
Abstract
Associations of regulated on activation, normal T cell expressed and secreted (RANTES) −403G/A, −28C/G, and In1.1T/C polymorphisms with HIV-1 infection and the progression of HIV-1 disease have been widely reported with inconsistent results. To clarify this situation, we performed an updated meta-analysis of all available studies from PubMed, EMBASE, and the China National Knowledge Infrastructure. A total of 24 eligible studies involving more than 10,000 subjects were included. By using the healthy controls, we found that −403G/A polymorphism was significantly associated with reduced susceptibility to HIV-1 infection in G/A+A/A versus GG (odds ratio [OR] = 0.755, 95% confidence interval [CI] = 0.581–0.982); and a significantly decreased risk was also found for −28C/G polymorphisms (G vs. C, OR = 0.804, 95% CI = 0.696–0.927; G/G+C/G vs. C/C, OR = 0.826, 95% CI = 0.704–0.969). Whereas for In1.1T/C polymorphism, increased risk of HIV-1 infection was revealed (C vs. T, OR = 1.216, 95% CI = 1.047–1.430; T/C vs. T/T, OR = 1.68, 95% CI = 1.263–2.234; T/C+T/T vs. C/C, OR = 1.466, 95% CI = 1.147–1.875). Subgroup analyses by ethnicity showed significant association among Asians, but not among Caucasians. When HIV-1-exposed seronegative (HESN) controls were used, no significant association was detected. Moreover, −403G/A and −28C/G polymorphisms were also not associated with long-term nonprogressive HIV-1 infection (all p > .05). This meta-analysis suggests that RANTES −403G/A and −28C/G polymorphisms confer possible protection against HIV-1 infection, whereas In1.1T/C polymorphism may increase risk of HIV-1 infection, especially in Asians. These results may contribute to finding a theoretical basis for effective control strategies against HIV/AIDS. Further investigations are needed to validate our conclusions.
Introduction
S
Regulated on activation, normal T cell expressed and secreted (RANTES, also known as CC chemokine ligand 5, or CCL5) is one of the natural ligands for HIV-1 coreceptors, which together with macrophage inflammatory protein (MIP)-1α and MIP-1β constitute the major HIV suppressive factors. 8 RANTES acts by blocking access of the HIV envelope glycoprotein gp120 to CC chemokine receptor 5 (CCR5) and by reducing surface levels of CCR5. 9 Moreover, cluster of differentiation 4-positive T lymphocytes (CD4+ T cells) from highly HIV-1-exposed seronegative individuals (HESNs) produce much more RANTES than those from random blood donor controls. 10 Smaller amounts of RANTES are produced by immortalized CD4+ T cells from patients with acquired immunodeficiency syndrome disease (AIDS) than from those produced by LTNPs. 11 However, it was also found that higher levels of RANTES in serum are associated with faster disease progression. 12 These findings suggest that RANTES expression may affect the risk of HIV-1 infection and the progression of HIV-1 disease. The gene encoding RANTES is located on chromosome17q11.2–q12. 13 Several frequent single-nucleotide polymorphisms (SNPs) in the RANTES gene have been reported to up- or downregulate RANTES, including −403G/A and −28C/G in the promoter region and In1.1T/C in the first intron region. 14 –16 Therefore, these three RANTES polymorphisms may affect the course of HIV-1 infection by modulating the transcription and expression of the RANTES gene.
Several studies have pointed to the potential association between the RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms and risk of HIV-1 infection and disease progression, 15,17 –35 however, the data were inconclusive or contradictory. This inconsistency may be due to problems such as low statistical power, racial differences, publication biases, and uncorrected multiple hypothesis testing. Previous meta-analyses 36,37 found that the RANTES −403G/A and −28C/G polymorphisms may play a role in resistance to HIV-1 infection among Asians; recently, additional studies have been published, 34,35 and further studies with stricter search criteria, methodological quality assessment of the studies/data selected, and investigation of disease progression must be conducted, which may be more reliable and lead to more accurate results., No meta-analyses have been performed with respect to In1.1T/C polymorphism. Thus, we conducted a comprehensive meta-analysis to evaluate the effects of RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms on susceptibility to HIV-1 infection and disease progression.
Materials and Methods
Literature search
Literatures were searched within PubMed, EMBASE, and the China National Knowledge Infrastructure (CNKI) using the following key words: HIV/AIDS/human immunodeficiency viruses, RANTES/CCL5, and polymorphism/variant. The latest search occurred in August 2015, without any language or country restriction. We also screened the references of retrieved articles to identify additional studies. In studies that did not include the required information, we contacted the authors and requested that they forward the relevant data through e-mail within 2 weeks.
Inclusion and exclusion criteria
The eligible studies met the following criteria: (a) case–control or cohort studies reporting the association between the RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms and HIV-1 infection susceptibility and disease progression; (b) studies involving HIV-1 infected cases and HIV-1 uninfected controls; and (c) studies providing sufficient data to estimate the odds ratios (ORs) and 95% confidence intervals (CIs). In addition, the exclusion criteria were as follows: (a) abstracts, reviews, and animal studies; (b) studies in which the requested information was not obtained by contacting the authors within 2 weeks; and (c) when multiple studies reported the same or overlapping data, we selected the latest one or the one providing more amount of information.
Data extraction and qualitative assessment
Study characteristics, including first author, year of publication, country of studied population, ethnicity, sample size of cases (LNTPs and/or usual progressors [Ups]) and controls (healthy controls [HCs] and/or HESNs), source of controls, genotype distribution in cases and controls, and p-value for controls in the Hardy–Weinberg equilibrium (HWE), were abstracted by two authors independently. If different results were generated, the disagreements were resolved by group discussion.
Two authors completed the quality assessment independently. A set of predefined criteria (Supplementary Table 1; Supplementary Data are available online at
Data analysis
The pooled ORs with 95% CIs were estimated for each polymorphism in different comparison models, including the allele model, codominant model, recessive genetic model, and dominant genetic model.
For each genetic contrast, heterogeneity was assessed by a χ 2-based Q statistic test. 39 If the results were heterogeneous (p < .10 or I 2 ≥ 50%), the pooled ORs were calculated with a random-effects model (the DerSimonian–Laird method). 40 When p ≥ .10 and I 2 < 50%, the between-study heterogeneity was considered not significant, and a fixed-effects model (the Mantel–Haenszel method) 41 was used. To further explore causes of heterogeneity, we performed logistic meta-regression and subgroup analyses. 42 The study characteristics, including ethnicity (Caucasian population versus Asian population versus others), source of controls (hospital-based [HB] versus population-based [PB]), genotyping methods (polymerase chain reaction–restriction fragment length polymorphism [PCR-RFLP] versus non-PCR-RFLP), and quality score (high-quality studies versus low-quality studies), were used as covariates in the logistic meta-regression analyses, and p < .05 was considered statistically significant. Subgroup analyses were conducted by stratification of ethnicity, source of controls, genotyping methods, and quality scores. Publication bias was evaluated by Egger's and Begg's tests (considered significant for p < .05). 43,44 To assess the stability of the results, we performed a sensitivity analysis by sequential omission of individual studies. 36 The HWE of the controls was calculated with a chi-square (X 2) test, and p < .05 was considered a statistically significant deviation from HWE. 45
Stata software, version 12.0 (Stata Corporation, College Station, TX), was used to conduct the statistical analysis. All p-values were two sided.
Results
Study characteristics
As shown in Figure 1, 210 relevant articles were identified through a search of the literature, of which 20 articles were determined to be eligible for further analysis. No additional articles were found by manual search of references cited in the published studies. Hence, 20 articles met the inclusion criteria for the meta-analysis. 15,17 –35 Among these, two eligible studies contained data on two different ethnic groups that were independently evaluated 17,19 ; one study contained data on three different ethnic groups. 23 We then treated them separately. As a result, a total of 24 separate studies were finally included in our meta-analysis. Among them, 19 high-quality studies, including 5,224 cases and 3,677 controls (3,262 HCs and 415 HESNs), were evaluated for the −403G/A variant, 16 high-quality studies, including 4,922 cases and 3,387 controls (3,008 HCs and 379 HESNs), were included for −28C/G polymorphism, and eight high-quality studies containing 1,208 cases and 970 HCs were evaluated for the In1.1T/C mutation. Table 1 shows the main characteristics of the included studies.

Flow diagram of study selection and exclusion.
PCR–RFLP, polymerase chain reaction–restriction fragment length polymorphism; PCR-SDCT, polymerase chain reaction-sequence with dideoxy-chain-termination method; RTPCR-FRET, real-time polymerase chain reaction-fluorescence resonance energy transfer; PCR-SSPs, polymerase chain reaction-sequence with specific primers; LTNP, long-term nonprogressors; UP, usual progressors; NA, not available; HC, healthy controls; HESN, HIV-1-exposed seronegative individuals; PB, population based; HB, hospital based; HWE, Hardy–Weinberg equilibrium of genotypes of controls.
RANTES polymorphisms and their association with HIV-1 infection
For the −403G/A polymorphism, the results suggest that there may be a significant association between the −403G/A polymorphism and reduced susceptibility to HIV-1 infection in the recessive genetic model (A/A vs. G/A+G/G, OR = 0.755, 95% CI = 0.581–0.982, p = .036) (Fig. 2A), using the healthy controls (Table 2). When we performed subgroup analyses according to ethnicity, we also found a significantly decreased HIV-1 infection risk among Asians (A/A vs. G/A+G/G, OR = 0.663, 95% CI = 0.533–0.862, p < .001) and Africans (allele model A vs. G, OR = 0.736, 95% CI = 0.612–0.884, p = .001; codominant model (a) A/A vs. G/G, OR = 0.538, 95% CI = 0.363–0.796, p = .002, (b) G/A vs. G/G, OR = 0.582, 95% CI = 0.428–0.792, p = .001; dominant genetic model A/A+G/A vs. G/G, OR = 0.569, 95% CI = 0.436–0.762, p < .001), but not among Caucasians (p > .05 for all genetic models) (Table 2). In the subgroup analyses stratified by source of controls, HWE, and genotyping methods, a statistically reduced HIV/AIDS risk was also observed in hospital-based, PCR-RFLP, HWE (Yes), and HWE (No) groups (Table 2). However, by using HESN controls, we failed to identify any significant association between the −403G/A polymorphism and risk of HIV-1 infection (Fig. 3A).

Forest plot of the association between the RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms and susceptibility to HIV-1 infection, using healthy controls.

Codominant model (a), the comparison of a variant homozygote and a wild homozygote; codominant model (b), the comparison of a heterozygote and a wild homozygote; c p-value for OR; d Model of meta-analysis; F, fixed-effects model; R, random-effects model; “—,” insufficient data for analysis.
The bold values mark the significant genetic models (p < 0.05).
For the −28C/G polymorphism, using healthy individuals as controls, significantly decreased risk of HIV-1 infection was revealed in the overall population (allele model G vs. C, OR = 0.804, 95% CI = 0.696–0.927, p = .003, [Fig. 2B]; dominant genetic model G/G+C/G vs. C/C, OR = 0.826, 95% CI = 0.704–0.969, p = .019). The stratified analyses revealed a significantly decreased susceptibility to HIV-1 infection among Asians (G vs. C, OR = 0.747, 95% CI = 0.586–0.953, p = .019); codominant model (a) G/G vs. C/C, OR = 0.372, 95% CI = 0.204–0.677, p = .001, (b) C/G vs. C/C, OR = 0.831, 95% CI = 0.692–0.999, p = .049; recessive genetic model G/G vs. C/G+C/C, OR = 0.428, 95% CI = 0.253–0.723, p = .002; C/G+G/G vs. C/C, OR = 0.784, 95% CI = 0.656–0.938, p = .008), PB, HB, and PCR-RFLP groups. We failed to find a significant association among Caucasians and non-PCR-RFLP studies (Table 2). By using HESN controls, no association between the polymorphism −28C/G and risk of HIV-1 infection was detected in the allelic model (Fig. 3B) or in the rest of the comparisons (data not shown).
However, as Table 2 shows, the results indicated that In1.1T/C polymorphism was associated with increased risk of HIV-1 infection based on the pooling of data from a number of different ethnic groups (allele model C vs. T, OR = 1.216, 95% CI = 1.047–1.430, p = .011, [Fig. 2C]; codominant model (b) C/T vs. T/T, OR = 1.680, 95% CI = 1.263–2.234, p < .001; dominant genetic model T/C+C/C vs. TT, OR = 1.466, 95% CI = 1.147–1.875, p = .002). When the subgroup analyses were categorized by ethnicity, significantly increased risk of HIV-1 infection was found among Asians (C vs. T, OR = 1.239, 95% CI = 1.023–1.501, p = .028; C/T vs. T/T, OR = 1.739, 95% CI = 1.249–2.421, p = .001; T/C+C/C vs. TT, OR = 1.455, 95% CI = 1.106–1.915, p = .007), but not among Caucasians (all p > .05 for all genetic models). When stratified by source of control, genotyping methods, and HWE, a statistically increased risk of HIV-1 infection was also found in the PB, non-PCR-RFLP, HWE (Yes), and HWE (No) groups (Table 2).
Interestingly, the data of −403G/A and −28C/G polymorphisms for AIDS progression (LTNPs vs. UPs) are available. Therefore, we performed meta-analyses about the effects of −403G/A and −28C/G polymorphisms on disease progression. However, −403G/A and −28C/G polymorphisms of the RANTES gene were not associated with long-term nonprogressive HIV-1 infection (Fig. 4).
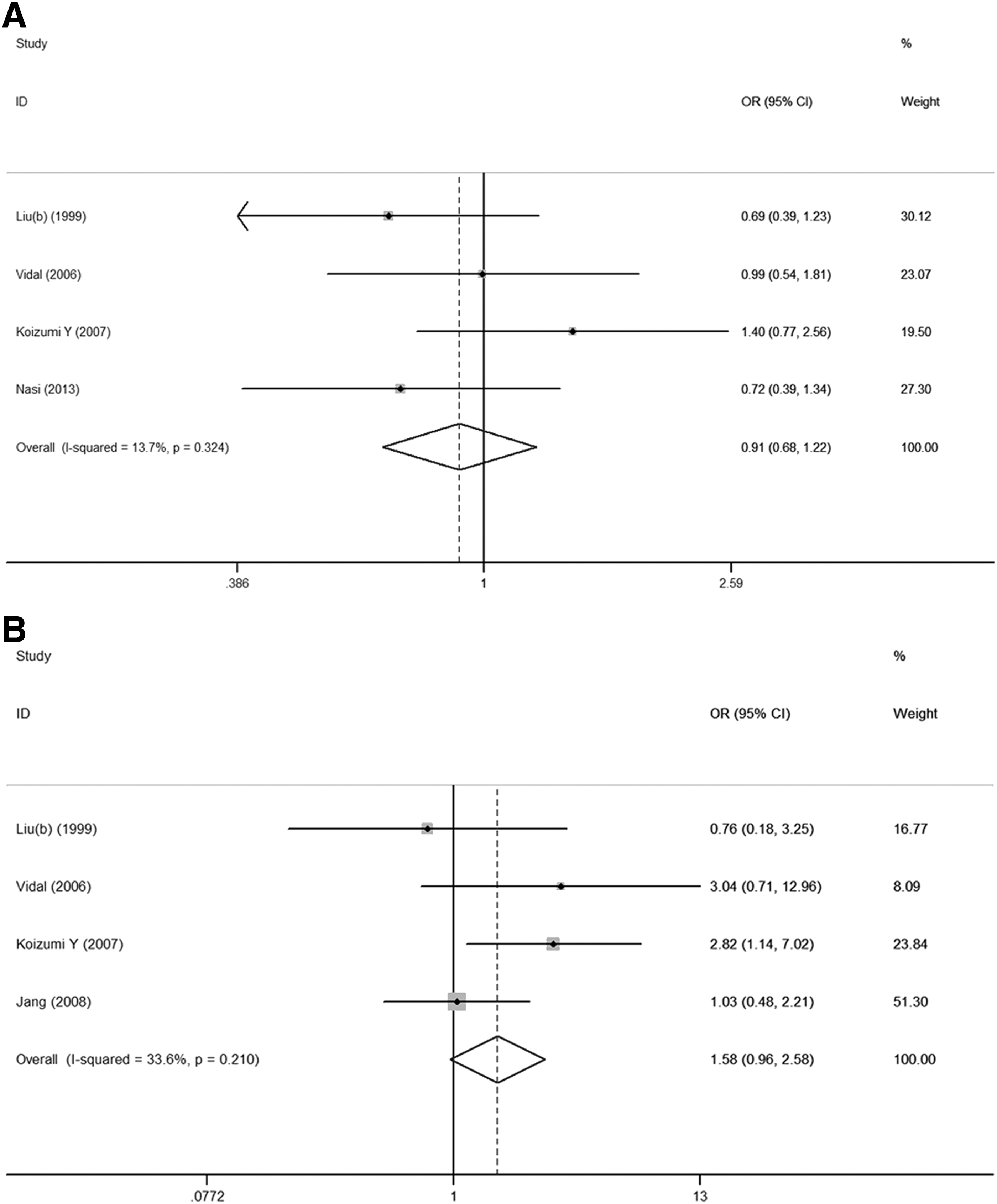
Meta-analysis of the association between the RANTES polymorphisms and progression of HIV-1 disease in allele model.
Heterogeneity analysis
For the −403G/A polymorphism, we found that all genetic models had p < .10 or I2 > 50% in overall populations, indicating statistically significant heterogeneity among the studies. The random-effects (DerSimonian–Laird) model (R) was used to calculate the data (Table 2). However, we chose the fixed-effects (Mantel–Haenszel) model (F) because of the absence of heterogeneity for −28C/G and In1.1T/C polymorphisms (Table 2). Logistic meta-regression analyses revealed that HWE may be the major source of the heterogeneity for the −403G/A polymorphism (allele model A vs. G: regression coefficient = −0.427, 95% CI = −0.845, −0.009, p = .046). The ethnicity, source of controls, and genotyping methods were not sources of heterogeneity (all p > .05 for meta-regression tests). Subsequently, subgroup analyses also show that a lack of heterogeneity was found only in the HWE (No) group in all genetic models.
Sensitivity analyses and publication bias
Sensitivity analyses were performed by deleting one study at a time and revaluating the ORs and 95% CIs. None of the studies had a significant effect on the final results of this meta-analysis (data not shown), indicating that our results are statistically robust. Although the genotype distribution in two studies for −403G/A polymorphism and in two studies for In1.1T/C polymorphism was not in accordance with HWE, the corresponding pooled ORs were not qualitatively altered when these studies were excluded. Subsequently, the Begg's funnel plot and Egger's test were conducted to further strengthen the confidence level in the results. No asymmetry was apparent in the funnel plots (Fig. 5), and statistical evidence from Egger's test in overall populations showed no significant publication bias in the studies (all p > .05 for Egger's test).
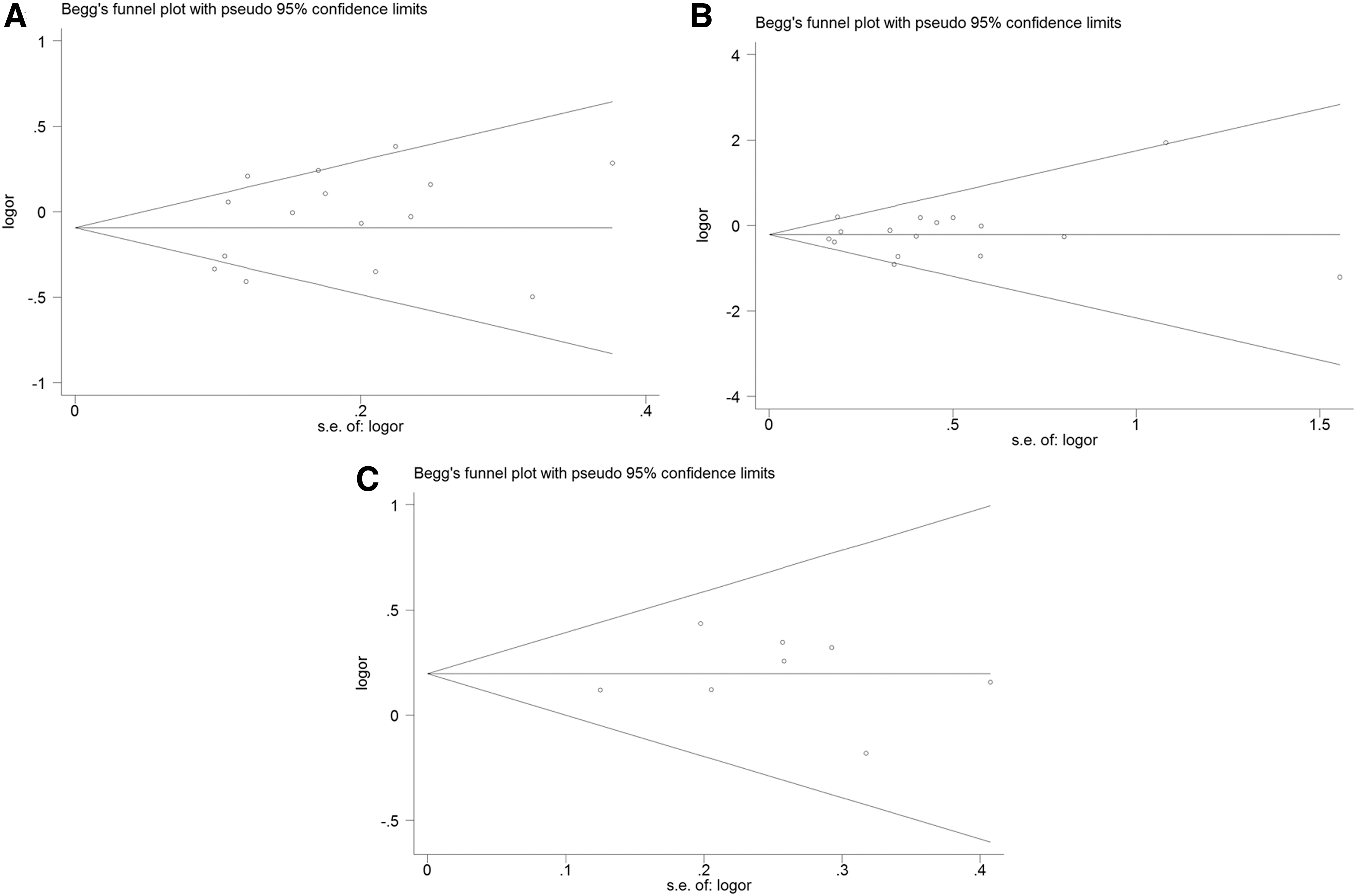
Begg's funnel plot and the Egger's test of publication bias.
Discussion
HIV/AIDS remains one of the world's most significant public health challenges. Till today, increasing numbers of studies focusing on the mechanism and pathogenesis of HIV/AIDS have been conducted to explore the effective control strategies against HIV/AIDS. These studies found that both viral genetics and host genetic background are significant determinants of HIV-1 infection and the progression of HIV-1disease. 46 –48 The RANTES-CCR5 pathway has been found to influence immune responses in multiple ways, 49,50 and it therefore plays an important role on risk of HIV-1 infection. When this pathway is blocked, the development and/or quality of antiviral immune responses are altered during chronic viral infection. 51 Moreover, therapeutic strategies targeting RANTES and CCR5 are being used for treatment against HIV infection. 52 Thus, SNPs in the encoding RANTES gene may be involved in susceptibility to HIV-1 infection and disease progression. In 1999, Liu et al. first examined the association between the RANTES −403G/A and −28C/G polymorphisms and HIV/AIDS risk in Japanese individuals. 15 In 2003, Liu et al. also examined the association between In1.1T/C polymorphism and HIV/AIDS risk in Chinese individuals. 21 Results of subsequent case–control studies conducted in different ethnicities were conflicting. 17 –20,22 –35 Therefore, the comprehensive meta-analysis can help elucidate the association between the RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms and susceptibility to HIV-1 infection and disease progression. This analysis was based on stricter search criteria and included a quality assessment of the studies selected, resulting in the most reliable and accurate results.
In the present meta-analysis, the results showed a decreased risk of HIV infection associated with the RANTES −403G/A polymorphism under the recessive genetic model (A/A vs. G/A+G/G), the pooled OR was 0.755 (95% CI = 0.581–0.912), especially among Asians (OR = 0.663, 95% CI = 0.533–0.836). However, no association was found among Caucasians (all p > .05 for all genetic models). In addition, our data also showed that the RANTES −28C/G polymorphism was associated with significantly decreased risk of HIV-1 infection, especially among Asians, but not among Caucasians (Table 2). Conversely, with respect to In1.1T/C polymorphism, a significantly increased risk of HIV-1 infection was found among Asians, but not among Caucasians. First, the most probable explanation is that both promoter polymorphisms (−403A and −28G alleles) can increase promoter activity, thereby upregulating the expression of RANTES protein, which may cause an increment in RANTES-CCR5 binding, which also contributes to reduce the depletion rate of CD4+ T cells, 15,16 ultimately inhibiting HIV-1 cell entry and replication. The RANTES In1.1C allele downregulates RANTES transcriptional activity, which shows a threefold reduction in gene expression relative to In1.1T and is probably associated with an increased risk for HIV-1 infection. 14 Second, the same gene polymorphism often plays different roles in influencing susceptibility to HIV-1 infection. It is possible that ethnic differences in genetic backgrounds and the environment may account for these differences. There have only been two studies of the association between RANTES gene polymorphisms with the risk of HIV-1 infection in African populations. 19 Additional studies are required to further validate the association of the RANTES −403G/A, −28C/G, and In1.1T/C polymorphisms with the risk of HIV-1 infection among Africans.
However, we found no correlation between the condition of LTNP and the −403G/A and −28C/G polymorphisms of the RANTES gene. This is most likely due to the small sample size that the only five eligible studies included, involving a total of only 253 LTNP cases and 403 UP controls. Additional studies are also needed to further validate this association.
The highly HESN individuals have been defined by Young et al., 53 and include the following groups: (a) commercial sex workers, (b) intravenous drug users, (c) discordant couples, (d) people with hemophilia, and (f) HIV-exposed but uninfected children. It seems that using HESN controls is more powerful than using the healthy controls. Subsequently, by using the HESN controls, the results showed no significant association between the RANTES −403G/A and −28C/G polymorphisms and the risk of HIV-1 infection. This may be due to the small sample sizes of the eligible studies and the HESNs in these studies always including subpopulations, such as intravenous drug users. More studies are required in the future on different ethnicities to explain the natural HIV-1 resistance in HESNs.
Moreover, we primarily found significant heterogeneity among the studies for the −403G/A polymorphism. Subsequently, logistic meta-regression and subgroup analyses revealed that HWE may be the major source of the heterogeneity, but the corresponding pooled ORs were not qualitatively altered when the studies in which the genotype distributions were not in accordance with HWE were excluded. Begg's funnel plot and Egger's test showed no significant publication bias in the studies. Thus, our results may be robust and reliable.
This meta-analysis should be interpreted within the context of some limitations. First, it may cover a small number of studies due to the fact that we only searched three electronic databases: PubMed, EMBASE, and the CNKI. An additional limitation was the exclusion of studies written in languages other than English and Chinese, which might bias our conclusions in this study. Second, the present meta-analysis was based on unadjusted estimates, and therefore, a more precise analysis of adjustment by other covariables (e.g., gender and environmental factors) could be performed if all individual raw data were available. Third, most studies were conducted in Asians and Caucasians. To avoid selection bias, further studies, including the full range of possible ethnic differences in genotype association studies, are required. Finally, gene–gene and gene–environment interactions may influence host susceptibility to HIV-1 infection and disease progression. In fact, many studies have proven that many other genes are related to risk of HIV-1 infection and disease progression, but we did not have enough data to eliminate these interfering factors. The socioeconomic status always influences the prevalence of HIV-1 infection and progression.
In conclusion, our meta-analysis involving more than 10,000 subjects shows that both the RANTES −403G/A and −28C/G polymorphisms are significantly associated with resistance to HIV-1 infection, whereas In1.1T/C polymorphism may increase the risk of HIV-1 infection, especially among Asian populations, but not among Caucasian populations. These results may contribute to finding a theoretical basis for effective control strategies against HIV/AIDS and enrich studies of gene polymorphisms in HIV/AIDS.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.