
Abstract
The development of immunodeficiency in HIV-infected patients is known to result from CD4+ lymphocyte depletion. Most CD4+ lymphocyte cells destined to die are not infected. The mechanism of HIV-uninfected cell death has not yet been fully elucidated. The aim of this study is to examine the role of anti-CD4 autoantibodies and physiological rheumatoid factor (RF) in the development of CD4+ lymphocytopenia. Immunization of Wistar rats with gp120 HIV-1 induces chronic production of anti-CD4 autoantibodies and decreases CD4+ lymphocytes in the blood. However, the anti-CD4 autoantibodies produced as part of the immune response to gp120 do not kill CD4+ cells directly. In rats producing anti-CD4 autoantibodies, a low level of peripheral CD4 lymphocytes is associated with high blood RF levels. The sera containing RF killed lymphocytes when the lymphocytes were pretreated with sera containing anti-CD4 autoantibodies. Thus, the death of CD4+ lymphocytes in rats immunized with gp120 is a result of the combined action of anti-CD4 autoantibodies and RF, and the action of these factors can be separated in time. The fact that two signals are needed for CD4+ lymphocyte death in HIV gp120-immunized rats does not contradict the hypothesis of the activation-induced death of uninfected CD4+ cells in HIV-infected humans.
Introduction
T
The mechanism of death of HIV-uninfected cells that results in immune deficiency has not yet been fully elucidated. Several hypotheses were offered. The role of apoptogenic HIV proteins and antibodies to gp120 that induce cross-linking of gp120 on CD4+ lymphocyte membrane has been considered. 1 –3 The counter-argument to hypothesis that consider viral proteins, independently or together with anti-gp120 antibodies, to be the cause of CD4 T lymphocyte death is that such theories do not explain the lack of correlation between viral load and blood levels of CD4 T cells in HIV-infected patients nor the AIDS-like symptoms without HIV infection that are observed, for instance, in idiopathic CD4 T lymphocytopenia and mixed connective tissue disease. 4 –8
The hypothesis of activation-induced cell death of uninfected CD4 T lymphocytes appears less contradictory. 9 Under this hypothesis, in HIV infection, uninfected CD4 T lymphocytes undergo a generalized activation that sensitizes the CD4 T cells to apoptosis. Subsequently, when the activated cells encounter an apoptosis inducer, they undergo apoptosis. However, neither the factor that induces this generalized activation of uninfected CD4 T lymphocytes nor the apoptosis inducer, that is, the factor that kills the activated CD4 T lymphocytes, is yet known.
Anti-CD4 autoantibodies can be the factor that causes the chronic polyclonal activation of uninfected CD4 cells in HIV infection and sensitizes them to apoptosis. 10 The hypothesis that the autoimmune response to CD4 plays an important role in AIDS pathogenesis is not a new idea. 11 Depletion of CD4+ T cells in HIV- or simian immunodeficiency virus-infected individuals was associated with plasma antibodies autoreactive with CD4+ T cells. 12 –15 Furthermore, Corre et al. demonstrated the presence of idiotype-anti-idiotype interactions between anti-gp120 antibodies and autoantibodies to CD4 in HIV-infected people, and the mechanism by which the autoantibodies to CD4 form during HIV infection is clear. 16 However, the autoimmune hypothesis of AIDS is not generally accepted. Perhaps the reason is that none of the studies conducted have clearly identified the mechanism by which anti-CD4 autoantibodies could mediate CD4+ T-cell depletion.
The aim of this study is to examine the role of anti-CD4 autoantibodies induced by gp120 HIV-1 in the development of CD4+ lymphocytopenia. The model chosen for study was immunization with purified gp120 protein from HIV, since under the autoimmune theory of AIDS, the immune response to gp120 serves as the cause of anti-CD4 autoantibody induction. 16 This model precludes a contribution by the virus or its apoptogenic proteins to the depletion of CD4 T lymphocytes.
Humanized mice are the most appropriate for inducing an autoimmune response to CD4. However, in the pilot version we tried using Wistar rats. The basis for this attempt was the homology between rat CD4 and human CD4 in the area responsible for binding with HIV-1 gp120. 17 The homology suggests that HIV-1 gp120 may be complementary to rat CD4, and consequently that the anti-CD4 lymphocytes that may be activated by anti-gp120 lymphocytes through idiotype–anti-idiotype interactions will be specific to rat CD4.
Previously we showed that resistance to experimental autoimmune diseases was associated with the production of rheumatoid factor (RF), which was detected by the method of agglutination of tanned erythrocytes loaded with homologous IgG. 18 The role of RF in the formation of resistance to developing an autoimmune response to CD4 or regulating the immune response to gp120 has not been studied. Therefore, one of the aims was to investigate the role of RF in regulating the immune response to HIV gp120.
Materials and Methods
Rats
Wistar rats (8–10 weeks old) were used. They were brought from the Stolbovaya breeding facility (Russia) and housed under standard laboratory conditions with food and water at a constant temperature of 20 ± 5°C. Animal experiments were performed in accordance with European Commission Directive 86/609/EEC (European convention for the protection of vertebrate animals used for experimental and other scientific purposes) and approved by the local animal care commission of Udmurt State University.
gp120 HIV-1 immunization and blood collection
Recombinant GP120 HIV-1 glycoprotein (ACROBiosystems, Newark, DE) was emulsified with the same volume of incomplete Freund's adjuvant (IFA; Sigma, St Louis, MO). Rats (n = 7) were injected at the tail base with gp120/adjuvant emulsion (20 μg of gp120 per rat). No booster immunization was performed. Control rats (n = 8) received an injection of an emulsion containing saline and IFA. Blood sampling for anti-gp120 antibody, anti-CD4 autoantibody, and RF detection was carried out every week. From time to time, blood samples were used to determine CD4+ cell counts. The number of exposed rats and days of CD4+ cell count determinations are presented in Figure 2a.
Determination of anti-gp120 antibodies by enzyme-linked immunosorbent assay
Plates (Corning-Costar, Acton, MA) were coated overnight at 4°C with 50 μl of gp120 HIV-1 (ACROBiosystems) [10 μg/ml in 0.15 M phosphate-buffered saline (PBS)]. Plates were blocked with 150 μl 5% milk powder in PBS. Serum samples were added in a 1:200 dilution with PBS/Tween-20 and incubated for 1 h at room temperature (RT). The plates were then incubated for 1 h at RT with 100 μl of goat antirat Ig (IgG, IgM, IgA) conjugated to horseradish peroxidase (IMTECH, Moscow, Russia). Then the substrate mixture [5 ml of citrate-buffered solution (pH 5.0)/3 mg ortho-phenylenediamine/15 ml 3% H2O2] was added. Absorbances were read after 15 min at 492 nm.
Determination of anti-CD4 antibodies by enzyme-linked immunosorbent assay
Plates (Corning-Costar) were coated overnight at 4°C with 50 μl of recombinant rat CD4 (R&D Systems, Minneapolis, MN) (10 μg/ml in 0.15 M PBS). Plates were blocked with 150 μl 5% milk powder in PBS. Serum samples were added in a 1:100 dilution with PBS/Tween-20 and incubated for 1 h at RT. The plates were then incubated for 1 h at RT with 100 μl of goat antirat Ig (IgG, IgM, IgA) conjugated to horseradish peroxidase (IMTECH). Then the substrate mixture [5 ml of citrate-buffered solution (pH 5.0)/3 mg ortho-phenylenediamine/15 ml 3% H2O2] was added. Absorbances were read after 15 min at 492 nm.
Measurement of RF
The RF titer was determined in an agglutination test using rat-IgG-loaded tanned erythrocytes. Group 0 human erythrocytes were fixed with 1% glutaric dialdehyde. The washed-off fixed erythrocytes [10% in PBS (pH 7.2)] were incubated for 10 min with the same amount of tannin solution in PBS at RT. The erythrocytes were then washed off. For sensitization, 4 ml of PBS (pH 6.4), 1 ml of a solution of 0.5 mg/ml normal IgG from rat serum (Equitech-Bio, Kerrville, TX) in 0.9% NaCl, and 70 μl of the pelleted tanned erythrocytes were mixed. Incubation lasted for 20 min at RT; erythrocytes were washed with 0.9% PBS containing 0.2% bovine serum albumin. Twofold serial dilutions of serum were prepared and 50 μl of which was put into wells. The same amount of rat-IgG-loaded 1.5% erythrocyte suspension was then added. Agglutination results were read in 3 h.
Determination of CD4+ cell counts
The lymphocytes were isolated in ficoll–urografin gradient (ρ = 1.08 g/cm3). Then aliquots of cells washed in PBS were stained for 1 h at 37°C with FITC-conjugated anti-rat-CD4 (eBioscience, San Diego, CA), 0.25 mg antibody per 106 cells in a final volume of 100 μl. The lymphocytes were then washed off. After that CD4+ lymphocyte counts were determined using laser-based flow cytometry (BD FACSCanto II Flow Cytometer).
Viability of lymphocytes treated with anti-rat CD4 autoantibodies and RF in vitro
The lymphocytes were isolated in ficoll–urografin gradient (ρ = 1.08 g/cm3) from intact rats. The lymphocytes were washed off in RPMI-1640. After that, 100 μl of serum from gp120-immunized rats containing antirat CD4 antibody or 100 μl of serum from IFA-injected rats containing no antirat CD4 antibody was added to 106 lymphocytes in RPMI-1640 in a final volume of 1 ml. The RF titer of the sera that were added was ≤1:4. The lymphocytes were incubated for 24 h in a 37°C, 5% CO2 incubator. The lymphocytes were then washed off in RPMI-1640 and were suspended in a volume of 100 μl. Then 100 μl of serum containing RF (RF titer ≥1:32) and obtained from intact rats or 100 μl of serum containing no RF (RF titer ≤1:4) and obtained from intact rats was added. After incubation for 3 h in a 37°C, 5% CO2 incubator, the number of cells was counted using trypan blue.
Statistical analysis of the data
The significance of differences was assessed by t-test.
Results
Production of anti-CD4 autoantibodies and CD4+ lymphocytopenia in rats immunized with HIV 1 gp120
Immunization of Wistar rats (n = 7) with HIV-1 gp120 induced the production not only of antibodies to HIV-1 gp120 but also of autoantibodies to rat CD4 (Fig. 1). Anti-CD4 autoantibody production was revealed in all of the gp120-immunized rats. In rats receiving IFA (n = 8), no anti-CD4 autoantibodies were found (Fig. 1). Figure 1 shows that the production of anti-gp120 antibodies induced by a single injection of HIV-1 gp120 did not attenuate over the 4 months of observation. The production of anti-CD4 autoantibodies also was chronic in nature; the level of anti-CD4 autoantibodies in the blood of rats immunized with gp120 remained significantly higher than that of IFA-injected rats even 4 months postimmunization. In summary, immunization with HIV-1 gp120 induces chronic anti-CD4 autoantibody production in Wistar rats.

Anti-CD4 autoantibodies production and time course of anti-gp120 antibodies in rats immunized with gp120 HIV-1. Each point is represented as mean ± SD of 7 (gp120-immunized rats) and 8 (IFA-injected rats). IFA, incomplete Freund's adjuvant; OD, optical density; SD, standard deviation.
The count of CD4+ cells in the blood of the gp120-immunized rats and the IFA-injected rats is shown in Figure 2a. In rats that have developed an autoimmune response to CD4, the average number of CD4+ cells was significantly lower than the average number of CD4+ cells in the peripheral blood of IFA-injected rats (Fig. 2b). The lowest number of CD4+ cells induced in rats immunized with gp120 that developed an autoimmune response to CD4 was 293 cells per micro liter of blood (Fig. 2c, left picture). Representative flow cytometry of CD4+ cells in IFA-injected rats is shown in Figure 2c (right picture). The decrease in CD4+ lymphocytes in rats immunized with gp120 protein was transitory in nature; a drop was followed by a compensatory rebound in CD4 lymphocyte levels.
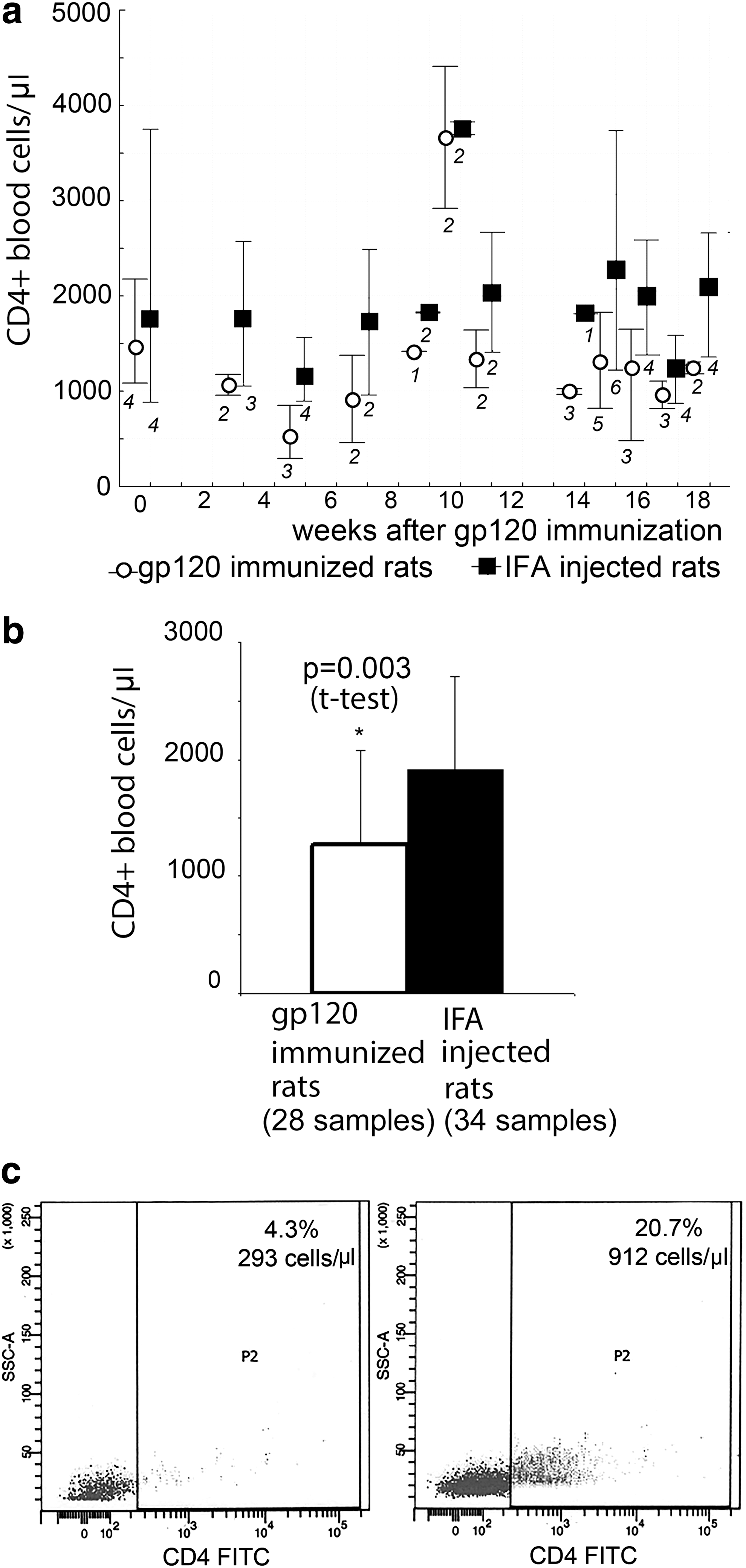
CD4+ cell counts in the blood of gp120 HIV-1-immunized and IFA-injected rats.
Contrary to expectations, no correlation was found between the level of anti-CD4 autoantibodies and the number of CD4+ lymphocytes in the blood (r = .24, p = .27). The drop in the number of CD4+ cells in the blood can be observed against a background of stable, relatively low anti-CD4 antibodies in the blood, and vice versa—when the level of anti-CD4 antibodies is increasing rapidly, the number of CD4 cells can remain stable as compared with the number of cells in IFA-injected rats. In sum, immunization of Wistar rats with HIV-1 gp120 induces chronic production of anti-CD4 autoantibodies and decreases CD4 lymphocytes in the blood. However, no correlation was found between antibodies to CD4 lymphocytes and the number of CD4 lymphocytes in the blood.
Role of RF in the development of CD4+ lymphocytopenia in rats immunized with HIV-1 gp120
The time course of RF in response to HIV-1 gp120 immunization and in IFA-injected rats is shown in Figure 3. RF levels were found not to increase in response to gp120 immunizations. The surges in RF production observed at 5, 9, 12, 15, and 17 weeks after gp120 immunization were delayed relative to immunization with the gp120 protein of HIV, whereas the kinetic curves of RF in gp120-immunized and IFA-injected rats almost entirely coincide (Fig. 3). Therefore, the observed changes in RF level in the blood of gp120-immunized rats are not the result of the immune response to gp120 nor the administration of animal IFA, but are spontaneous in nature and are natural variations in RF level. In sum, RF is not involved in downregulating the immune response to gp120 HIV in rats.
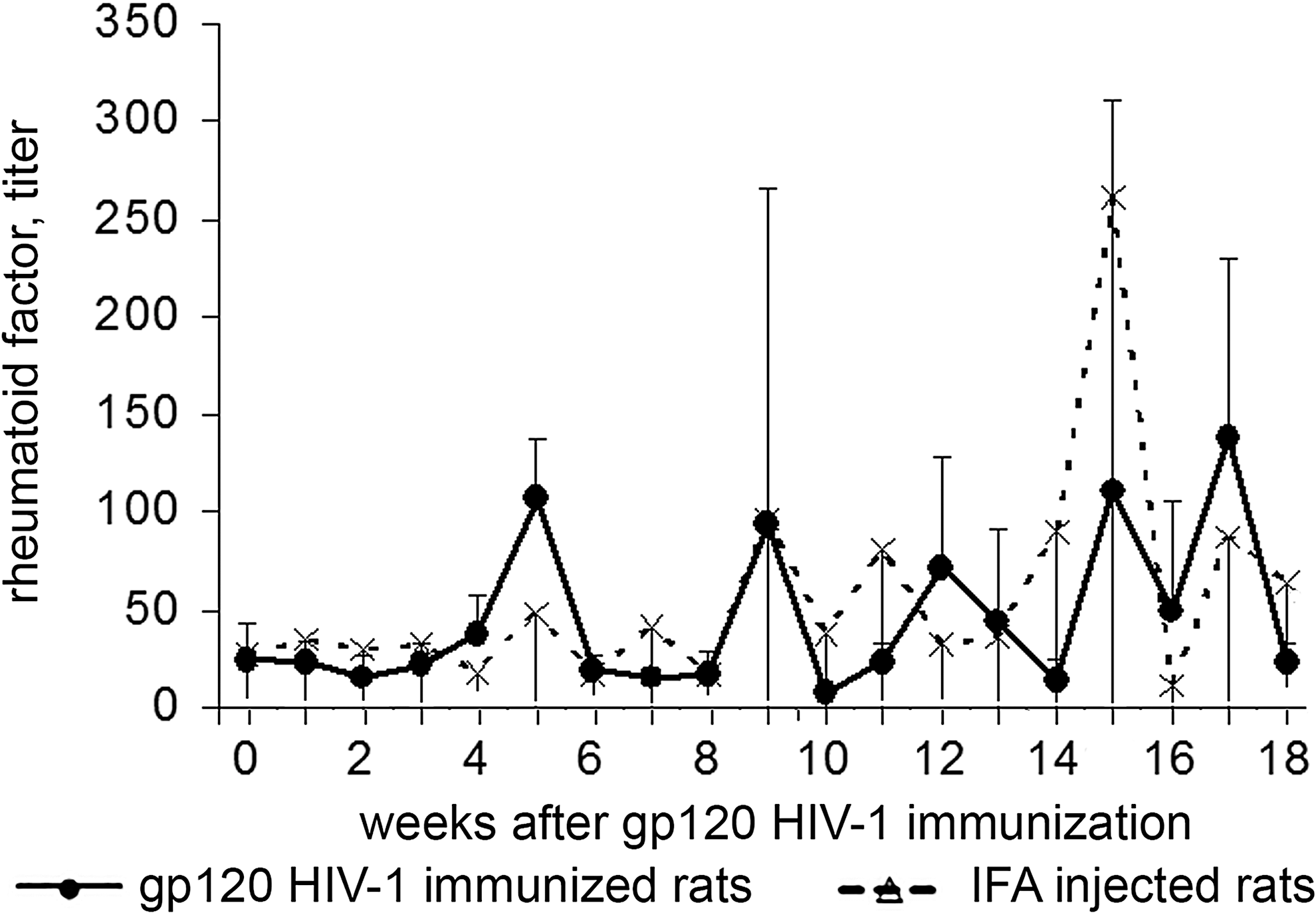
The time course of RF in rats in response to HIV-1 gp120 immunization. RF, rheumatoid factor.
In an individual analysis of the parameters studied, we noticed that in the rats immunized with HIV gp120, the minimum of the blood CD4 lymphocytes corresponded with the maximum of the blood RF levels (Fig. 4a). No such association was found in the rats that had received IFA in place of gp120.
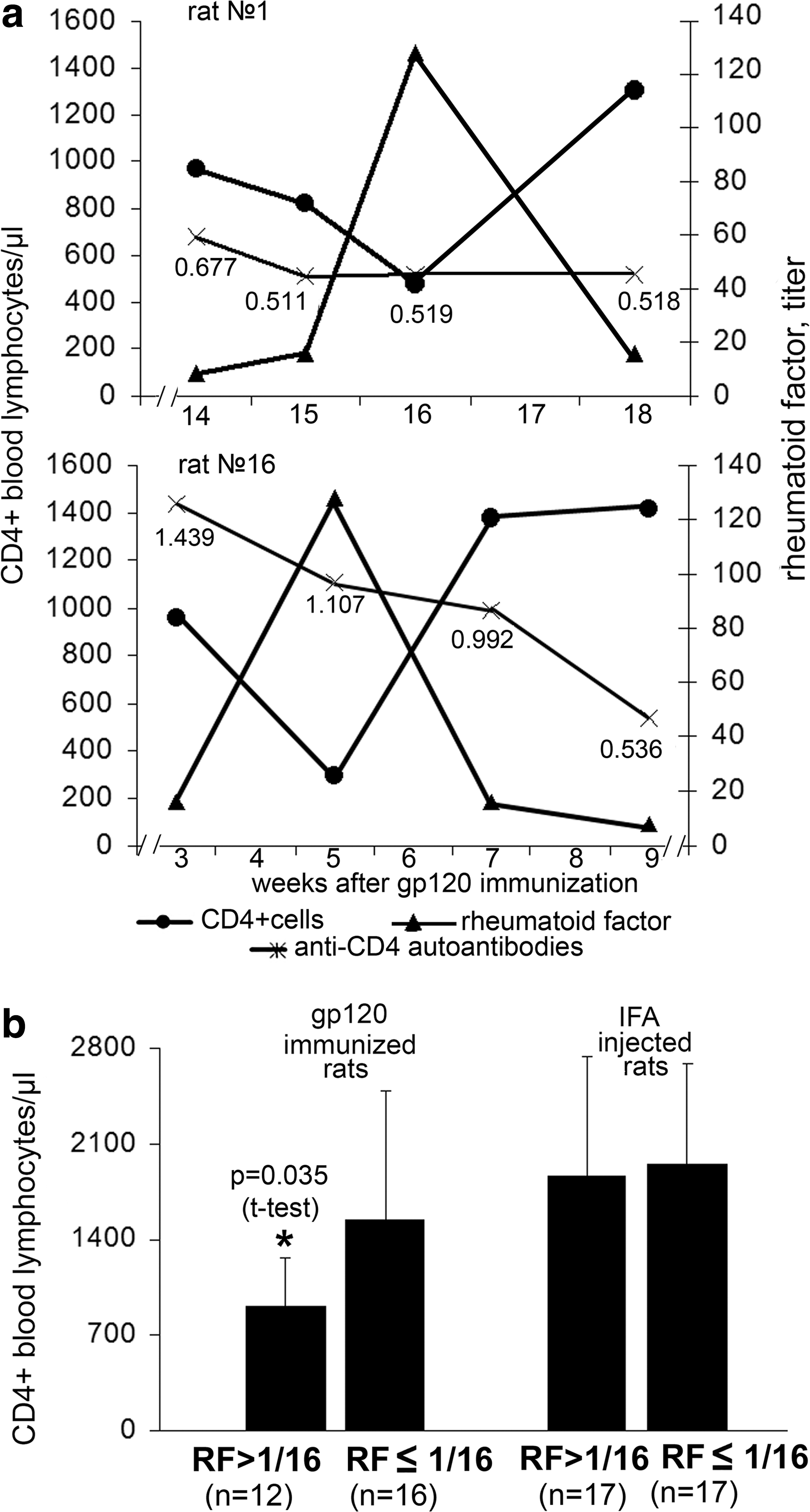
Number of CD4+ lymphocytes in the blood of rats with high and low titer of RF.
We compared the level of CD4 cells at a relatively high RF level (titer >1/16) and a relatively low RF level (titer ≤1/16) in the blood of rats immunized with gp120. The same test was performed in IFA-injected rats. In the gp120-immunized rats, when RF blood levels were relatively high, the number of CD4 lymphocytes was significantly lower than that in the rats with a relatively low RF level (Fig. 4b). In the IFA-injected rats, whether the blood RF was relatively high or relatively low, the blood CD4 cell levels were the same (Fig. 4b). In sum, in rats immunized with gp120 and producing anti-CD4 autoantibodies, a low level of peripheral CD4 lymphocytes is associated with high blood RF levels. In the IFA-injected rats, which have no anti-CD4 autoantibodies, the number of CD4 cells does not depend on blood RF levels. This information prompts us to suggest that the death of CD4 lymphocytes in rats immunized with gp120 is a result of the combined action of CD4 autoantibodies and RF, and the action of the factors can be separated in time.
Effect of serum containing anti-CD4 antibodies and serum containing RF on lymphocyte viability in vitro
To test the assumption that CD4+ lymphocyte death in rats immunized with gp120 results from the combined action of anti-CD4 autoantibodies and RF, we performed additional in vitro studies. Lymphocytes from intact rats (n = 11) were incubated with sera containing anti-CD4 antibodies and obtained from gp120-immunized rats (n = 11). Next, the lymphocytes were treated with serum from intact rats (n = 11) that contains RF.
We found that the lymphocytes pretreated with serum containing anti-CD4 antibodies died in response to the presentation of serum containing RF (Fig. 5). Lymphocytes did not die if serum without RF was added to lymphocytes incubated with serum containing anti-CD4 (Fig. 5). The lymphocytes also stayed alive when the lymphocytes were incubated in serum without anti-CD4 antibodies and then presented with RF-containing serum. Thus, anti-CD4 autoantibodies and RF do not induce lymphocyte death on their own; the death of CD4 lymphocytes results from the combined action of RF and anti-CD4 antibodies obtained from rats immunized with HIV-1 gp120.

Death of lymphocytes after incubation with or without sera containing anti-CD4 autoantibodies and treatment with RF-containing sera (mean ± SD).
Discussion
The aim of this study was to test the hypothesis that the cause of CD4 lymphocytopenia in HIV infection is an autoimmune response to CD4 induced by HIV-1 gp120. It is known that in HIV infection, lymphocytes specific to CD4 are activated by lymphocytes against HIV-1 gp120 through idiotype–anti-idiotype interactions. The ability of lymphocytes against HIV gp120 to enter into idiotype–anti-idiotype interactions with lymphocytes against human CD4 is prompted by the complementarity of the antigen-recognizing receptors of these lymphocytes, which itself is a consequence of the complementarity between the gp120 CD4-binding sequence and the first domain of human CD4.
In choosing rats as the subjects for testing the hypothesis, we assumed that rat lymphocytes, which can be activated by anti-gp120 lymphocytes through idiotype–anti-idiotype interactions in response to HIV-1 gp120 immunization, would not be specific to rat CD4. This doubt was supported by data on the homology of about 50% between the first domain of human CD4, which is responsible for HIV-1 gp120 binding, and the analogous section of the rat CD4 molecule. The fact that antibodies to rat CD4 appeared in the rats in response to gp120 immunization indicated that the rat lymphocytes specific to HIV gp120 enter into idiotype–anti-idiotype interactions with autoreactive lymphocytes against rat CD4 and that immunization of rats with HIV-1 gp120 can be used to test the hypothesis.
To measure anti-CD4 autoantibodies, recombinant rat CD4 was used. Some researchers are skeptical of the use of recombinant CD4 (rCD4) to measure anti-CD4 lymphocyte antibodies, because they believe that rCD4 differs from the membrane-bound molecule. 19,20 However, rCD4-specific antibodies can appear in rats in response to gp120 immunization only if the lymphocytes that produce antibodies specific to rCD4 enter into idiotype–anti-idiotype interactions with lymphocytes against gp120, and subsequently carry the internal image of gp120.
Considering that gp120 interacts with membrane-bound lymphocyte CD4, we can assert that the rCD4 preparation we used also bears sections responsible for gp120 binding, and consequently in this area it is the same as membrane-bound CD4, and the auto-antibodies revealed by rCD4 are specific to the available determinants on membrane-bound CD4. This assertion does not contradict the result obtained by Corre et al. that anti-idiotypic antibodies to human anti-gp120 antibodies bind recombinant and cellular human CD4. 16
According to the autoimmune hypothesis of AIDS, the rat anti-CD4 autoantibodies should induce death of rat CD4 T lymphocytes. In rats immunized with gp120 and producing anti-CD4 antibodies, the average levels of CD4 lymphocytes were significantly lower for the experiment than in the IFA-injected rats. However, the anti-CD4 autoantibodies produced as part of the immune response to gp120 do not kill cells directly. The lack of a correlation between anti-CD4 autoantibodies and the number of CD4 lymphocytes in the blood of gp120-immunized rats, as well as the results of in vitro experiments in which we tried to induce lymphocyte death in the presence of serum that contained CD4 autoantibodies obtained from gp120-immunized mice, serves as evidence of this.
The death of lymphocytes exposed to anti-CD4 autoantibodies induced by gp120 immunization commences in the presence of RF, both in vivo and in vitro. The fact that two signals, from anti-CD4 autoantibodies and RF, are needed for CD4 lymphocyte death in HIV gp120-immunized rats does not contradict the hypothesis of the activation-induced death of uninfected CD4+ cells in HIV-infected humans. This hypothesis also indicates that two factors are needed: a factor that activates the cells and a factor that kills the activated cells, and the action of the factors can be separated in time. Furthermore, our data coincide with results obtained by Susal et al., who found the association between a high anti-Fab/anti-rCD4 ratio and a low level of CD4+ helper T cells in AIDS patients. Anti-Fab antibodies and RF have similar characteristics. Both RF and anti-Fab antibodies possess anti-Fab activity and downregulate the immune response. 18,21
At the same time, our data raise new questions, particularly concerning the mechanism of action of RF that results in CD4 lymphocyte death.
We previously conducted detailed studies of the RF population, which is associated with the drop in blood CD4 lymphocytes in HIV gp120-immunized rats. This RF population is found in intact animals; therefore, it can be considered physiological. We showed that an increase in RF production in response to immunization with antigen inducers of experimental autoimmune diseases prevents the development of those diseases and is also associated with termination of the normal immune response. 18
A study of RF specificity found that every RF molecule has two kinds of paratopes: a unique individual paratope and a shared paratope; the specificity of the shared paratope is the same across the different RF molecules. The individual paratopes of RFs represent internal images of the foreign antigens; therefore, the RFs are anti-idiotypic antibodies, which recognize the antigen-binding sites of antibodies to foreign antigens. So the RF naturally occurring in the blood is heterogeneous in its specificity population of anti-idiotypic antibodies. 18 The shared paratope recognizes the recurrent sequence that exists on antibodies of various specificities and is located in the Fab region but outside the antigen-binding site. 18
Furthermore, we showed that the antigenic determinants for the shared RF paratope can be created in the Fc hinge region of IgG fragments through papain cleavage of IgG. 18 The specificity of the shared paratope of RF to the hinge region of IgG Fc fragments distinguishes the given RF from other known RF populations, which have determinants localized between the CH2 and CH3 domains. 22
If RF is classified in accordance with the list of types of anti-idiotypic antibodies proposed by Parsons et al., 23 the properties of the individual paratope of the RF molecule enable the RF to be classed as Ab2 Beta (Ab recognizes the antigen-binding site), whereas the properties of the shared paratope of RF resemble the properties of the Ab2 Delta class (Ab recognizes nonbinding site, nondisease specific, and outbred shared idiotope). Thus, RF is simultaneously both Ab2 Beta and Ab2 Delta.
Based on this information, we proposed a hypothesis explaining how RF prevents autoimmune response. 24 We think that RF normally acts as a factor depleting activated antigen-specific CD4 cells. In the course of an immune response to a foreign antigen (including antigens that induce experimental autoimmune diseases), RF is formed as anti-idiotypic antibodies; therefore, its target is only antigen-specific lymphocytes. RF does not recognize CD4 lymphocytes with other specificities. Transmission of the death signal to the activated antigen-specific CD4 lymphocytes may occur by virtue of the shared paratope on RF. The ligand for the shared paratope on the CD4 lymphocyte membrane may be an inducible molecule that is structurally similar to the Fc fragments of IgG and appears only in activated lymphocytes.
Anti-CD4 autoantibodies produced in response to gp120 immunization can induce polyclonal activation of peripheral CD4 lymphocytes and the appearance thereon of the ligand for the shared paratope of RF (Fig. 6). Therefore, the RF normally in the blood gets the opportunity to bind with CD4 lymphocytes of any specificity and exercise its ability to kill the lymphocytes (Fig. 6). In this way, we suggest that anti-CD4 autoantibodies turn CD4 lymphocytes into a target for RF and involve RF in a pathological process that results in immunodeficiency.
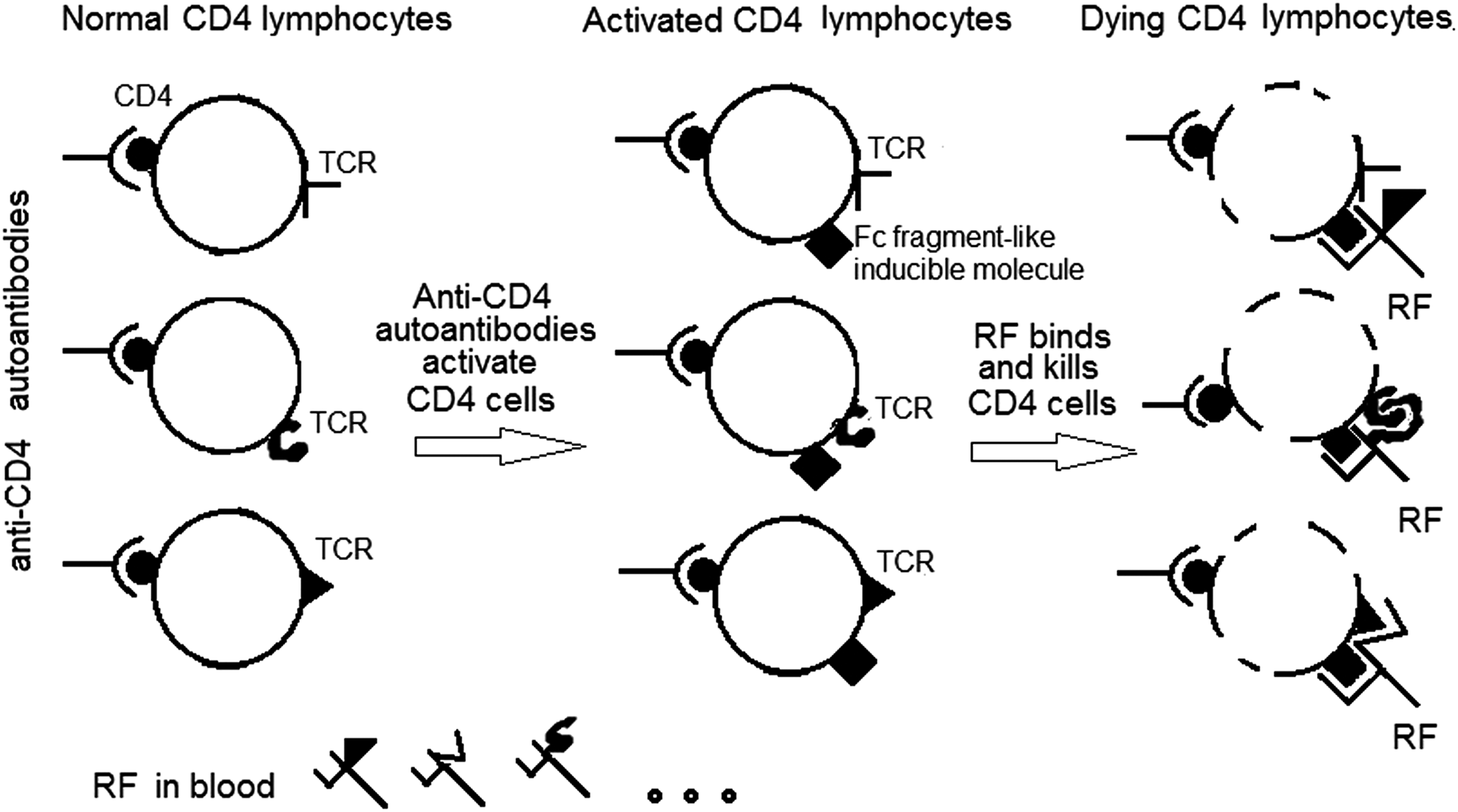
A hypothetical scheme of death of HIV-uninfected CD4 cells in the course of HIV infection. Anti-CD4 autoantibodies produced in response to gp120 HIV-1 induce polyclonal activation of peripheral CD4 lymphocytes and the appearance thereon of the ligand (IgG Fc fragment-like molecule) for the shared paratope of RF. The RF normally in the blood gets the opportunity to bind with CD4+ lymphocytes and exercise its ability to kill the activated lymphocytes.
Footnotes
Acknowledgments
This study was supported by the Ministry of Education and Science of the Russian Federation (grant no. 14.124.13.1159-MD). We are grateful to Jennifer Guernsey for editorial assistance. We thank Tatiana Tolstolutskaya at the Izhevsk State Medical Academy for technical assistance.
Author Disclosure Statement
No competing financial interests exist.