
Abstract
The effect of serial HIV-1 infection on the development of the broadly neutralizing antibody (bNAb) response was studied in an individual, H01-10366, with a serial HIV-1 superinfection (SI), hence triple infection, and compared with the bNAb response in three superinfected as well as 11 monoinfected men who have had sex with men (MSM) from Amsterdam, the Netherlands. Neutralization assays measuring heterologous neutralizing antibody (NAb) titers on a panel of six representative viruses from different HIV-1 subtypes were performed on blood serum samples obtained ∼3 years after primary HIV infection (PHI) and longitudinally for H01-10366. A bNAb response was defined as having a geometric mean neutralization titer (the reciprocal serum dilution giving 50% inhibition of virus infection, inhibitory dilution (ID50)) ≥100 and neutralizing >50% of viruses in the panel with an ID50 titer ≥100. H01-10366 quickly developed a potent NAb response against subtype B viruses before subtype B SI, but no broadening of the response occurred after the second subtype B infection or the third infection with CRF01_AE. When comparing H01-10366 with matched monoinfected (N = 11) and superinfected (N = 3) individuals analyzed 3 years after PHI, we found that 5 of the 15 individuals (4/11 monoinfected, 1/4 SI) developed a bNAb response. However, there was no statistically discernible difference between the bNAb response and HIV-1 SI. Thus, HIV-1 SI was not associated with the breadth and potency of the bNAb response in this small group of Dutch MSM with SI that included a triple HIV-1-infected individual.
Introduction
T
Molecular analysis of isolated potent monoclonal anti-Env bNAbs showed that they were all the result of long-term evolution and adaptation as exemplified by extensive somatic hypermutation and selection. This explains why the bNAb response takes a relatively long time to mature and often fails to develop in the first place. In mice engineered to express human immunoglobulin heavy-chain genes, immunization with a series of related Env immunogens was used to recreate the maturation of the bNAb response. 15 The limited cross-clade breadth observed here, suggested again that a repeated stimulation of the immune system is in all likelihood a necessity for the generation of a potent bNAb response.
Although a HIV-1 superinfection (SI) actually represents a failure of the immune system to protect from a second infection, there has been much interest in HIV-1 SI in relation to the development of bNAb. These serial infections can be seen as a natural booster of the immune response, where the already infected patient is challenged with a second virus strain carrying a divergent env gene. Indeed, broadening and strengthening of the NAb response were found in the Kenyan and Cameroonian cohorts of heterosexually infected women after intra- or intersubtype SI. 16 –18 In addition, a potent, high titer bNAb response peaking 3 years postinfection was described in a South African female who had experienced intrasubtype C SI 15 weeks after primary HIV infection (PHI). 19 However, SI was not associated with the development of bNAbs in a Tanzanian mixed subtype cohort. 20 Also, two men with intrasubtype B dual infection from the San Diego Primary Infection Cohort that contains mainly, but not exclusively, white men who have sex with men (MSM) as participants, were reported not to differ from monoinfected controls in the NAb response. 21
Not only the influence of SI but also the effects of gender and ethnicity upon the generation of NAb responses in HIV-1 infection have not yet been decisively resolved. In HIV-1 vaccine studies with healthy volunteers, it was reported that the NAb response is significantly stronger in women compared to men 22 and in African Americans compared to whites. 23 However, the findings might be different in HIV-1-infected persons, as HIV-1 infection severely affects the humoral immunity (reviewed by 24 ), Indeed, one study of mainly African American HIV-1-infected individuals showed that men had a higher bNAb activity compared to women. 25 In contrast, in another study, the breadth of neutralization was not associated with gender or ethnicity, 26 while a third found that NAb responses were significantly associated with the site of origin of the tested sample, through which European and Australian samples more often showed lack of neutralization and also a decreased breadth compared with African samples. 27 To complicate matters even more, differences in neutralization breadth were also found between two distinct African female cohorts from South Africa and Tanzania. 20
It is thus possible that the effects of HIV-1 SI upon the neutralizing activity are dependent upon gender and/or geographic origin of the patients or on the HIV-1 subtype that also often has a relationship with geography.
Although HIV-1 SI could be an adequate booster of the humoral immune system, it is likely that a single stimulation will not be sufficient in vaccine trials. Animal studies already suggest that repeated stimuli are needed. 15 Thus, individuals with serial SI could be more informative regarding the development of the NAb response. Although HIV-1 SI is not rare and is estimated to occur at rates that can be similar to incidence rates of primary infection, 28 –30 very few cases of serial SI are known. In 2005, we described a patient, H01-10366, with serial SI resulting in a triple HIV-1 infection. 31 We reasoned that it might be valuable to investigate the development of the NAb response in this individual in relation to the occurrence of SI. Therefore, we investigated the effect of serial HIV-1 infections on the breadth and potency (as indicated by serum NAb titers) of the heterologous NAb response in patient H01-10366. In addition, we incorporated 11 HIV-1 monoinfected individuals and three superinfected individuals from the same cohort of Dutch MSM as matched controls in the study. All monoinfections were with subtype B strains, while the HIV-1 SI cases consisted of one case with an intrasubtype B SI and two with intersubtype SI. The triple infected individual had been serially infected with two subtype B strains, followed by an infection with CRF01_AE.
Materials and Methods
Patient characteristics
The triple HIV-1-infected MSM H01-10366 (B/B/CRF01_AE) has been described before. 31 H01-10366 carries no protective HLA alleles 32 or the delta-32 deletion variant of the CCR5 gene (result not shown). The first SI occurred around 22 months after the estimated month of PHI, the second around 31 months after this moment. CD19+ B-cell counts were available for this individual from entry into care until month 37 after PHI. Three HIV-1 superinfected MSM with a PHI with subtype B and subsequent SI were identified in an earlier study. 33 Two had experienced intersubtype SI (SI2 B/CRF02_AG; SI3 B/CRF01_AE), while SI1 was superinfected with a second subtype B strain (B/B). SI occurred 6–12 months after PHI in these three individuals, and in all cases the initial and superinfecting virus strains persisted throughout the study period. NAb responses were measured at the latest time point after SI but before the initiation of antiretroviral therapy. Patient characteristics are summarized in Table 1.
The time point of 37 months was used in the group analysis, associated values are shown in bold.
MI, monoinfected; NA, not available; PVL, plasma viral load; SC, seroconversion; SI, superinfected.
A total of 11 HIV-1 subtype B monoinfected MSM with PHI were selected from the Amsterdam Cohort Studies on HIV infection and AIDS (
Sequencing of ∼300 nucleotide fragments encompassing the env V3–V4 region from the first and the follow-up sample used in this study of each control patient, as described previously, 34 revealed no HIV-1 SI during this period. The research protocol was approved by the Medical Ethics Committee of the Academic Medical Center, and all participants provided written informed consent.
All individuals were white, except one SI case with Asian ancestry. HIV-1 infections in the selected individuals occurred between 2001 and 2009, and all individuals were antiretroviral therapy naive during the study period.
Estimation of timing of the first HIV-1 infection in the triple infected patient
The triple HIV-1-infected patient H01-10366, who tested HIV-1 antibody positive in March 2001, was superinfected with a second subtype B strain in mid-2002 and with CRF01_AE in mid-2003.
31
To estimate the date of the first subtype B HIV-1 infection, ∼630 nucleotide env-gene fragments were amplified, cloned, and sequenced as described
35
from serial plasma samples obtained in 2001–2002 (antedating the first SI). Time to most recent common ancestor analysis was done with BEAST v1.6.2
36
using the GTR model with an invariant gamma-shape substitution model and empirical nucleotide frequencies under an uncorrelated relaxed clock and a coalescent tree prior assuming a constant population size, as the pVL remained low and unchanged at around 2,000 copies/ml. The Markov Chain Monte Carlo (MCMC) was run for 100.000.000 replications; the first 2,000 replications were discarded from the analysis (burn-in). From these data, it was estimated that PHI had occurred ±3 months before the first available sample (April 6, 2001), for example, in or around January 2001. Using a non-Bayesian method implemented in Path-O-Gen (tree.bio.ed.ac.uk/software/pathogen/)
37
to estimate the “best fitting root,” the position that is most compatible when assuming a molecular clock for trees that have been inferred without a molecular clock assumption, on the tip-dated maximum-likelihood tree obtained with MEGA5 (
Statistical analyses
Statistical analyses were performed in GraphPad Prism 5 (GraphPad Software, La Jolla, CA).
U87/Pseudovirus assay for testing of HIV-1 heterologous NAb in serum
Data on bNAb in serum for MSM were either available from our previous studies 38 or newly generated. In short, sera were tested in duplicate for bNAb in a single round viral infection assay involving six-tier two pseudoviruses and U87 cell lines that express either CCR5 or CXCR4, as developed and executed by Monogram Biosciences. 27 Previously, we have shown that classification of bNAbs in patient serum samples, as determined on an independent 23 viral panel or on the smaller six viral panel used in this study, was highly correlated. 39 Reactivity was measured at the latest time point after SI and before the initiation of antiretroviral therapy (median 36 months after PHI [range 28–44 months]) to exclude time of infection as factor for the development of a broad neutralizing response. For the triple infected patient, the time point estimated at 37 months post-PHI was used in the group analysis. Longitudinal samples from this patient were also tested with this assay. The geometric mean inhibitory dilution (ID50) titer was calculated per patient across the six viral panels. Neutralization titers were considered positive if they were three times greater than the negative Abelson murine leukemia virus control. The lowest serum dilution used in the assay was 1:40. For calculation of ID50 values for viruses that were not inhibited by the 1:40 serum dilution, we assumed that 50% inhibition would have occurred at a 1:20 serum dilution. A broad neutralizing response was defined as having a GM ID50 titer ≥100 with neutralization of >50% of viruses in the panel, for example, neutralizing at least four out of six viruses with an ID50 titer ≥100. 27
Results
Longitudinal heterologous NAb responses in the triple infected patient
To investigate the effect of serial SI on the development of the NAb response, the triple infected individual was followed longitudinally. Relatively high NAb titers against the subtype B viral strains in the panel (JRCSF and 92BR020) were detected throughout follow-up, consistent with clade B infection (Fig. 1A). Moderately high NAb titers (ID50 > 500) against strain JRCSF were already present at an early time point, for example, at ∼3 months after PHI, and were increased (ID50 > 1000) 6 months later. Consistent low-level neutralization of one of the subtype C viruses (93IN905) from the panel was seen throughout follow-up, corresponding with an earlier observation that 93IN905 is relatively easy to neutralize. 27 After SI with recombinant CRF01_AE, a small transient increase in the NAb response with a virus from the same subclass was observed (Fig. 1A, virus 92TH021). A similar weak response was seen against the reference subtype A strain in the panel, with a transient increase after the third infection, while the second subtype C virus was not neutralized at all.
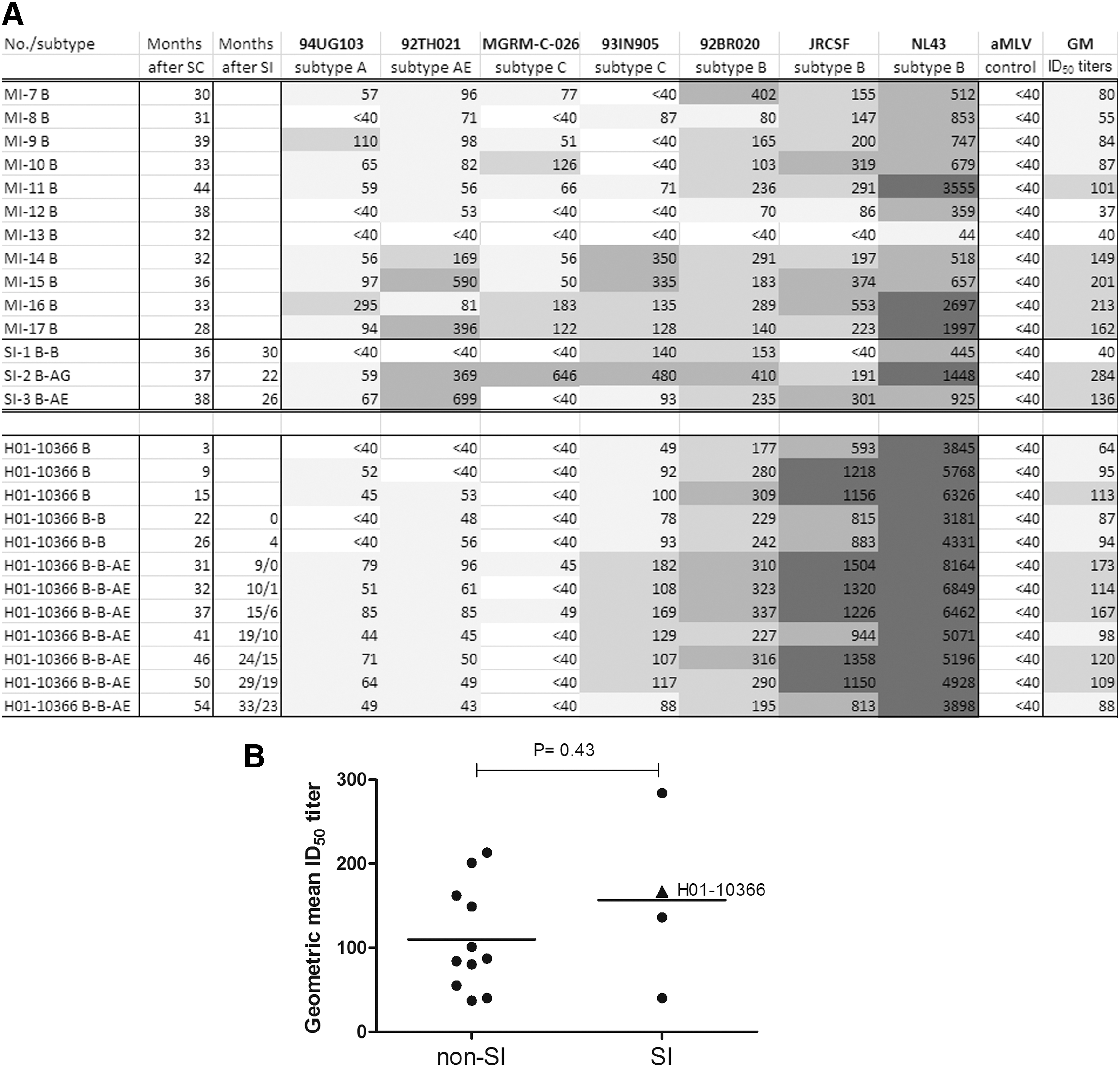
Broad neutralizing activity between individuals with mono- or serial HIV-1 infection.
So, after triple HIV-1 infection, no broadening of the NAb response was seen after the first SI with a virus from the same subtype, while intersubtype SI did lead to a minor increase in GM ID50 titers. A strong neutralizing response against the first infecting subtype B was already present within 1 year after PHI. This anti-subtype B response is stimulated but not significantly broadened after each SI, even after SI with a distinct subtype, but did lead to a small increase in NAb response to other subtypes.
Longitudinal B-cell dynamics and NAb titers in the triple infected patient
B cells, the cell type producing antibodies, are known to be severely affected by HIV-1 infection (for a review see 24 ). Elevated levels of B-cell apoptosis, and thus decreased B-cell counts, were seen in all HIV-1-infected individuals. 40,41 Higher rates of B-cell depletion in patients correlated with poor neutralizing activity against HIV-1 in both an European and an African cohort. 40,41 Longitudinal CD19+ B-cell counts of patient H01-10366 were plotted in relation to the GM ID50 titers to investigate their behavior in this specific individual (Fig. 2). GM ID50 titers increase after each SI, but finally decrease below 100 at 50 months. The CD19+ B-cell counts show a delayed but sharp increase followed by a steep decline after each SI. B-cell numbers first returned to the normal range (between 100 and 500 cells/ml for healthy Dutch adults 42 ) after the first SI, but remained below normal after the second SI. For healthy white males from Germany, a neighboring country, an age-related decline in CD19+ B-cell counts has been reported, through which the typical range for males <50 years of age would be 140–600 cells/ml blood. 43 Using such a modified age-dependent range would imply that patient H01-10366, who was well below 40 years of age at the study endpoint, has significantly decreased peripheral B-cell numbers at almost every time point. Irrespective of the reference range, however, it is clear that circulating CD19+ B-cell numbers decline after each HIV-1 infection, while the specific anti-HIV response lags behind. Stimulation and differentiating of resting memory cells to antibody-secreting plasma cells residing mostly in the bone marrow probably explain the delay in anti-HIV response.
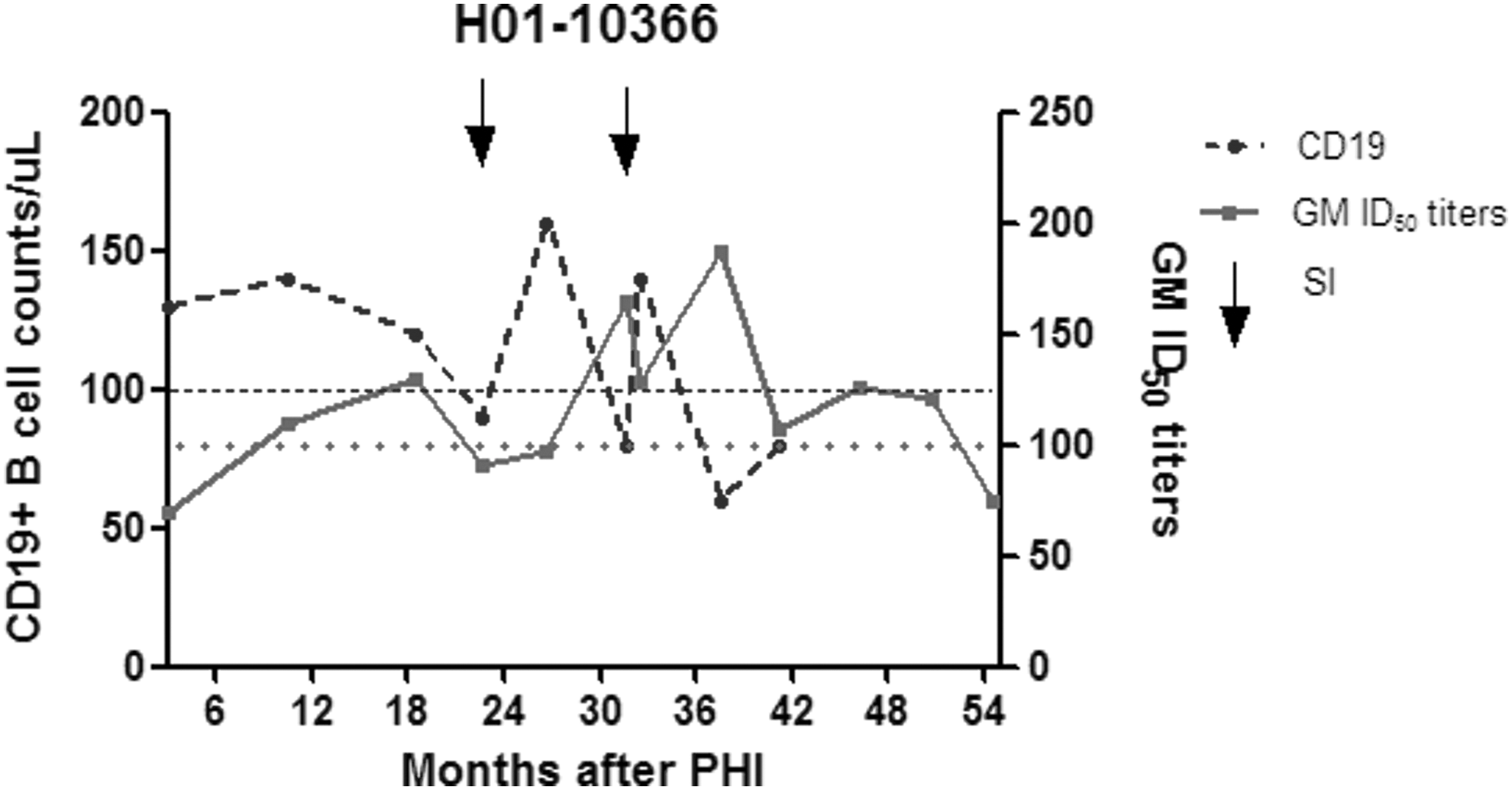
Longitudinal CD19+ B-cell counts and GM ID50 titers in patient H01-10366. Longitudinal CD19+ B-cell counts and heterologous neutralizing activity in patient H01-10366 with two HIV-1 SIs (indicated by arrows). Total CD19+ B-cell counts in peripheral blood over a course of 41 months of follow-up are shown. Reference values for CD19+ B cells in healthy Dutch adults are 100–500 cells/ml 43 (lower limit indicated by a dashed line). The GM ID50 titers during 54 months of follow-up are shown. GM ID50 titers are considered significant when ≥100 (indicated by a dotted line).
Heterologous NAb responses in Dutch MSM with and without SI
To compare the heterologous NAb response seen in patient H01-10366 with the development of the NAb response in a larger group of Dutch MSM, single time point samples from three individuals with HIV-1 SI and 11 time-matched individuals with mono-HIV-1 infection from contemporary Dutch MSM infected with mainly subtype B viruses were also tested for bNAbs across the six-virus panel. The 37-month follow-up sample from H01-10366 was used in the comparison (Fig. 1A). There were no statistical differences in the number of viruses neutralized (Fig. 1A) or the GM ID50 titers (Fig. 1B) between the monoinfected and SI cases, including the triple infected case (Mann–Whitney test, p = .43). More specifically, 4 of the 11 (36%) monoinfected (MI-14, MI-15, MI-16, and MI-17) and 1 of 4 (25%) superinfected (SI-2) individuals, but not H01-10366, developed a bNAb response after 3 years. Further analysis showed that no single parameter, such as age at infection, pVL at baseline, CD4+ T-cell counts at baseline or sampling, was significantly different between the five individuals who developed bNAb and the group that did not show such a response (result not shown). Only pVL at sampling was borderline significant (Student t-test, two tailed, p = .049) between the groups, but the implication of this finding is unclear and a larger group should be analyzed to confirm such an association. An earlier study in this MSM cohort indicated a significant association between CD4+ T-cell counts at baseline with the development of bNAb, 27,39 but this finding could not be corroborated in the present study, possibly because our patients were selected based upon their pVL and CD4+ T-cell counts.
Interestingly, an enhanced bNAb response was seen in the two individuals with an intersubtype SI compared with the patient with an intrasubtype SI (Fig. 1A). However, in the triple infected individual, an intersubtype SI only marginally and transiently increased the bNAb response.
Discussion
The NAb response against HIV-1 is highly variable, but most individuals appear capable of generating at least low to moderate NAb levels during chronic infection. 44 Time since PHI is an important parameter in predicting NAb levels, as the NAb response is slow to mature; viral replication and evolution appear to be essential during the process (see 1,7 ). Boosting the humoral immune system once or serially with a divergent HIV-1, as has been proposed for future vaccination schemes, could likely increase and broaden the NAb response. 15 As HIV-1 SI constitutes a natural booster and a second SI would likely enhance the response even more, we examined the breadth and potency of the NAb response in a HIV-1-infected individual with two subsequent HIV-1 SIs and three HIV-1-infected MSM with a single SI and compared the response to 11 matched HIV-1-infected MSM in whom SI was not detected. As the time since infection is a major contributor to the development of the NAb response, we only examined samples from individuals who were either seroconverters or presented with PHI. For the triple infected patient, we estimated the time of PHI through sequence analysis and demonstrated that he was in all probability infected with the first virus shortly before entering the study.
When the triple infected patient H01-10366 was tested for the heterologous neutralizing activity, a very fast and potent response against the subtype B virus JRCSF and a somewhat lesser response to another subtype B virus 92BR020 were seen. Cross neutralization of other subtypes in the test panel, for example, subtypes A, C, or CRF01_AE, was lower during the whole study than to subtype B strains and did not increase after an intrasubtype B SI. However, there was a small but transient increase to cross subtypes after the intrasubtype SI, implying that the neutralizing response was at best marginally and transiently broadened after stimulation of the immune system with a third divergent virus. This subdued response is surprising because infection with CRF01_AE in 2003 was accompanied by a sharp increase in the pVL to 106 copies/ml that could be attributed to the incoming virus 31 and CRF01_AE persisted and replicated to intermediate levels for almost 2 years. 32 The weak heterologous reaction to CRF01_AE could be due to the chronic immunodeficiency caused by more than 30 months of HIV-1 infection, with CD4+ T cells being relatively low at 310 cells/μl at the moment of the second SI. 31 However, there was no general deficiency in humoral activity, as NAb titers to subtype B viruses remained high over time and were even increased after the second SI.
Total B-cell counts in blood increased sharply after every SI and then decreased to or below threshold value. A reduction in peripheral total B cells is more pronounced during PHI than during chronic HIV-1 infection, 45 suggesting that SI is similar here to PHI in reducing B-cell numbers. The initial rise in peripheral blood B-cell numbers could likely be associated with a rise in plasmablasts that is related to the SI-induced HIV-1 viremia. 46 There is no noticeable relationship between the occurrence of SI, peripheral B-cell counts, and the concentration of bNAb in serum in patient H01-10366, although after each SI, the GM ID50 values peak, although on a different timescale. Recently, not peripheral plasmablasts but HIV-1 Env-specific plasma cells in the bone marrow have been associated with HIV-specific circulating antibody concentrations, 46 which may explain why no clear relationship between B-cell counts and bNAb titers is seen in H01-10366.
An inverse relationship between declining CD19+ peripheral B cells and elevated mean NAb titers against the infecting subtype was observed in an African HIV-1-infected cohort, 41 suggesting that low circulating B-cell numbers do not preclude an NAb response. Indeed, low B-cell numbers did not prohibit an NAb response in H01-10366. In this study, the highest titers were always directed against the initial infecting subtype. There was no significant broadening of the NAb response according to our criteria, suggesting that ongoing stimulation of the originally induced B-cell lineage(s) probably resulted in enhancement of the initial antibody response. An inability to induce new B-cell lineages would be suggestive of “repertoire freeze,” clonal dominance after a restricted B-cell response, a common occurrence in HIV-1 infection 47 This concept, termed “original antigenic sin,” was first coined in 1953 by Francis et al. 48 and has subsequently been postulated for many different pathogens, including HIV-1. 49
Relatively high titer NAbs against subtype B viruses were elicited by H01-10366 at the early time point of ∼9 months post-PHI, but this response did not protect from intrasubtype SI. It has been reported that the quality of the heterologous NAb response is not correlated with the occurrence of HIV-1 SI, 50 although often an absent or narrow autologous or heterologous response is seen before SI. 51 –53
When analyzing all MSM in this study together, no significant differences in NAb breadth and potency associated with serial HIV-1 infection were observed, in contrast to a report on African women that had also studied individuals with PHI before and after SI. 16 It is possible that there is a dissimilarity between men and women in the robustness of the NAb response, but there is also some other variation between the studies apart from gender, ethnicity, subtype, and mode of infection. In the study by Cortez et al., 16 the post-SI time point analyzed was much later (∼5 years after PHI) than in our study (≤3 years after PHI). NAb responses have been shown to increase at least up to 4 years after PHI, 6,54 although we have reported early bNAb responses in individuals with elite neutralizing ability. 10,55 Increased time of follow-up could possibly enable the amplification and detection of initially subtle differences. In contrast, the observation that we observed no significant differences in bNAb levels even after 3 years of infection indicates that SI does not in general boost or quicken the broadly neutralizing response. Although the incidence of SI can be as high as that of PHI (reviewed in 56 ), suggesting that host factors besides exposure are not important in acquiring SI, it has been noted that in SI cases the primary virus is often replicating at a low level. 33 Consequently, this virus might not induce an adequate cross-reactive NAb response thereby facilitating the SI with a different, better replicating strain. In line with our findings, two individuals with intrasubtype B dual infection from the San Diego Primary Infection Cohort that contains mainly, but not exclusively, white MSM as participants were reported not to differ from monoinfected controls in the NAb response. 21
Besides increasing with declining B-cell counts, the NAb response has been shown to increase also with declining CD4+ T-cell counts. 6,39 As the CD4+ T-cell count was similar here at baseline and at sampling in the single and SI infected individuals, the absence of an SI effect on the NAb response is probably not due to a difference in the number of CD4+ T cells. As CD4+ T-cell counts and pVL are correlated, pVL could also be an important factor inducing the NAb response. Indeed, in a recent study in two African HIV-1-infected female cohorts analyzed 2 years after PHI, the strongest predictor of the development of NAb response was found to be the initial pVL. Dual infection was not among the predicting factors, although the only elite neutralizer identified in this study was again a patient with HIV-1 SI. 20 However, in our study, the baseline pVL did not differ between the mono- and superinfected groups, and also was not different retrospectively between those that did or did not develop a bNAb response. The pVL at sampling was marginally associated with the development of a bNAb response, in line with many earlier studies that find such an association. 4 –6,26 However, low numbers preclude the detection of a robust association in the present study.
Another explanation for the failure of SI to stimulate the NAb response is that the SI virus is too divergent from the original infecting virus and therefore not naturally guiding the B-cell response initiated by the first virus to a matured bNAb response. Viral evolution through escape from NAb pressure has been postulated to be the most important mechanism in shaping the development of the HIV-1 neutralizing breadth (reviewed in 8 ). Therefore, in future vaccine schemes, it is probably essential to vaccinate serially with more advanced viral Env proteins, for example, a protein with longer variable regions and increased numbers of potential asparagine (N)-linked glycosylation sites, to guide B-cell reactivity. 1 In SI cases, the second infecting virus is again a primary variant, although different in sequence, and it might not be able to trigger the right B-cell lineages. A recent analysis of the characteristics of the superinfecting virus in three HIV-1 subtype C-infected women showed that SI was established by a “single CCR5-using founder virus with shorter, less glycosylated variable regions than matched chronic viruses”. 57 SI then indeed occurs with a virus type similar to PHI, although it might differ substantially in the primary amino acid sequence, which might explain its perceived failure to broaden and stimulate bNAb responses.
In conclusion, HIV-1 SI does not significantly enhance the heterologous NAb response in a Dutch cohort of MSM that included an individual with triple HIV-1 infection.
Footnotes
Acknowledgments
The study was financially supported by a Vici grant to H.S. from the Netherlands Organization for Scientific Research (grant no. 918.66.628), a grant from the European Community's Sixth Framework Program “European HIV Enterprise (EUROPRISE)” (FP6/2007–2012), under grant agreement 037611, and the European Community's Seventh Framework Program “Next-Generation HIV-1 Immunogens Inducing Broadly Reactive Neutralizing Antibodies (NGIN)” (FP7/2007–2013), under grant agreement 201433. M.J.v.G. was supported by a grant from the Aids Fonds Netherlands (grant No. 2012041). R.W.S. is a recipient of a Vidi grant from The Netherlands Organization for Scientific Research (NWO) and a Starting Investigator grant from the European Research Council (ERC-StG-2011-280829-SHEV). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
The Amsterdam Cohort Studies on HIV infection and AIDS, a collaboration between the Public Health Service of Amsterdam, the Academic Medical Center of the University of Amsterdam, Sanquin Blood Supply Foundation, Medical Center Jan van Goyen, and the HIV Focus Center of the DC-Clinics, are part of the Netherlands HIV Monitoring Foundation and financially supported by the Center for Infectious Disease Control of the Netherlands National Institute for Public Health and the Environment.
The authors thank Evelien Bunnik for sharing data.
Author Disclosure Statement
No competing financial interests exist.