
Abstract
The circulating subtype distribution of HIV-1 has not been well characterized in female sex worker (FSW) populations in Thailand. To understand the mechanisms and interrelationships of epidemics involving FSWs in Thailand, we performed a large molecular epidemiological study of FSWs aged 25 years with recently acquired HIV-1 infections. The samples were collected in 2005, 2007, 2009, and 2011 in 38 provinces, representing every region of Thailand. After gag (p24), pol (pro-RT), and env (C2/V3) were sequenced, comprehensive genome analysis was performed. Genetic subtypes were determined in 159 plasma samples. The percentage of circulating recombinant forms (CRFs) CRF01_AE (90.6%) predominated, while subtype B (1.3%), other CRFs (1.9%), and unique recombinant forms (URFs) (6.2%) were identified as minor populations. Interestingly, the unique recombinant nature of these HIV-1 strains was verified in 10 specimens, indicating the presence of new forms of HIV-1 intersubtypes G/A, C/B, AE/B/C, and AE/B with different recombination breakpoints. Subtype B has contributed to these new generations of unique CRF01/B recombinants, especially in the pol (RT) gene, in which the template switching of the RT genomes occurred during reverse transcription. These results imply that the several unique recombinant viruses circulating in Thailand were probably generated in the population or introduced from neighboring countries. Our study helps clarify the patterns of viral transmission and define transmission pathways in Thailand.
I
Since 1989, HIV-1 subtype B has been the predominant subtype among IDUs, whereas CRF01_AE has been the main clade among individuals with heterosexual exposure risk. 4 The HIV-1 epidemic in Thailand has changed, with CRF01_AE now accounting for the majority of HIV-1 infections in high-risk groups, that is, MSMs and IDUs. 5,6
However, there is no recent information about the distribution of HIV-1 subtypes/CRFs in the FSW population. Therefore, an understanding of the distribution of HIV-1 clarifies the dynamics of HIV-1 epidemics among the FSW populations in Thailand.
We determined the nucleotide sequences of the gag, pol, and env regions from plasma viral RNA. All samples were characterized by targeted genomic sequencing (Table 1). The nested polymerase chain reaction (PCR) products of the gag (p24), pol (pol-RT), and env (C2V3) regions were 671, 1,277, and 565 bp long, respectively. The samples showing possible recombinant or non-AE were further amplified and sequenced with the gp41-nef primer set. The nested PCR products were 800 bp long. The nucleotide sequences were aligned with the HIV-1 group M (subtypes A–J) reference strains, and selected references were found previously in Asia: HIV-1 subtype B-US (B) (accession numbers: AY835778 and K02007), subtype B-Thai (B′) (AY945710 and DQ354116), subtype C (AF067158 and AF361875), and CRF01_AE (JX446848, JX446654, AY713425, U51189, and AY008714) strains. Phylogenetic trees were constructed by the neighbor-joining method with 100 bootstrap replicates. These analyses were conducted using MEGA version 6.0 software.
Primers are assigned a primer name_primer direction (forward, F, or reverse, R), followed by a nested outward (O) or inward (I) primer.
Provided by Dr. Masashi Tatsumi.
All samples were collected from Thailand's National HIV Drug Resistance Surveillance Program. The plasma samples of venue-based FSWs were then sent from 38 of 77 provinces, representing every region of Thailand. The FSWs had neither a previous pregnancy nor an antiretroviral treatment history. The enrolled HIV-infected FSWs are shown in Table 2. In brief, the samples were screened and confirmed for the presence of HIV-1 antibodies. The World Health Organization (WHO) recommends collecting 45–65 samples for an HIV drug resistance surveillance program. 7 When the number of enrolled samples using the BED IgG-Capture Enzyme Immunoassay (BED-CEIA) was insufficient, we used the criterion of 25 years of age at the time of enrollment.
CRF, circulating recombinant form; WHO, World Health Organization.
To identify the recombination breakpoints, we used SimPlot 2.5 software, recombination detection and analysis (RDP2), and
As Table 1 shows, of the 34,313 FSWs enrolled in 2005, 2007, 2009, and 2011, 1,408 FSWs were positive for HIV-1 infection. The prevalence of HIV-1–positive FSWs was 4.1% (95% CI: 3.9–4.3). We successfully amplified and sequenced gag, pol, and env genes from 159 samples. Of these subjects, the mean age was 21.2 years (range, 16–25 years). The genetic subtype for each of the regions gag, pol, and env was determined in the 159 specimens (Table 1). The neighbor-joining analysis placed 90.6% (144/159) of the sequences in HIV-1 CRF01_AE with high bootstrap values (more than 90/100) for the three gene regions (Fig. 1). The prevalence of CRF01_AE in 2005, 2007, 2009, and 2011 was 87.2%, 88.6%, 100%, and 90%, respectively.
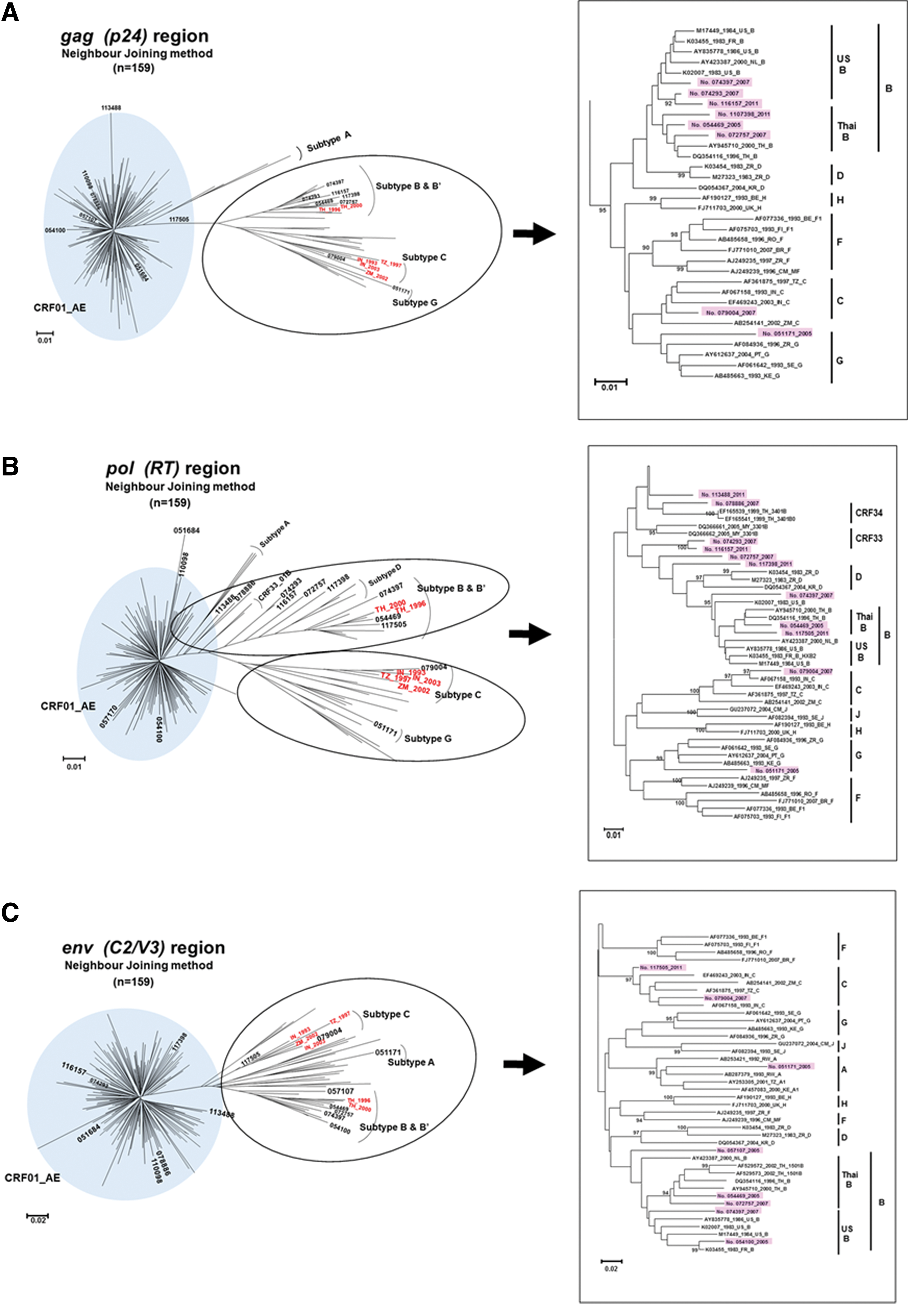
The distribution of HIV-1 subtypes circulating among female sex workers in Thailand. Phylogenetic trees based on
In the gag trees, most of the sequences belonged to the monophyletic cluster of HIV-1 CRF01_AE, distinct from the nonrecombinant form of subtype A (Fig. 1A). There were eight gag sequences located in different clusters in the phylogenetic tree (Fig. 1A). The phylogenetic analysis of the pol region showed several clusters, and most of the sequences were located in pol CRF01_AE (Fig. 1B). Interestingly, many putative recombination patterns between CRF01_AE and B were detected in this pol tree. We found breakpoints of the pol gene in several specimens (Fig. 2), and most of them were recombinant between subtype B and CRF01_AE. Most of the env sequences formed a single cluster within env CRF01_AE, except for eight specimens that were placed in env subtypes B-US, B′, C, and A (Fig. 1C).

Genome structures of 15 samples. The nucleotide sequence determination in relation to the reading frames of corresponding HIV-1 genes. Each deduced subtype structure is illustrated based on the results of the phylogenetic tree analysis. Regions in color could be assigned to known subtypes. Orange, subtype A; blue, subtype B; yellow, subtype C; red, CRF01_AE; and purple, subtype G. The region in white was not PCR amplified. PCR, polymerase chain reaction. Color images available online at
The subtype classification and distribution of 15 samples were obtained with the use of the jpHMM program based on four genes (Fig. 2). The genetic subtype of the three gene regions was determined to be CRF01_AE in 90.6% (144/159). Of the 159 samples, 15 showed a non-AE result, and we further sequenced the gp41-nef regions of these 15 samples. There were one subtype B-US, one B′, two CRF15_01B, and one CRF54_01B. The other 10 samples showed discrepancies from all known CRFs and subtypes, indicating that we found URFs in this study. No. 117505 and no. 110098 showed complicated genome patterns, each composed of three subtypes: CRF01_AE, B, and C (Fig. 2). No. 051171 and no. 079004 were similar to, but distinct from, CRF02_AG and subtype C, respectively.
We used the recombinant identification programs SimPlot, RDP2, and jpHMM to determine whether the apparent discrepancy of subtypes in the gag, pol, and env regions in the specimens was due to coinfections or a recombination of multiple HIV-1 subtypes (Figs. 3 and 4). Since these programs gave similar results, only the plots created by the SimPlot program are presented (Fig. 3). We identified 10 putative recombinant forms between HIV-1 subtype B and CRF01_AE (Fig. 2). However, the locations of the recombination breakpoints in these specimens were not identical (Fig. 3A–H). Multiple recombination breakpoints between HIV-1 subtype B and subtype G were identified in the pol region of no. 051171 (Fig. 3I).

Recombination breakpoint analysis in the 1,277-bp region encompassing RT of pol region. The specimens
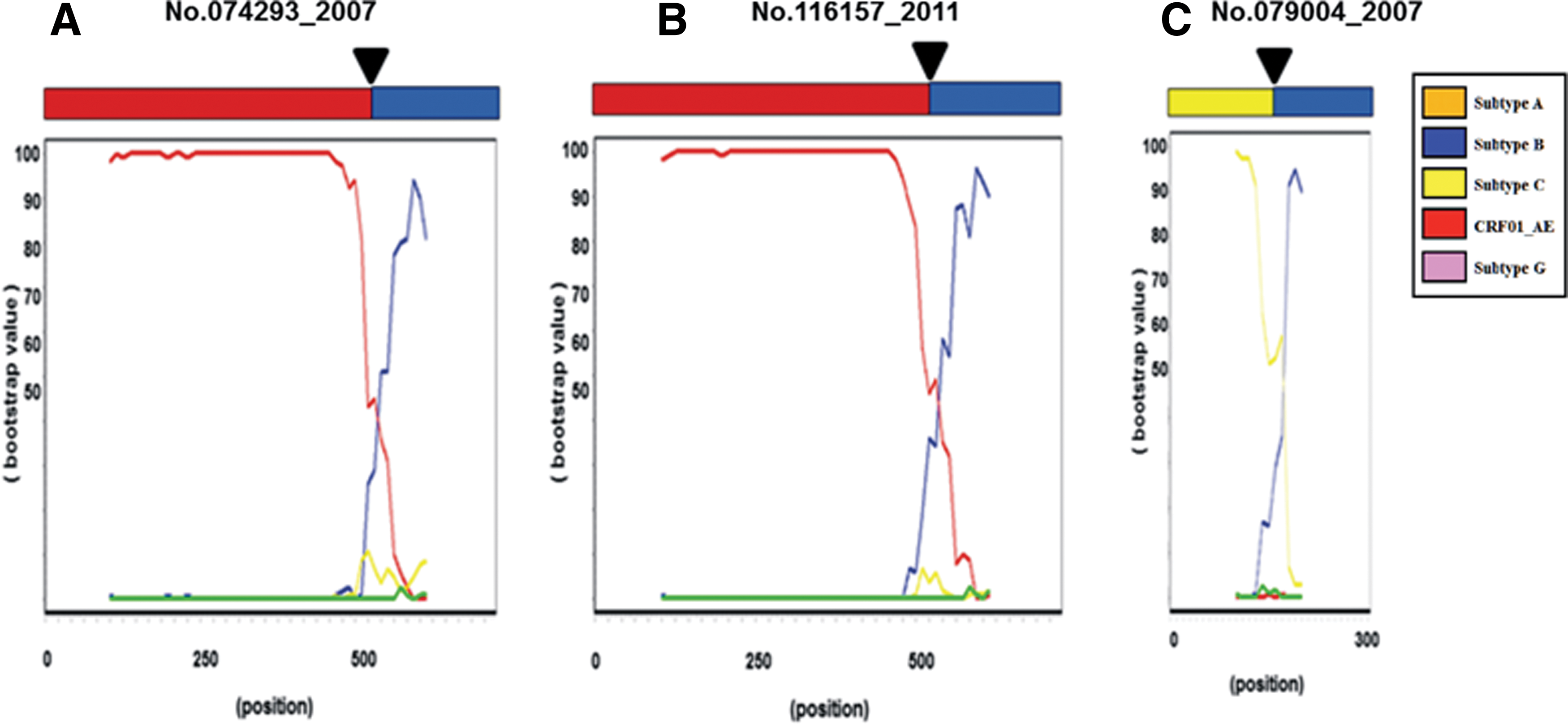
Recombination breakpoint analysis in the 674-bp
These breakpoints in the pol region of no. 051171 belonged to different genome patterns of CRF02_AG (Fig. 3I). No. 051171_2005 from Chachoengsao Province revealed a new unique recombinant exhibiting subtype G gag (not subtype A gag) and an independent cluster and subtype G pol. Similarly, in no. 072757_2007, multiple recombination breakpoints between HIV-1 subtype B and CRF01_AE were identified in the pol region (Fig. 3B). No. 072757_2007 carried gag subtype B, a pol-independent cluster, and subtype B (Fig. 3B). Recombination breakpoints were not identified in the pol regions of the remaining two specimens, no. 054100_2005 and no. 057107_2005. However, their genome patterns were similar to those of CRF15_01B (Fig. 2). These specimens will be further analyzed to clarify whether the genome patterns of the specimens are due to the recombinant form or coinfection.
To discriminate the genomic patterns of the specimens, we also analyzed the gp41-nef region using SimPlot. Two of the 10 subtype B/CRF01_AE recombinants, no. 074293_2007 and no. 116157_2011, were found to contain the breakpoints of the gp41-nef region (Fig. 4A, B). No. 079004_2007 from Tak Province was initially classified as HIV-1 subtype C (gag C/pol C/env C). However, a breakpoint was identified in the gp41-nef region and was found to be a new and unique recombinant form (URF) between subtype C and subtype B (Fig. 4C). No. 117505_2011 from Pang-Nga Province revealed three different subtypes in each fragment gene (gag CRF01_AE/pol B/env C) (Fig. 2).
This is the first large-scale molecular epidemiological study of the FSW population in Thailand to be conducted from samples collected between 2005 and 2011. Our data were derived from a large longitudinal cohort study, which is a part of the National HIV Drug Resistance Surveillance Program. Our results provide evidence of patterns of viral transmission from region to region and a clue toward defining the transmission pathways between groups and individuals based on sequence data.
Our study revealed that the predominant HIV-1 subtypes are CRF01_AE in 90.6%, B in 1.3%, CRF01_AE/B recombinants in 5.7%, and other URFs in 2.5%, as depicted in Figure 1. This distribution is mostly consistent with the results of the Bangkok MSM Cohort Study, 5 which reported 82% CRF01_AE, 5% B, and 13% CRF01_AE/B recombinants. In Thailand, CRF01_AE and CRF01_AE/B recombinants are the major circulating strains among FSWs and MSMs. The molecular epidemiology of HIV-1 among IDUs in northern Thailand indicated that CRF01_AE (86.8%) and CRF01_AE/B′ (13.2%) recombinants are dominant in this risk group. 6 An increasing number of infections of CRF01_AE/B′ recombinants were reported among all risk groups in Thailand. 6,8,9 Furthermore, the distribution has shifted from B to CRF01_AE among IDUs. Other studies reported similar shifts in China and Japan. 10 –13 The reasons for the changes in distribution are unclear, but the shifts could be related to different behaviors between these high-risk groups. Our present phylogenetic analyses of the gag, pol, and env regions showed that URFs similar to HIV-1 subtype C sequences from the western area of Thailand tended to be phylogenetically more closely related to those found in India, whereas they were weakly related to the HIV-1 subtype C distributed in African countries (Fig. 1). Subtype C was identified in Tak Province, near the border with Myanmar. This sample is so closely related to subtype C identified in India that it could be linked with Myanmar or India.
A limitation of the present study was that, because of the cost of analysis, we analyzed only parts of the full-length genome and used fewer clinical samples than would have been desirable. The four fragment genes might have failed to detect new recombinant form strains or URFs. Our results indicate that subtype B has not become extinct but rather has contributed to these new generations of CRF01_AE/B recombinants, especially in the pol (RT) gene. The recombination points of many URFs were identified in the pol (RT) region. One possibility among the many recombinations in the RT gene is the template switching of the RT enzyme between the two RNA genomes during reverse transcription. 14 Since FSWs have many opportunities to become exposed to genetically different HIV-1 strains, many URFs might be easily generated between CRF01_AE/B recombinants and pure CRF01_AE, which are the two major dominants in Thailand.
Our analyses of intersubtype mosaicism detected possible recombination breakpoints in at least 12 specimens. Of these, 10 specimens showed intersubtype recombination between HIV-1 CRF01_AE and subtype B. One specimen appeared to have breakpoints between subtype A and subtype G. The mosaic pattern of the latter specimen was not consistent with CRF02_AG. A new recombination virus may emerge among high-risk populations exhibiting the highest HIV infection rates in the region. Two specimens appeared to be these recombinants: CRF15_01B and CRF54_01B. Actually, many CRF01_AE/B′ recombinants have been detected in Southeast Asian countries. 15,16 The present findings indicate that our study should be continued to detect the emergence of more CRFs and URFs.
The unique recombinant nature of these HIV-1 strains was verified in 10 specimens, indicating the presence of new forms of HIV-1 intersubtypes G/A, C/B, AE/B/C, and AE/B with different recombination breakpoints. These data suggest that multiple subtypes of B, C, and CRF01_AE are cocirculating in Thailand, leading to the evolution of new forms of recombinants among the risk populations that exhibit high HIV infection rates there. Thailand functions as a crucial intersection, connecting the countries in Southeast Asia as well as international traffic with southern China (Yunnan Province) and Myanmar. Reflecting Thailand's geography, the results of the present study revealed the unique distribution and occurrence of HIV-1 subtypes and their intersubtype recombinants.
In conclusion, the cocirculations of multiple subtypes of HIV-1 among persons engaging in high-risk behaviors in Thailand provide unique opportunities for coinfection of different HIV-1 subtypes and the generation of various forms of HIV-1 intersubtype recombinants. More structural information on the HIV-1 strains prevalent in these geographically intermingled regions is crucial to elucidate the dynamics of how HIV spreads and develop vaccine strategies in this unique area in Asia. To decrease the incidence of HIV, we need to continue the surveillance of HIV molecular epidemiology every 3–5 years. Thailand has intensified HIV prevention efforts among MSMs (including TG), MSWs, and FSWs as well as their clients. This includes expanded outreach to target populations, improvements in service linkages, and increased referrals for prevention and treatment as needed.
Ethical Statement
Questionnaire information was deidentified by the reviewer and recoded so that no information could be linked to any individual participant. The protocol of this study was reviewed and approved by the Ethics Committee of the Institute for the Development of Human Research Protections (IHRP) in Thailand. The Ethics Committee waived the need for consent.
Footnotes
Acknowledgments
We acknowledge the support of and funding from the Japan Initiative for Global Research Network on Infectious Diseases (J-GRID). We thank all the following individuals: the FSWs who participated in the study; the staff of the Bureau of Epidemiology; the 38 Provincial Public Health Offices (for collecting the samples); Dr. Achara Teeraratkul and Dr. Kunjanakorn Phokhasawad of the Thai MOPH–U.S. CDC Collaboration (for their supportive and helpful advice); Dr. Masashi Tatsumi (for the primer information); Mr. Nattapong Kamklai, Ms. Ratrawee Boonmuang, and Dr. Piraporn Utchee (for their laboratory support); and Ms. Dorothy L. Southern for providing guidance on quantitative article writing and critical reviews of the article.
Author Disclosure Statement
No competing financial interests exist.
Sequence Data
Partial sequences of the HIV CRF01_AE gag, pol, and env genes analyzed in this study have been deposited in GenBank under accession numbers LC114516–LC114978. The subtypes were further confirmed by phylogenetic analysis using the reference sequences representing subtypes A–D, F–H, J, K, CRF01_AE, CRF15_01B, CRF33_01B, and CRF34_01B (