
Abstract
Vaccines based on live attenuated viruses are highly effective immunogens in the simian immunodeficiency virus (SIV)/rhesus macaque animal model and offer the possibility of studying correlates of protection against infection with virulent virus. We utilized a tether system for studying, in naive macaques and animals vaccinated with a live-attenuated vaccine, the acute events after challenge with pathogenic SIV. This approach allowed for the frequent sampling of small blood volumes without sedation or restraining of the animals, thus reducing the confounding effect of sampling stress. Before challenge, vaccinated animals presented significantly higher levels of proliferating and activated B cells than naive macaques, which were manifested by high expression of CD8 on B cells. After SIV challenge, the only changes observed in protected vaccinated macaques were significant increases in expression of the NK marker NKG2C on CD4 and CD8 T cells. We also identified that infection of naive macaques with SIV resulted in a transient peak of expression of CD20 on CD8 T cells and a constant rise in the number of B cells expressing CD8. Finally, analysis of a larger cohort of vaccinated animals identified that, even when circulating levels of vaccine virus are below the limit of detection, live attenuated vaccines induce systemic increases of IP-10 and perforin. These studies indicate that components of both the innate and adaptive immune systems of animals inoculated with a live-attenuated SIV vaccine respond to and control infection with virulent virus. Persistence of the vaccine virus in tissues may explain the elevated cytokine and B-cell activation levels. In addition, our report underpins the utility of the tether system for the intensive study of acute immune responses to viral infections.
Introduction
I
One of the most widely utilized, live attenuated SIV vaccines is the SIVmac239Δnef vaccine. Using this live attenuated vaccine in rhesus macaques with a subsequent challenge with pathogenic SIV has resulted in the identification of many factors that are associated with protection. These factors include anentropic specificity and repertoire depth of CD8 T cells, 12 localized innate immune responses, 13 inhibition of CD4 T-cell recruitment, 14 CD8 T-cell-related transcription factor profiles, 15,16 env-specific antibody-dependent cell-mediated cytotoxicity, 17 antibodies, 18 –20 the location, effector functions and phenotype of CD8+ T cells, 21 –27 the interactions between types of challenge and viral load, 28 and the contribution of CD4+ T lymphocytes 29 –31 ; we also reported an association between increases in cytotoxic T lymphocyte (CTLs) specific for conserved epitopes and vaccine-induced protection. 32 However, the relative contribution of each of these factors to the overall protective response remains unclear. Therefore, it is likely that multiple effector mechanisms are involved in vaccine-induced protection and innate, humoral and cell-mediated immunity may all be required for protection.
We sought to evaluate the acute phase events after challenge with a pathogenic SIV in vaccinated and unvaccinated animals with the intent to identify those immune response factors that contribute to controlling infection. Through the use of a tether system 33 during the first month after challenge, blood samples were obtained from the animals every 48–72 h allowing for a robust examination of acute phase changes without the confounding issues of stress or sedation-related immune response changes. The vaccinated animals were fully protected against SIVmac251 and did not show any signs of viral circulation for more than a year after challenge. In this study, we report that in the vaccinated animals, we observed that NKG2C significantly increased on circulating T cells; also, we report that vaccinated animals have higher circulating levels of CD8+ B cells than naive animals, and this cell subset increases in naive animals after infection with SIVmac.
Material and Methods
Challenge of rhesus macaques
Eight colony-bred, weight- and age-matched adult, Indian female rhesus macaques (Macaca mulatta) were used in this experiment. Animals were seronegative for simian type D retroviruses, simian T cell leukemia virus, and herpes B virus. The animals were used and cared for in accordance with the Association for the Assessment and Accreditation of Laboratory Animal Care (AAALAC) International Guidelines and in accordance with protocols approved by the Texas Biomedical Research Institute Institutional Animal Care and Use Committee (IACUC).
Four animals had received a live attenuated SIV vaccine (SIVmac239Δnef) 2 years before challenge 34 and were considered virus negative as viral loads were below the limit of detection (nucleic acid sequence based amplification assay) and viral isolation efforts were unsuccessful. Four naive animals served as controls. All macaques were challenged by the IV route with 1 ml of RPMI containing 50 TCID50 of SIVmac251. Animals were scheduled to be euthanized if they showed three or more of the following clinical symptoms: (a) weight loss >10% in 2 weeks or 30% in 2 months, (b) chronic diarrhea that was unresponsive to treatment, (c) infections that were unresponsive to antibiotic treatment, (d) inability to maintain body heat or fluids without supplementation, (e) persistent marked hematological abnormalities, including lymphopenia, anemia, thrombocytopenia, or neutropenia, and (f) persistent marked splenomegaly or hepatomegaly.
Tether system
The eight animals were placed on a tether system that allowed for blood sampling without sedation. 32,33 Briefly, 2 weeks before challenge, the animals underwent surgery for the implantation of catheters into the femoral vein. The plastic tubes were tunneled under the skin and exited through a small incision in the animal's back into a protected flexible metal hose. The animals were fitted with a jacket to keep them from being able to touch the exit site. The catheters were attached to a port in the cage by a flexible shaft. Animals could move freely in the cages during inoculation of the virus and drawing of the blood samples, therefore sedation was not needed. The catheters were removed from the animals at 28 days postchallenge (DPC).
Phenotyping of immune cells
Phenotypic characterization of peripheral blood mononuclear cells (PBMCs) was performed by flow cytometry using multicolor direct immunofluorescence. Surface staining was performed by incubating fresh blood or PBMCs with monoclonal antibodies (mAbs) conjugated to different fluorochromes (FITC, PE, PerCP-Cy5.5, APC, and APC-Cy7) for 30 min at 4°. Antibodies to human CD3 (clone SP34-2), CD4 (clone L200), CD8 (clone SK1), HLA-DR (clone L243), CD123 (clone 7G3), CD11c (clone S-HCL3), CD20 (clone 2H7), CD95 (clone DX2), CD28 (clone CD28-2), CD27 (clone M-T271), and Ki67 (clone B56) were from BD Biosciences (San Jose, CA); CD14 (clone MY4) and CD20 (clone B1) were from Beckman-Coulter (Indianapolis, IN); CD16 (clone 3G8) from Life Technologies (Carlsbad, CA); NKG2C (clone 134522) was from R&D Systems (Minneapolis, MN), and CD69 (clone FN50) was from DAKO (Carpinteria, CA). For intracellular detection of Ki67 antigen, samples were incubated with surface antibodies first, and then fixed and permeabilized using the CitoFix-CytoPerm solutions from BD-Biosciences. Incubation with Ki67 was performed in ice-cold 1× CytoPerm and the same solution was used for washing. All samples were fixed with 1.6% alcohol-free formaldehyde before acquisition. Samples were acquired in a Cyan LX flow cytometer (Beckman-Coulter) and data were analyzed with Summit software. Lymphocytes were gated based on their characteristic forward versus side scatter pattern, and a second gate for singlets was established using forward scatter area versus forward scatter height. T cells were identified by CD3 fluorescence, and CD20+ B cells were determined in the CD3--lymphoid population. Memory cells within the CD3+CD4+ or CD3+CD8+ populations were defined as CD95+ CD28lo/hi. Absolute values for cells in whole blood were obtained by combining the percentages obtained by flow cytometry with the values of total white blood cell count per microliter and the differential formula for each animal at each time point.
Multiplex analyses of cytokines in plasma
Plasma samples were assayed for 33 nonhuman primate cytokines and chemokines with the Luminex System as described elsewhere. 35 The analytes included G-CSF, GM-CSF, GRO-α, IFN-α, IFN-γ, IL-1β, IL-1Ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12 (p40), IL-12 (p70), IL-13, IL-15, IL-17, IL-18, IP-10, MCP-1, MDC, MIP-1α, MIP-1β, Perforin, RANTES, sCD40L, sIL-6R, sFASL, TNF-α, and TNF-β. Plasma samples were diluted with two volumes of Serum Dilution Buffer (Millipore, Billerica, MA) and the cytokine concentrations were determined using human cytokines as standards. The plates were measured on the Luminex xMAP 100 system (Luminex Corp., Austin, TX) and analyzed using Masterplex QT 2010 software (MiraiBio Corp., South San Francisco, CA). In addition, we also analyzed the same cytokines in plasma samples from 20 rhesus macaques before and 64 weeks after receiving live attenuated SIV vaccines. 34,36
Statistical analyses
All statistical analyses were performed with GraphPad Prism (GraphPad Software, San Diego, CA;
Results
Live attenuated vaccine protection of macaques challenged with SIVmac251
We previously reported that IV challenge with SIVmac251 resulted in sterilizing immunity for animals vaccinated with a live attenuated vaccine, but rapid infection of naive macaques, with viral loads approaching 107 genome equivalents (gen.eq.)/ml by week 2 postchallenge (Fig. 1A). 32 The tether system was used to obtain serial blood samples from all animals in the absence of restrain or sedation, and lymphocytes were analyzed by flow cytometry for a number of activation and proliferation markers.
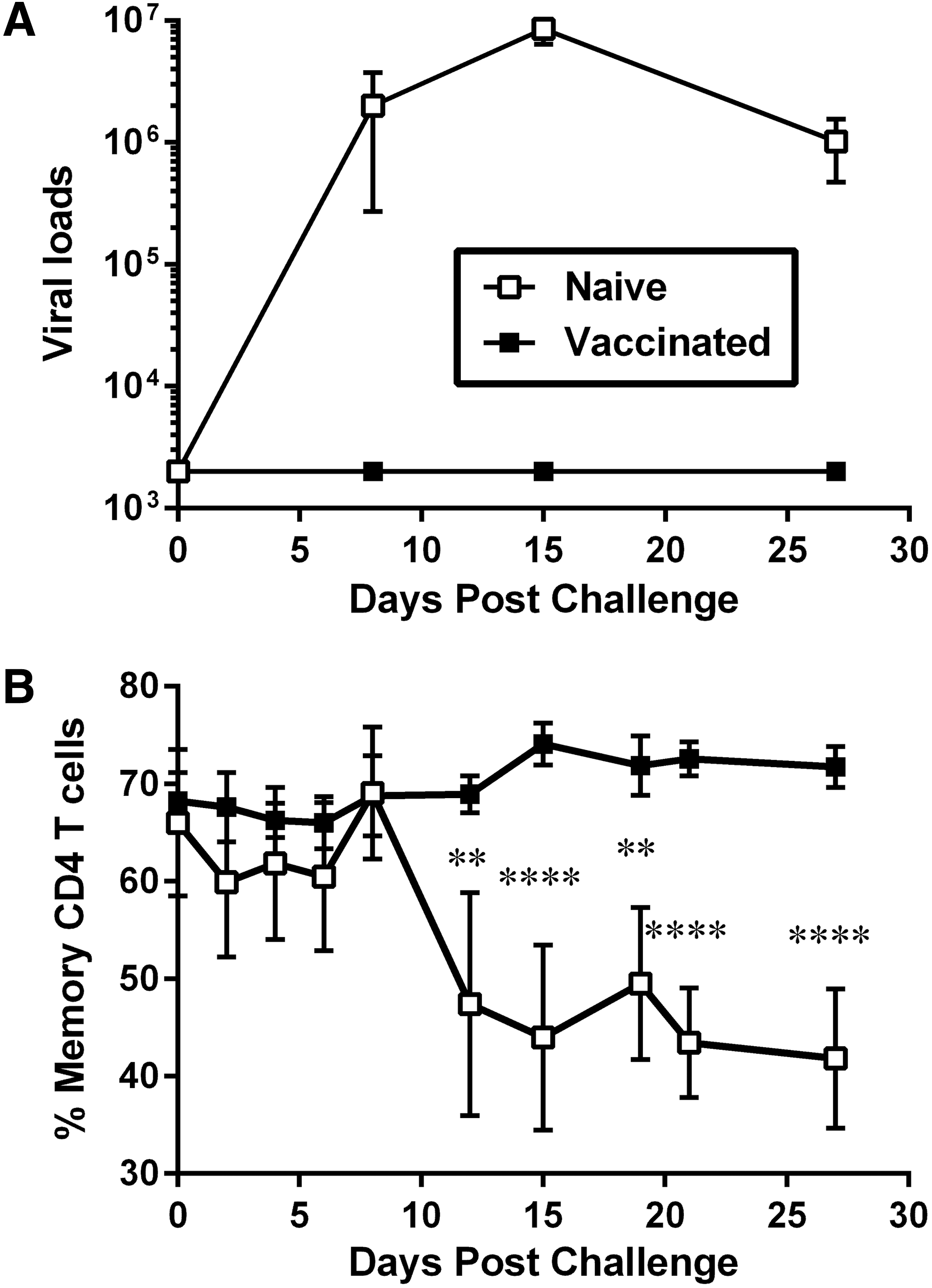
Changes in levels of macaque memory CD4 T cells after IV challenge with SIVmac. Naive macaques (n = 4, open squares) and macaques vaccinated with a live attenuated SIV vaccine (n = 4; closed squares) were challenged with 50 TCID50 of SIVmac251 by the IV route. Blood samples were obtained every 2–3 days with the use of a tether system.
Analysis of circulating memory CD4 T cells, defined as CD3+CD4+CD95Hi, revealed that while vaccinated animals maintained a constant percentage of memory T cells after challenge (∼70% of CD4 T cells), unvaccinated animals lost memory T cells after 12 DPC (∼45% of CD4 T cells), concomitant with the peak of viremia (Fig. 1B). These low levels of memory CD4 T cells remained significantly lower for the naive animals compared to levels of vaccinated macaques for the remaining of the acute phase postchallenge.
Changes in lymphocyte proliferation after challenge with SIVmac251
Levels of lymphocyte proliferation after challenge were measured by flow cytometry by determining expression of the nuclear antigen Ki67 on CD4 T cells (Fig. 2A), CD8 T cells (Fig. 2B), and B cells (Fig. 2C). After challenge, both naive and vaccinated animals experienced a transient decline in proliferating CD4 T cells that was significant for vaccinated animals at 10 and 15 DPC (mean of 9.3% at 12 DPC and 9.2% at 15 DPC, compared to prechallenge values of 14.6% of Ki67+ CD4 T cells) and extremely significant for naive macaques (mean of 2.6% at 12 DPC compared to prechallenge values of 11.1% of Ki67+ CD4 T cells). Proliferation of CD4 T cells in naive animals increased after 15 DPC and returned to similar levels to the ones seen before challenge (Fig. 2A).
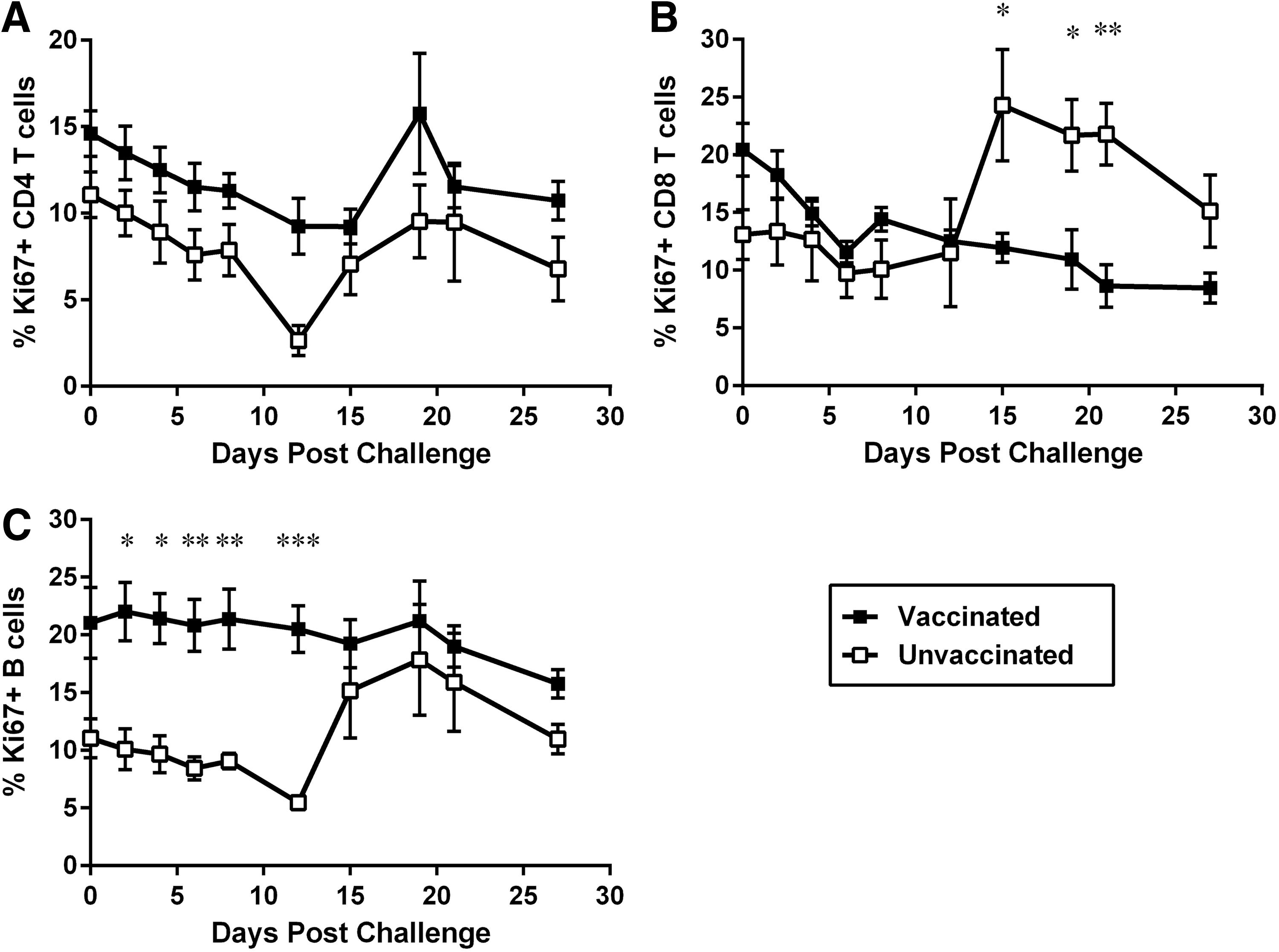
Changes in proliferation levels of macaque lymphocytes after IV challenge with SIVmac. Naive macaques (n = 4, open squares) and macaques vaccinated with a live attenuated SIV vaccine (n = 4; closed squares) were challenged with 50 TCID50 of SIVmac251 by the IV route. Blood samples were obtained every 2–3 days with the use of a tether system, and the percentage of proliferating Ki67+ cells was determined by flow cytometry.
Proliferation of CD8 T cells also showed a constant decline for vaccinated animals, which was very significant compared to prechallenge values by 21 and 28 DPC (mean of 8.6% at 21 DPC and 8.4% at 28 DPC, compared with prechallenge means of 20.5% of Ki67+ CD8 T cells). On the contrary, there was a significant increase in proliferation of CD8 T cells for naive animals, which peaked by 15 DPC (24.3% at 15 DPC compared with prechallenge mean of 13.1% of Ki67+ CD8 T cells). Differences in CD8 T-cell proliferation were significant between vaccinated and naive animals after 15 DPC (24.3% for naive vs. 11.9% Ki67+ CD8 T cells for vaccinated macaques; Fig. 2B).
Finally, analysis of B cells showed that vaccinated animals presented significantly higher levels of proliferating B cells before challenge (mean of 11.1% for naive vs. 21% of Ki67+ B cells for vaccinated macaques before challenge), with these levels not changing significantly after exposure to SIV. For naive macaques, on the contrary, B cell proliferation increased at around 15 DPC and became similar to what was seen for vaccinated macaques (mean of 15.2% of Ki67+ B cells for naive macaques at 15 DPC, Fig. 2C).
Changes in atypical lymphocyte markers after exposure to SIVmac251
Lymphocytes from both vaccinated and unvaccinated animals were analyzed by flow cytometry for various atypical markers, such as expression of CD20 (a B cell marker) on CD8 T cells or expression of CD8 α chain (mainly present in T and NK cells) by B cells.
CD3+CD8+ T cells expressed dimmer levels of CD20 [CD20 mean fluorescence intensity (MFI) between 101 and 102], compared to ones seen for CD3- B cells (CD20 MFI between 102 and 103, Fig. 3A). CD3+CD8+ T cells expressing CD20 were below 1% of the total T cells in both groups before challenge with SIVmac251 (mean of 9 cells/μl in naive and 4 CD8+ CD20+ T cells/μl for vaccinated macaques) and remained low in vaccinated macaques throughout the acute phase of infection. However, naive animals experienced a dramatic increase in CD20+ CD8 T cells after 19 DPC, which peaked by 21 DPC (mean of 99 cells/μl in naive vs. 7 CD8+ CD20+ T cells/μl in vaccinated macaques) and remained significantly elevated by 27 DPC (Fig. 3B).
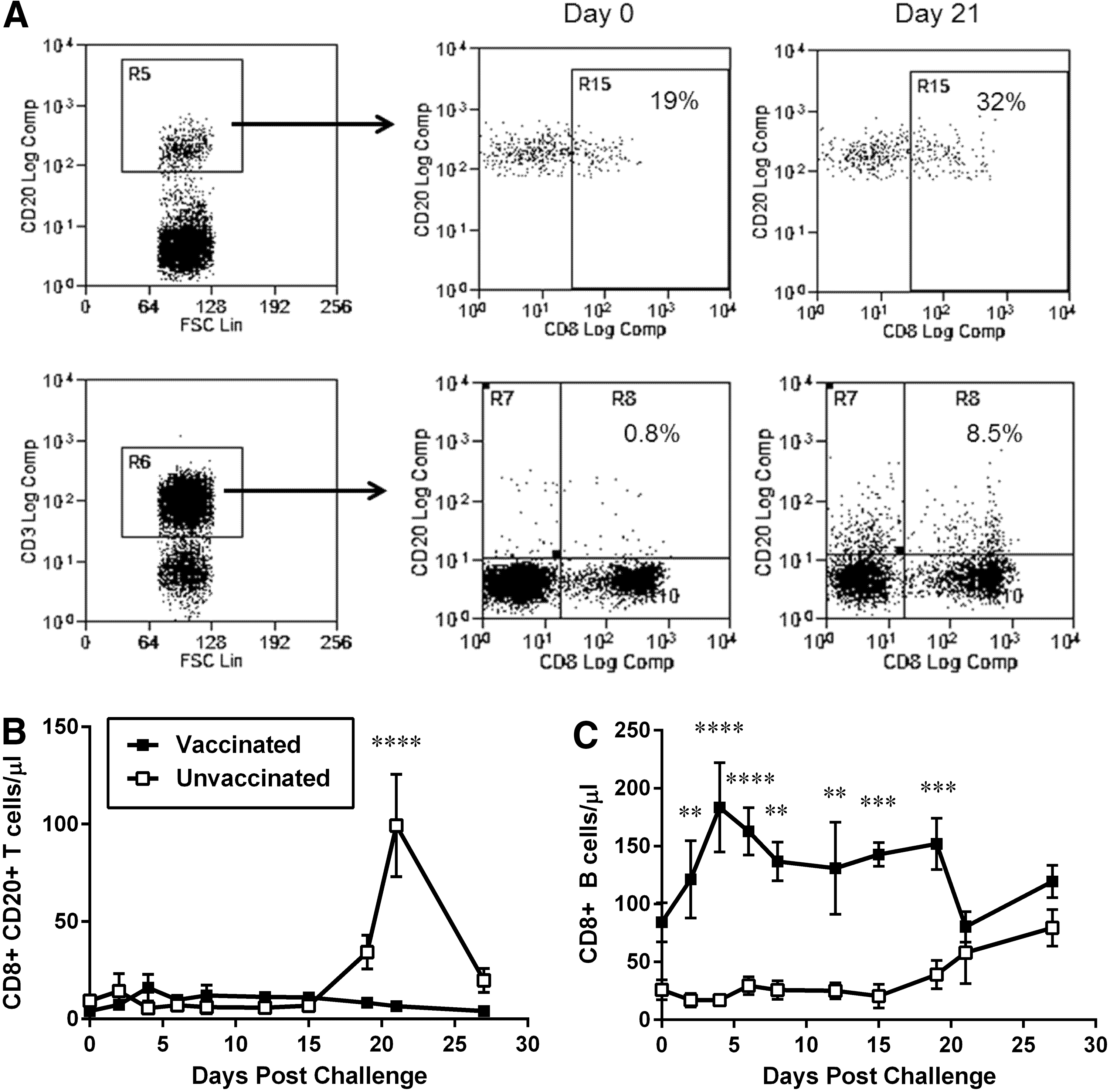
Changes in expression levels of atypical markers on macaque lymphocytes after IV challenge with SIVmac. Naive macaques (n = 4, open squares) and macaques vaccinated with a live attenuated SIV vaccine (n = 4; closed squares) were challenged with 50 TCID50 of SIVmac251 by the IV route. Blood samples were obtained every 2–3 days with the use of a tether system and cell subsets and markers were identified by flow cytometry.
Similarly, CD3-CD20+ B cells also expressed dim levels of CD8 α (CD8 MFI between 101 and 102), compared to what is usually seen for CD8 CD3 T cells (CD8 MFI between 102 and 103, Fig. 4A). Interestingly, CD3-CD20+ B cells expressing CD8α were significantly more numerous in vaccinated animals compared to naive macaques, even before challenge with pathogenic SIV (mean of 26 cells/μl in naive vs. 84 CD8+ B cells/μl in vaccinated macaques); these cells had a transient, but significant elevation by 4 DPC (mean 184 CD8+ B cells/μl in vaccinated macaques) and remained stable during the duration of the study. For naive macaques, infection with SIV resulted in a gradual increase in the percentage and absolute values of B cells expressing CD8, which reached levels similar to the ones seen for vaccinated macaques by the end of the acute infection period (mean of 80 cells/μl in naive vs. 120 CD8+ B cells/μl in vaccinated macaques; Fig. 3C).
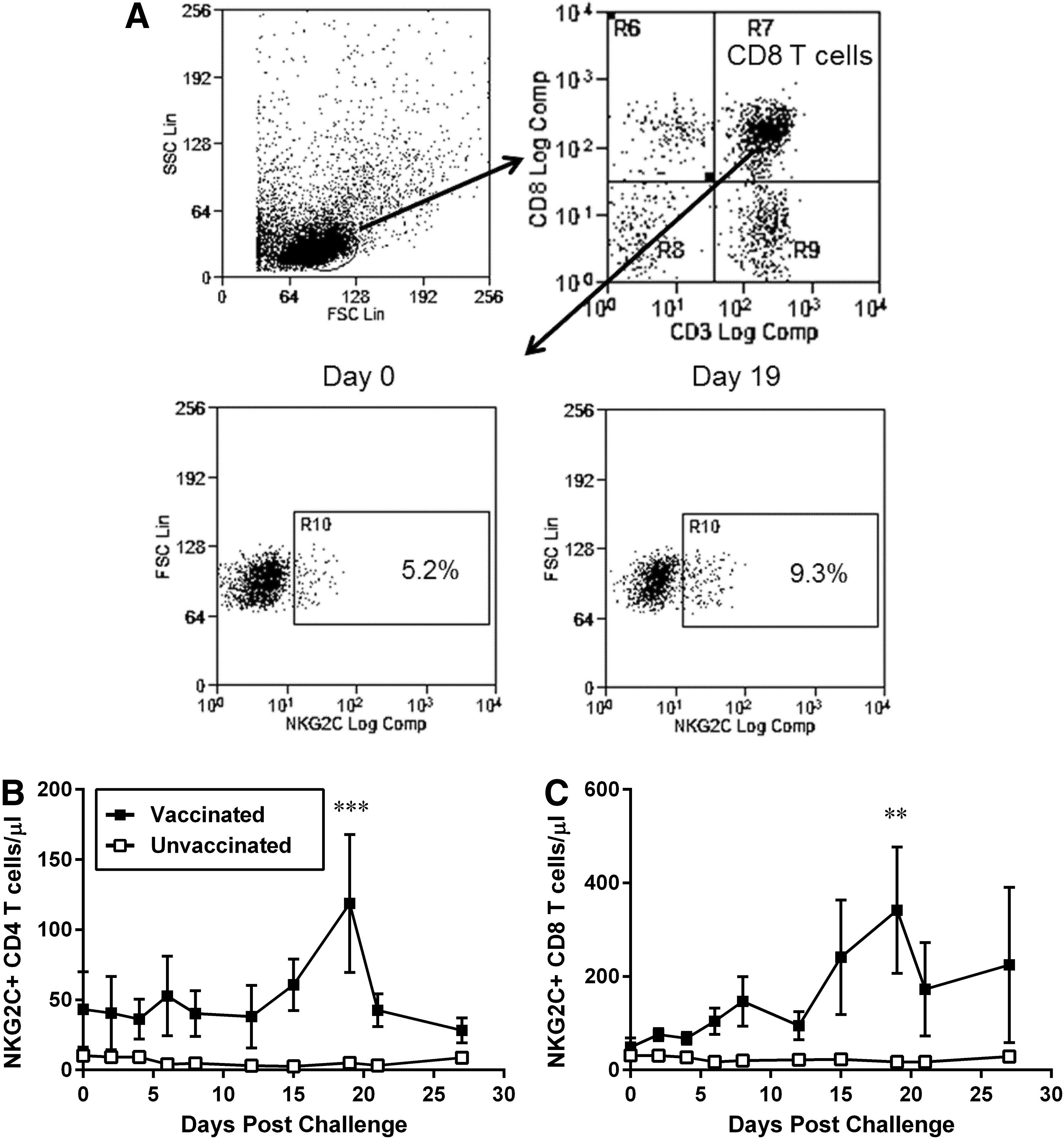
Changes in expression levels of the activating receptor NKG2C on macaque lymphocytes after IV challenge with SIVmac. Naive macaques (n = 4, open squares) and macaques vaccinated with a live attenuated SIV vaccine (n = 4; closed squares) were challenged with 50 TCID50 of SIVmac251 by the IV route. Blood samples were obtained every 2–3 days with the use of a tether system, and the expression of NKG2C was determined by flow cytometry.
Variation in NKG2C expression on T cells after challenge with SIVmac251
The level of expression of the natural cytotoxicity activating receptor NKG2C was evaluated by flow cytometry in several lymphocyte subsets, including CD4+ T and CD8+ T cells, in both groups of challenged animals. Gating for either CD4 or CD8 T cells was performed for CD3+ CD4+ CD8− or CD3+ CD4− CD8+, respectively, therefore excluding CD4+CD8+ T cells (Fig. 4A). The levels of CD4 T cells expressing NKG2C were low in both groups of animals before challenge (mean of 11 cells/μl in naive vs. 43 NKG2C+ CD4 T cells/μl in vaccinated macaques: Fig. 4B). While these levels remained unchanged in CD4 T cells from naive animals after challenge, NKG2C+ CD4 T cells from vaccinated animals showed an increase by 15 DPC that peaked by 19 DPC (mean of 119 NKG2C+ CD4 T cells/μl in vaccinated macaques).
Expression of NKG2C on CD8 T cells also showed no significant variation for unvaccinated macaques (mean of 31 cells/μl prechallenge vs. 17 NKG2C+ CD8 T cells/μl at 19 DPC in naive); however, vaccinated macaques had generally higher levels of NKG2C+ CD8 T cells at all time points, which also peaked by 19 DPC (mean of 49 cells/μl prechallenge vs. 342 NKG2C+ CD8 T cells/μl at 19 DPC in vaccinates macaques; Fig. 4C).
Proinflammatory cytokines and perforin do not increase postchallenge in vaccinated animals
The variation in the concentration of plasma cytokines as a consequence of challenge with SIVmac251 was measured with Luminex Technology, which allowed for the simultaneous identification of 33 cytokines in a single small volume of sample. Unvaccinated macaques exposed to pathogenic SIV showed the expected increase in concentration of proinflammatory cytokines and effector molecules associated with acute infection. These animals had transient increases in IFN-α, IP10, MCP-1, Perforin, IFN-γ, IL-1Ra, IL-8, and IL-18 (data not shown). In general, cytokine concentrations in unvaccinated animals varied in parallel with viral loads, reaching concentration peaks by 9–12 DPC. On the contrary, no variation in the levels of all cytokines that were measured was evident for the macaques vaccinated with the live attenuated vaccine; one interesting characteristic of these vaccinated animals was that they had constitutively higher levels of IFN-α than the naive macaques before and after peak viremia; however, these differences were not statistically significant. As a consequence of this finding and to identify significant long-term changes induced by live attenuated vaccines, cytokines were measured in the plasma of 20 macaques that had received SIVmac Δnef-based vaccines. 34,37 The samples that were assayed included prevaccination time points and samples obtained 64 weeks after vaccination, when vaccine viral loads were below the limit of detection. Analysis of the data showed that vaccination with live attenuated vaccines resulted in significant increases in the concentration of IP-10 (mean of 361 pg/ml prevaccination vs. 553.4 pg/ml at 64 weeks postvaccination) and perforin (mean of 316.2 pg/ml prevaccination vs. 493 pg/ml at 64 weeks postvaccination; Fig. 5).
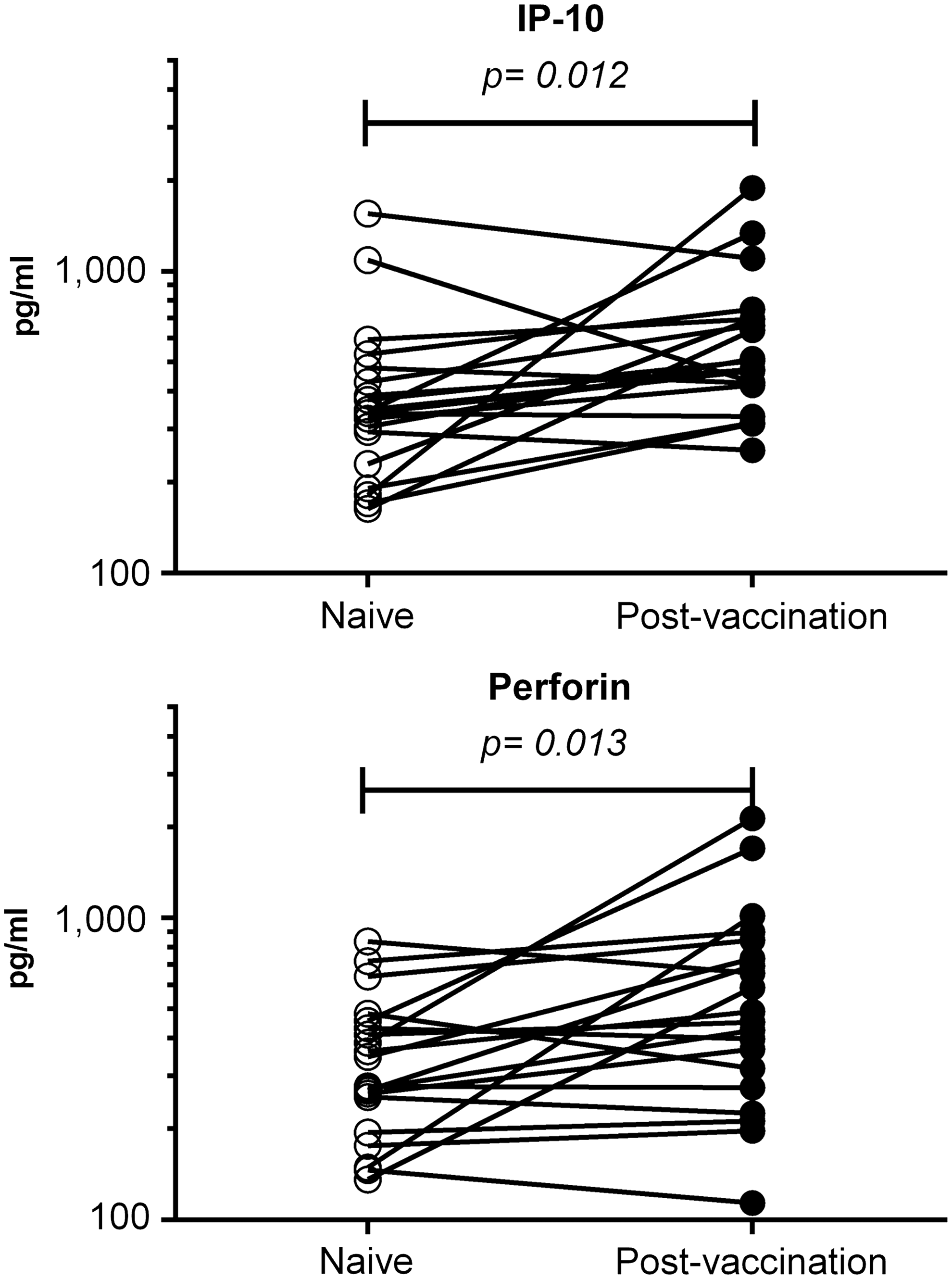
Changes in concentration of plasma proinflammatory cytokines in macaques after vaccination with SIVmacΔnef-based vaccines. Plasma samples were obtained from 20 rhesus macaques before (open circles) and 64 weeks after (closed circles) vaccination with SIVΔnef-based vaccines. 34,37 A total of 33 cytokines, chemokines, and other molecules were measured with an nonhuman primate-validated Luminex assay. Levels of IP-10 and perforin were significantly elevated in vaccinated macaques.
Discussion
The tether sampling system makes possible the evaluation of multiple immune response factors at multiple time points within the acute phase of infection. In our previous publication we demonstrated that vaccination with live attenuated SIV resulted in sterilizing protection against IV challenge with pathogenic SIVmac251 (Fig. 1A), which was associated with expansion of cytotoxic T cells specific for immunodominant epitopes in the presence of high anti-SIV antibody titers. 32 Our next step was to evaluate by flow cytometry various cell phenotype, activation, and proliferation markers to identify and better understand factors that may be indicative of protection from infection. Our vaccinated animals had received the live attenuated SIVΔnef vaccine 34 2 years before these challenge studies and were challenged at a time when the levels of circulating vaccine virus were below the limit of detection for SIV RNA assay. As displayed in Figure 1B, we demonstrated that vaccinated animals maintained unchanged their population of memory CD4 T cells, while unvaccinated animals began losing memory CD4 T cells a week after infection, which is consistent with reports of memory CD4 T cells as targets of SIV infection. 38 Similarly, analysis of proliferation for CD4 T cells from naive animals showed a profound decline by 12 DPC, consistent with the peak of viral replication (Fig. 2A). 39 When studying lymphocyte proliferation of vaccinated macaques, either no change or slight decreases were observed, with the exception of a brief peak of CD4 T-cell replication by 21 DPC (Fig. 2A). It is possible that 2 years of antigenic stimulation by the live attenuated vaccine resulted in anergy of these cell populations, similar to what has been reported for NK cells in chronic infection 40 ; this would also be consistent with the observed transient CD4 T-cell proliferation, which is the cell population reported to be most resistant to anergy in chronic SIV infection. 41 The increase in proliferation for CD8 T cells and B cells in naive animals at the time of peak infection is consistent with the acute primary immune responses classically seen in this model. 42,43 One novel finding of our study was that the prechallenge levels of proliferating B cells were significantly higher in vaccinated macaques compared to naive animals (Fig. 2C). This higher level of proliferating B cells may reflect the persistence of the vaccine virus in CD4 T cells present in immune privileged areas of secondary lymphoid organs, which has been shown to drive increased turnover of B cells. 20,44
Another novel finding of this experiment was the discovery that CD8+ T cells in naive animals transiently increase the level of expression of the B cell marker CD20 during the acute phase of SIV infection (Fig. 3B). The significance of this dim CD20 expression is not clear, but CD8+CD20+ T cells have been shown to be present in patients with rheumatoid arthritis, 45 multiple sclerosis, 46,47 and ovarian cancer. 48 de Bruyn et al. reported that CD20 is transferred from B cells to T cells upon intimate T-cell/B cell interaction in ovarian cancer patients 48 ; also, in the context of multiple sclerosis, CD20+CD8+ T cells have been shown to have inflammatory characteristics and primarily produce IFN-γ. 46 All this information, combined with the transient nature of this CD20 expression by CD8 T cells occurring after peak of viral replication, suggest that these CD20+ CD8 T cells may also originate by T-cell/B cell contact in inflamed lymphoid tissues during primary immune responses to SIV infection. Further studies will be needed to demonstrate this hypothesis.
We also observed in vaccinated animals that, before challenge with SIV, about 30% of all CD20+ B cells expressed the CD8 marker (Fig. 3C). There have been reports of aberrant expression of CD8 by B cells, but they have been in the context of chronic lymphocytic leukemia 49,50 or Non-Hodgkin lymphoma. 51 Steady increased expression of CD8 by B cells was also seen for naive macaques after infection with SIV, reaching similar levels to the ones seen for vaccinated macaques by 3 weeks postchallenge. This pattern of CD8 expression on B cells follows similar trends to the ones seen for expression Ki67 on B cells (Fig. 2C), which suggests that CD8 expression may be a marker of B cell activation and proliferation.
The CD94/NKG2 family of NK cell receptors has been identified in humans, chimpanzees, orangutans, rhesus monkeys, and rodents. These receptors are heterodimers composed of the CD94 molecule covalently bound to activating or inhibitory NKG2 molecules; upon cross-linking of the activating CD94/NKG2C complex, a signaling cascade is initiated that leads to activation of NK cell-mediated lysis. 52 Besides NK cells, CD94/NKG2C expression has been identified in a subset of CD8+ T cells with a highly differentiated effector phenotype 53 and in small numbers of TCR γδ and rare CD4 T cells, suggesting that CD94/NKG2C may constitute an alternative T-cell activation pathway capable of driving the expansion and triggering the effector functions of a CTL subset. 54 Consistent with these reports, it is interesting to note that the only remarkable changes that we observed in vaccinated macaques after challenge were significant increases in the expression of NKG2C on CD4 and CD8 T cells, around 3 weeks postchallenge (Fig. 4B, C). Although these changes in NKG2C expression that we measured were systemic, that is, not detected exclusively in SIV-specific T cells, they paralleled the increase in SIV-specific CTL that we previously reported 32 and the induction of T-cell activation associated with vaccine protection reported by others. 16
The use of the tether system minimized any variation in cytokine concentration due to repeated sedation and stress, which makes this approach the ideal way to investigate changes in the levels of various circulating cytokines that were due solely to infection. This is best illustrated by the stable concentration levels seen after SIV challenge in vaccinated macaques, where sterilizing immunity resulted in no significant variation of any of the 33 cytokines that we measured (data not shown). On the other hand, naive macaques showed the typical increase of proinflammatory cytokines and perforin induced by viral infection. It is interesting to note that vaccinated macaques had elevated levels of IP-10 and perforin in circulation more than a year after vaccination (Fig. 5); higher systemic levels of IP-10 and perforin could be attributed to vaccine virus persistence in lymphoid tissues, even though circulating vaccine virus was below the limit of detection of the viral RNA assay. Increases of IP-10 in infected naive macaques (Fig. 5) are consistent with IP-10 being an indicator of increased viral load in patients with previous control of HIV infection and being associated with an increase in CD4 and CD8 T-cell activation. 55 On the other hand, perforin has been proposed to have an immunoregulatory role, which is distinguishable from its pathogen clearance function, by suppressing antigen presentation by limiting T-cell activation in the context of chronic infection, 56 suggesting that the perforin values seen in vaccinated animals may help to control T-cell activation state.
In summary, these studies corroborate the hypothesis that multiple factors are responsible for protection against SIV infection induced by live attenuated vaccines. In addition, we provide the first report that B cells in these vaccinated animals have significant turnover and activation, which is associated with elevated Ki67 and CD8 expression, and SIV challenge exposure results in increased expression of the activating NK receptor NKG2C on T cells. Finally, we also describe that SIV infection in naive macaques induces the transient appearance of CD20 on activated T cells. Our report supports the utility of the tether system for the intensive study of acute immune responses to viral infections.
Footnotes
Acknowledgments
The authors thank the veterinary personnel from the Southwest National Primate Research Center of the Texas Biomedical Research Institute for assistance with the animal manipulations. This work was supported by the US National Institutes of Health (grant nos. R01 AI41923, R21 AI44363, and P51 OD011133).
Author Disclosure Statement
No competing financial interests exist.