cumulative resistance mutation scores (RMS) as susceptible (RMS: <10), low-level (RMS: 10–25), intermediate (RMS: 26–55), and high-level resistance (>55). Mann–Whitney U and χ2 tests were performed when necessary. Statistical analysis was performed in SPSS v22 (IBM, USA). p < 0.05 was considered significant.
This study was approved by the Health Research Ethics Committee (HREC) of Stellenbosch University (N11/09/274) with a waiver of patient consent. The data were made anonymous: patients' identities were removed before any analysis. The HREC complies with the South Africa National Health Act No. 612003 and the United States code of Federal Regulations title 45 Part 46. The committee also abides by the ethical norms and principles for research as established by the Declaration of Helsinki, the South African Medical Research Council Guidelines, as well as the Department of Health Guidelines.
All patients (n = 1026) were infected with HIV-1CZA and they represented a South African cohort of ART-failure patients. The median age of the patients was 35 years (range 1–68) and 61% were females (628/1026). Gag-p6, which contained both the TSG101-binding motif and the ALIX-binding motif, was successfully extracted in 845/1026 (83%) of the patients. Partial (PTA/PTV/APP) or complete P(T/S)APP duplications (3–19 amino acid long duplications) in the TSG101-binding motif were observed in 14% (116/845) of the therapy failure patients, which was less than in the therapy-naive patients' HIV-1CZA sequences downloaded from Los Alamos (30%; p < 0.001) (Fig. 2A). However, the tetrapeptide PYxE insertion was observed at a significantly higher frequency in ART-failure patients than in therapy-naive patients (5% vs. 1%; p < 0.001) (Fig. 2B).
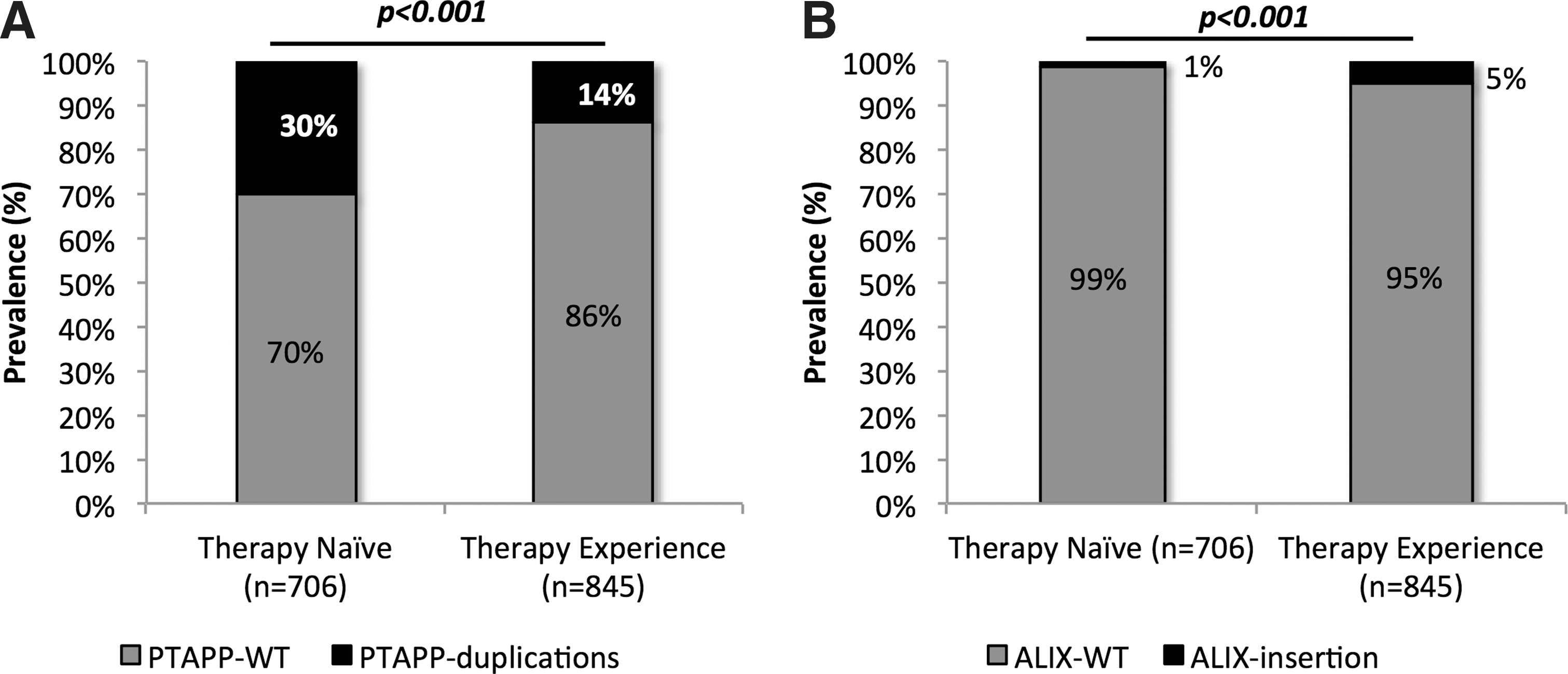
Gag-p6 mutations among the antiretroviral therapy (ART)-failure patients in the South African clinical cohort and therapy-naive HIV-1CZA patients' sequences downloaded from the Los Alamos database.
The sequence logo of the unique PYxE motif is presented in Fig. 3. The association of P(T/S)APP duplications and response to ART is debatable. Studies from Switzerland (HIV-1B), Brazil (HIV-1CBR), and India (HIV-1CIN) identified more duplication among ART-experienced than ART-naive patients 9,17 in contrast to two other studies. 6,11 Also, these mutations have been associated with decreased viral responses among highly treatment-experienced patients in one study, 7 which was not confirmed in a larger study of 461 patients from Canada. 5

Sequence logo of the ALIX-binding motif. The sequence logo was created with the sequences that had PYxE insertion (n = 42) from the South African therapy-failure cohort.
Based on our earlier large study of HIV-1C, a subtype that is more prone to duplication/insertions in Gag-p6, we hypothesized that the viral genetic background could influence the preferential selection of duplications and insertions in Gag-p6. 11 However, in our current cohort of HIV-1CZA we observed a significantly decreased prevalence of P(T/S)APP duplication in ART-failure patients compared to ART-naive ones. Combining the results of our study with others, we therefore postulate that the P(T/S)APP duplication in HIV-1C might be occurring naturally and may not have any influence on therapy. 5
We further analyzed the therapy response in HIV-1C-infected patients (n = 128) with a minimum duration of 2 years of therapy from the Swedish InfCare cohort. No association was observed with primary (viral load >50 copies/ml to 9 months of therapy) or secondary (single viral load >500 copies/ml or two consecutive viral loads >50 copies/ml) viral failure with the presence of PTAP duplications before therapy (U. Neogi, unpublished observations). However, its role in viral fitness and pathogenicity requires further investigation.
In contrast, the novel PYxE insertion, earlier identified by us, 11 in the C-terminal region of Gag-p6 could have a fitness compensatory role in highly treated individuals who have several DRMs in the RT and/or PR. Unlike HIV-1CET strains in which we observed PYRE, PYKE, and PYQE, the HIV-1CZA strains contain only PYRE and PYKE insertions. In our earlier study we observed that the PYxE insertion was associated with PI failure among the Indian HIV-1CIN patients. In the current study we observed a 5-fold increase in the occurrence of the PYxE insertion among South African patients failing ART.
The drug resistance pattern of 845 patients who had failed ART is presented in Table 1. Thymidine analogue mutations (TAMs) were identified in 22% of the patients. The TAM-2 pathway (T215F, D67N, K70R/E, and K219E/Q) was observed more frequently than the TAM-1 pathway (T215Y, M41L, and L210W) (18% vs. 8%; p < 0.05). The TAM mixed pathway was observed in 4% of the patients. The non-TAM mutations M184V/I and K65R occurred in 64% and 21% of the sequences, respectively. Of the NNRTI mutations, the most common mutations observed were K103N (43%), V106M (29%), G190A (16%), and Y181C (12%). Major PI resistance mutations were observed in 5% of the patients; among them V82A (65%; 28/43), I54V (63%; 27/43), L76V (23%; 10/43), and L90M (16%; 7/43) were the most frequent.
NRTI, nucleoside reverse transcriptase inhibitors; TAM, thymidine analogue mutation; NNRTI, nonnucleoside reverse transcriptase inhibitor; RTI, reverse transcriptase inhibitor.
The level of resistance based on the cumulative RMS is presented in Fig. 4. The highest proportion of susceptibility was toward zidovudine while 40% of the patients showed a certain degree of resistance to tenofovir. As expected, 75% of the patients had a high level of resistance to nevirapine and efavirenz. Various degrees of cross-resistance to second generation NNRTIs, etravirine and rilpivirine, was observed in 66% of the patients.
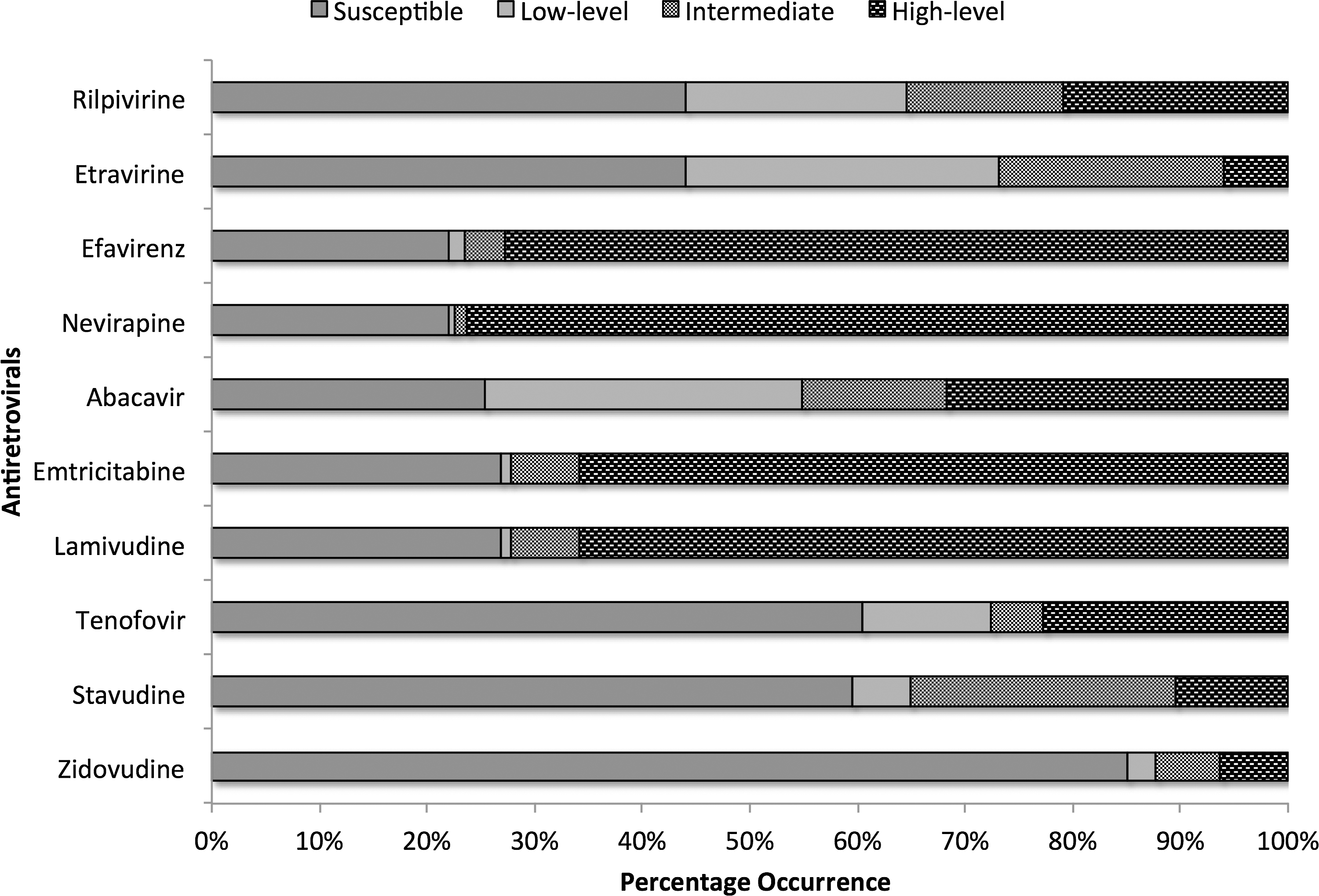
The level of resistance based on the cumulative resistance mutation scores (RMS). Susceptible (RMS: <10), low-level resistance (RMS: 10–25), intermediate-level resistance (RMS: 26–55), and high-level resistance (>55).
Due to nonavailability of drug history and duration of treatment, we categorized the patients into heavily ART exposed, defined as those having ≥4 RTI mutations and/or ≥2 major PI mutations; nonadherent to treatment (no DRM detected); and early failure [only a few mutations (≤3 DRMs and no PI mutation) with single signature mutations such as M41L, K70R, M184V, and T215Y/F]. No association of P(T/S)APP duplications was observed with the number or type of drug resistance mutations. In contrast, in the heavily ART-exposed individuals (n = 427) the frequency of the PYxE insertion in the ALIX-binding motif was significantly higher than in the other two (early failure or noncompliance) categories (6% vs. 3%; p = 0.03). The number of RTI mutations was also significantly associated with the ALIX insertion (p = 0.04, Mann–Whitney test). A large number of RTI-DRM significantly associated with the PYxE insertion could indicate that it has a compensatory viral fitness effect. However, it would require functional studies to substantiate this. Due to the small number of patients with PI mutations we did not find any association with only the PI-DRM and PYRE insertion. Reanalysis of our earlier published data 11 confirms that all six HIV-1CIN sequences that had the PYxE insertion were obtained from patients who had failed both RTI-based first-line ART and a second-line PI-based ART, exhibiting extensive RTI-DRMs and/or PI-DRMs (≥6).
The study had the following limitations and strengths: First, the lack of patients' clinical and drug history. As the patient samples were part of the South African National ART program, the samples were often sent as a result of virological failure without providing additional clinical or ART history. Second, this is a cross-sectional study, which limits the analysis of the dynamics of duplication and insertion. Nevertheless, the major strength of the study is the comparatively large number of samples analyzed, which limits the sampling bias. Moreover, this study represents patient samples from five provinces of South Africa, which provides an excellent base of comparison for the database sequences in which the majority of the sequences are from these regions.
In conclusion, using the data from therapy-failure patients enrolled in the South African national ART program representing therapy-naive sequences from a secondary database, we determined that partial or complete P(T/S)APP duplication in the TSG101-binding site was significantly less common in HIV-1CZA sequences from patients with therapy failure and did not have any association with drug resistance mutations. We therefore postulate that the P(T/S)APP duplication in HIV-1CZA is naturally occurring and might not have any effect on therapy response. 18 This is further supported by the patients' follow-up data from the Swedish cohort. As the P(T/S)APP motif is a target for host cellular factors it might be associated with host immune escape or be related to viral fitness. 5,6,18 In contrast, the tetrapeptide insertion identified by us 11 might have a role in the changes in viral fitness associated with the evolution of DRMs due to failure. Further longitudinal studies are required to identify the role of the PYxE insertion in the ALIX-binding motif in highly treatment-experienced patients and its association with the therapeutic response to salvage therapy.
Footnotes
Acknowledgments
The study was partially funded by Karolinska Institutet Research Foundation Grants (2014fobi41250) to U.N. and grants from the Swedish Research Council to A.S., the National Research Foundation (NRF) of South Africa (G.J. and S.E.), the Medical Research Council (MRC) (S.E.), and the Poliomyelitis Research Foundation (PRF) (S.E.). All the sequences have been submitted to GenBank under the following accession numbers: KT592705–KT593549.
Ujjwal Neogi and Susan Engelbrecht contributed equally to this work.
Author Disclosure Statement
No competing financial interests exist.