
Abstract
CRF01_AE is one of the four dominant HIV-1 strains circulating in China. In this study, we performed genetic and phenotypic analyses using a total of 60 full-length envelope gene clones from 14 HIV-1-infected individuals in the Beijing area. Among the 60 sequences analyzed, 32 have a complete open reading frame (ORF), whereas the others contain premature stop codons. The phylogenetic tree analysis suggested that all of the sequences maintained a close relationship with the CRF01_AE strain. Most of the potential N-linked glycosylation sites (PNGS) were located within the V1/V2, V4, C2, or C3 regions. In relation to gp41, the majority of the glycosylation sites were located in the ectodomain. The 32 env genes that contained intact ORFs were used to construct Env-pseudotyped viruses, and eight strains that resulted in high titers were further studied. All the eight strains used CCR5 as the co-receptor for infection, and they were sensitive to neutralization by the broadly neutralizing monoclonal antibodies, including VRC_01, PG9, PG16, and NIH45-46, but they were insensitive to 2G12. Notably, seven of these eight strains lacked a glycan at residues 295 or 332 (or both), suggesting that these two PNGSs play an important role in 2G12 binding and neutralization. In addition, the pseudoviruses were more sensitive to neutralization by plasma isolated from individuals infected with subtypes CRF01_AE and CRF07/08_BC, suggesting the occurrence of a cross-neutralizing antibody profile between these two strains. These findings are likely to have important implications for the design of an effective HIV vaccine and relevant therapeutic drugs.
Introduction
T
In recent years, CRF01_AE has spread rapidly in China. A recent molecular epidemiology survey in Beijing identified HIV-1 CRF01_AE (40.4%) as the dominant strain in HIV infections. 9 The factors associated with the rapid increase of CRF01_AE infections in China are not completely understood. The genetic complexity of this strain posed tremendous challenges to human immune systems and vaccine development. A better understanding of how the biological functions of the HIV-1 envelope (Env) change during disease progression may aid in the design of an efficacious anti-HIV-1 vaccine. Although studies from patients have provided some insights in relation to these issues, differences in the study cohorts and associated methodologies have made it difficult to reach a consensus related to the variations associated with HIV-1 Env function during disease progression.
In the present study, full-length env genes were amplified from proviral DNA isolated from CRF01_AE-infected patients. The genetic properties of the cloned env genes were analyzed and it was found that the V1, V2, V4, and V5 regions exhibited substantial length variations, whereas the observed V3 regions were relatively conserved. The cloned env genes that were shown to contain intact open reading frames (ORF) were used to construct Env-pseudotyped viruses. Following these manipulations, eight strains that facilitated the development of high titers were observed. Each of the latter strains was shown to use CCR5 as the co-receptor that facilitated infection. These strains were also shown to display sensitivity to neutralization by VRC_01, PG9, PG16, and NIH45-46. Conversely, the strains were insensitive to 2G12. Additionally, the pseudoviruses were more sensitive to neutralization by plasma isolated from patients infected with subtype CRF01_AE and CRF07/08_BC. It is very likely that these findings will have important implications for the design of an effective HIV vaccine and HIV-related therapeutic drugs.
Materials and Methods
Study subjects
This study was approved by the Ethics Committee of the National Institute for Viral Disease Control and Prevention, the Chinese Center for Disease Control and Prevention and written inform consents were obtained from all participants. The peripheral blood samples for cloning env genes were collected at a single time point from 14 HIV-1-infected patients. The demographic and immunologic characteristics of these subjects are summarized in Table 1. These samples were received by Beijing Ditan hospital.
The age of patients at the time of sampling.
CRF, circulating recombinant form; F, female; M, male; NA, not available.
Plasma samples and broadly neutralizing monoclonal antibodies
Individual plasma samples were obtained from 88 infected patients. Viral RNA was extracted from patients’ plasma using a QIAamp® Viral RNA Mini Kit (Qiagen, Inc.). A region of the gag gene was amplified from viral RNA using TakaRa's One-Step RNA PCR Kit (AMV) as previously described. 2 The subtypes associated with each of the samples were determined by partial gag sequence analysis. All plasma samples were heat inactivated for 1 h at 56°C and stored at −80°C thereafter until further required.
Besides 2G12, several recently identified potent broadly neutralizing monoclonal antibodies (bnmAbs), PG9, PG16, VRC_01, and NIH45-46 were also used to evaluate the neutralization sensitivity in this study. The 2G12, VRC_01, PG9, and PG16 were expressed and purified in our labs earlier. NIH45-46 was obtained from the National Institutes of Health AIDS reagents program.
Polymerase chain reaction amplification and cloning of env gene
Full-length envelope genes were amplified using the nested polymerase chain reaction (PCR). The amplifications were performed on proviral DNA that had been extracted from patients’ uncultured peripheral blood mononuclear cells using a QIAamp blood DNA isolation kit (Qiagen, Inc.). The following primer sets were designed after comparing multiple complete HIV-1 CRF01_AE sequences. These primer sets were subsequently used for the nested PCR:
The outer primers, env-F1 (5′-GTAGATCCTAACCTAGAG-3′, nucleotides 5840–5857) and env-R1 (5′-CTAGATCTTGAGATACTGCTCCTAC-3′, nucleotides 8884–8908), were used for the initial PCR. The PCR was conducted using an initial denaturation step at 94°C for 5 min, followed by 30 cycles of 94°C for 30 s, 50°C for 30 s, and 68°C for 3 min 30 s and was followed by a final extension at 68°C for 10 min. Inner primers, env-F2 (5′-GCGAATTCATGGCAGGAAGAAGCG-3′, nucleotides 5970–5985) and env-R2 (5′-CCGCTCGAGGACCACTTGCCTCCCATGTTATA-3′, nucleotides 8791–8813), were used for the nested PCR. The amplification required an initial denaturation step of 94°C for 5 min, followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, and 68°C for 3 min, with a final extension of 68°C for 10 min.
The numbers associated with the nucleotide positions correspond to the HXB2 nucleotide sequence (GeneBank accession No. K03455). The amplified HIV-1 genomic fragment, containing the full-length Env precursor glycoprotein gp160 and Rev genetic sequences, was cloned into pcDNA3.1 (+). The cloning reaction required the use of EcoRI and XhoI restriction enzyme recognition sequences, which were located in the “Inner primer” sequences. Positive clones, containing the desired genomic fragment, were selected and sequenced using vector universal primers. Cycle sequencing and dye terminator methods facilitated by an automated DNA sequencer (made by Tsinke Co. Ltd.) were used to generate sequencing data.
Phylogenetic and sequence analyses
Phylogenetic analysis of the HIV-1 env gene sequences was conducted using MEGA4 software, following the use of the multiple alignment Clustal W algorithm tool and manual editing. Potential N-linked glycosylation sites (PNGSs) were evaluated using N-glycosite, and they were counted manually.
Preparation of pseudoviruses
Env-pseudotyped viruses were generated after co-transfection of env/rev expressing plasmids together with an env-deficient HIV-1 backbone vector (pSG3ΔEnv) into 293T cells. Forty-eight hours post-transfection, pseudovirus-containing culture supernatants were harvested, titrated, and stored (in small aliquots) at −80°C until further required.
Determination of viral co-receptor usage
Determination of the use of co-receptors by env-pseudotyped viruses was conducted as previously described. 10 Briefly, GHOST cells expressing CD4 and CXCR4 (Ghost-X4), or CCR5 (Ghost-R5), were seeded in 24-well plates at a cell density of 1 × 105 cells/well and incubated at 37°C in a 5% CO2 incubator overnight. The next day, the monolayers associated with the GHOST cells were infected with virus stocks containing 40 μg/ml of 2-diethylaminoethyl (DEAE)–dextran (to enhance the infection efficiency). After 48 h, the cells were harvested and analyzed using flow cytometry. Approximately 10-fold shifts in fluorescence levels between the mean GFP fluorescence associated with infected cells and uninfected cells were considered positive for the corresponding co-receptor. 10 Ghost-R5 and Ghost-X4 cells infected with HIV-1, HIV-1 SF162 (R5), and HIV-1 NL4-3 (X4) were used as positive controls and the HIV-1-uninfected cells were used as the negative control.
Neutralization assay
Neutralization activity associated with patients’ plasma samples and bnmAb activity against the pseudoviruses were measured using a reduction in Luc-reporter gene expression after a single round of viral infection in TZM-bl cells as previously described. 11 The 50% inhibitory dose (ID50) was defined as the plasma dilution or sample concentration at which the level of relative light units (RLU) was reduced by 50% compared with virus-containing control wells. This value was generated after subtraction of the background RLU value pertaining to the cell-containing control wells. Briefly, 100 tissue culture infective dose 50 (TCID50) of virus was incubated with various dilutions of test patients’ plasma (50-fold dilution) or mAb samples (14 dilutions in a 2-fold stepwise manner). This incubation was performed in duplicate for 1 h at 37°C in a total volume of 150 μl of growth medium in 96-well flat-bottom culture plates. Control wells lacking mAbs were also used, and this was performed in quadruplicate. Freshly trypsinized cells (10,000 cells in 100 μl of growth medium containing 40 μg/ml of DEAE–dextran) were added to each well. After 48 h of incubation, 150 μl of culture medium was removed from each well and 100 μl of the luciferase reporter gene assay system reagent (Bright-Glo; Promega) was added. After a short incubation period (minimum of 2 min), 150 μl of lysate attained from each well was transferred to 96-well black solid plates for luminescence measurement using a luminometer. Mock-infected cells were used to determine background luminescence.
Results
Functional domain analysis of full-length envelope sequence
A total of 60 full-length envelope genes were successfully amplified from peripheral blood samples from 14 HIV-1-infected individuals. Among the 60 sequences that were analyzed, 32 contained a complete ORF, whereas the remaining 28 sequences contained premature stop codons. The phylogenetic tree that was generated to compare these sequences confirmed that all of the sequences analyzed had a close relationship with CRF01_AE (Fig. 1). The identity and degree of polymorphism length associated with the deduced amino-acid sequences from the conserved and variable regions of Env gp120 (as well as gp41) were analyzed after a comparison with HIV-1 gp160 sequences. The results showed that the identities of the amino-acid sequences in the gp120 variable regions, V1/V2, V4, V5, were lower than those determined for other regions, including gp41, and the conserved regions, C1-C5 of gp120 (Table 2). The V3 region showed relatively high identity when compared with the other variable regions (Table 2). In addition, C3 showed the lowest identity among the conserved regions of gp120 (Table 2). Finally, gp41 showed higher identity than gp120, suggesting that gp41 is more conserved (Table 2). The V1/V2 (72.3 ± 3.1), V4 (26.7 ± 2.5), and V5 (10.8 ± 1.1) regions displayed a relatively high degree of length variation, whereas the C1 (129.0 ± 0.2), C2 (99.0 ± 0), C3 (51.6 ± 0.6), C4 (41.0 ± 0), and C5 (41.0 ± 0) regions showed little length variation. The V3 loop showed constant length among the sequences analyzed (35 residues). Furthermore, we examined the changes in the number and location of PNGS in the functional regions of gp160. The number of PNGS in gp120 and gp41, on average, is 25.3 and 4.1, respectively. Little change was found in the number of PNGS in the V3, V4, and C5 regions. In contrast, some variation was observed for V1/V2 and V5 (Table 3). Most of these glycosylation sites were located within the V1/V2, V4, C2, and C3 regions (Table 3). In gp41, a large majority of the glycosylation sites were located in the ectodomain, outside the transmembrane (Fig. 2).
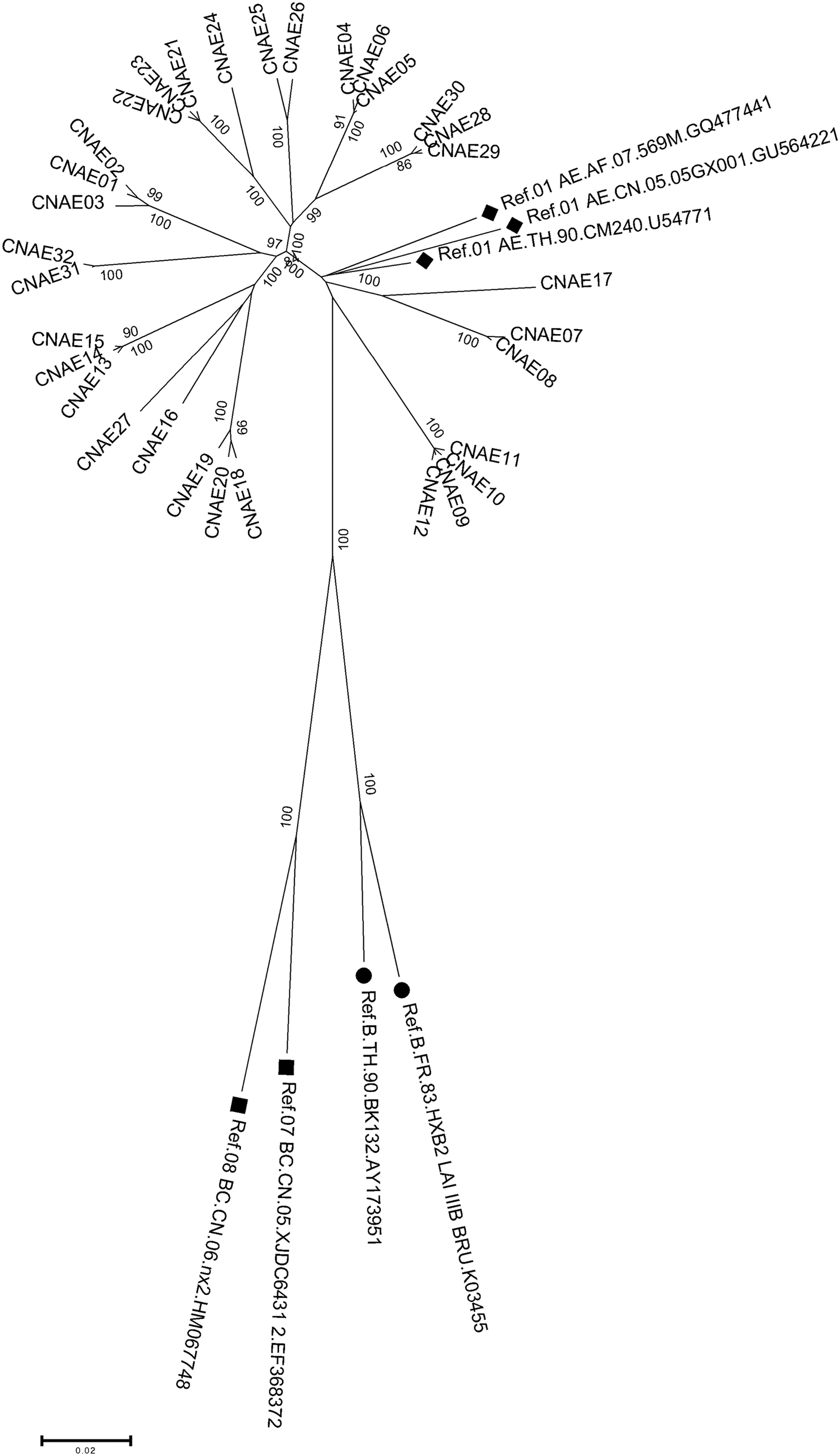
Phylogenetic analysis of the gp160 envelope region. The analysis was performed using the neighbor-joining method using MEGA version 5.0. Positions containing an alignment gap were included for pair-wise sequence analysis. The black solid icons indicate reference sequences from the Los Alamos HIV sequence database.
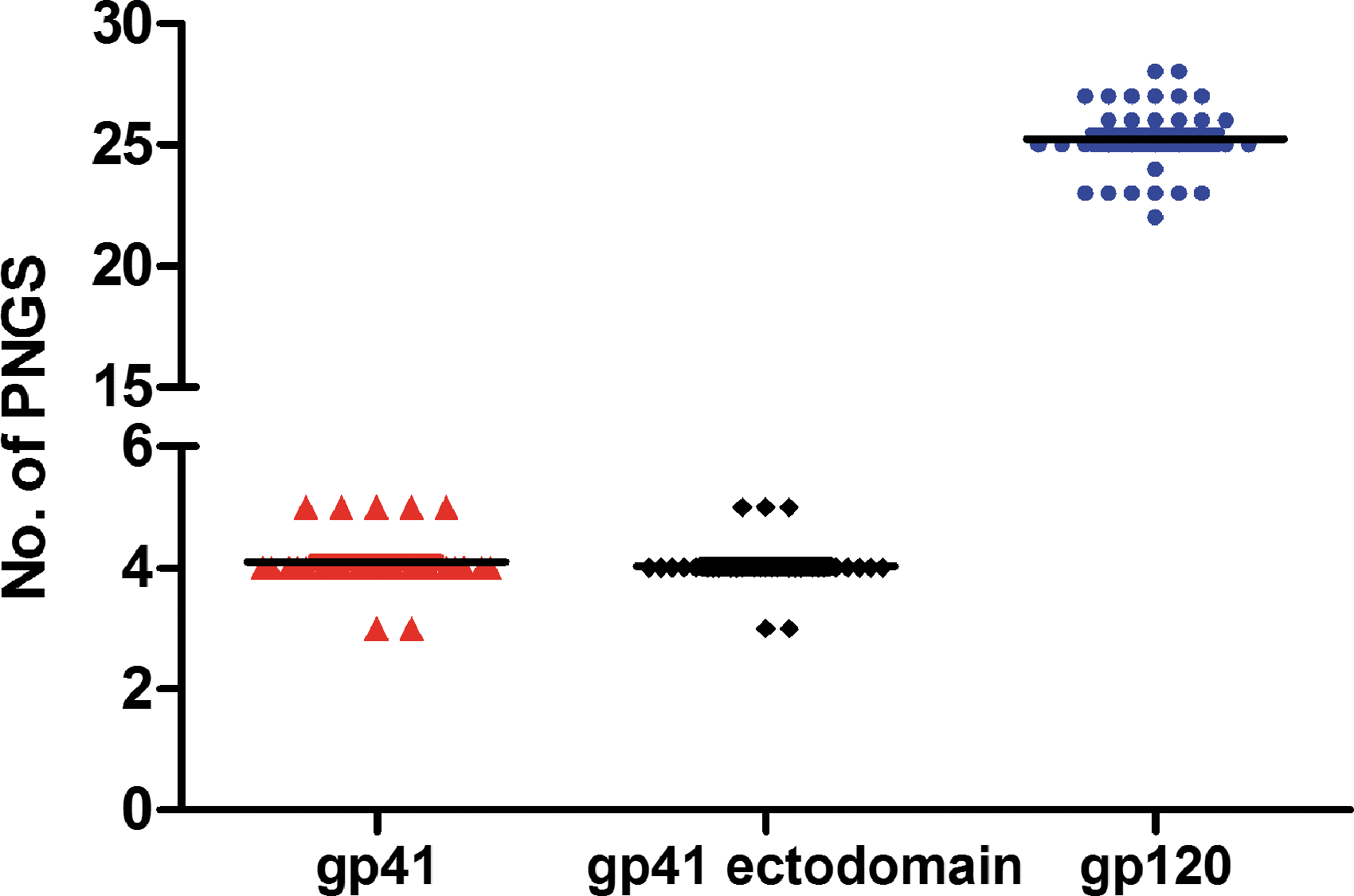
Number of potential N-linked glycosylation sites in the env sequences.
Reference strain.
SD, standard deviation.
Co-receptor usage of pseudotyped viruses
Overall, 32 pseudotyped viruses were generated. Infectivity associated with these viruses was tested by measuring luciferase activity in infected TZM-bl cells. We found that only 14 of the generated pseudotyped viruses were able to mediate viral entry, whereas the remaining 18 env-recombinant viruses demonstrated no infectivity, despite having complete ORFs at the nucleotide level. This indicated the existence of functional defects associated with the latter pseudotyped viruses. Eight of the 14 pseudotyped viruses demonstrating high infectivity were chosen to study biological and neutralization sensitivity. Genotypic predictions associated with co-receptor usage, based on V3 sequence and net charge, suggested that all of the eight viruses chosen used the CCR5 co-receptor. A comparative examination that was performed on GHOST cells verified this assessment (Table 4).
Neutralization sensitivity of env-pseudotyped viruses to bnmAbs
We initially examined the neutralization susceptibility associated with CRF01_AE Env-recombinant viruses in relation to five well-known bnmAbs, that is, 2G12, PG9, PG16, VRC_01, and NIH45-46. A single pseudotyped virus expressing an Env antigen from a sensitive primary virus (SF162) was included as a control. As shown in Table 5, 2G12 failed to cause neutralization of the eight analyzed viruses with the half maximal inhibitory concentration (IC50) exceeding 50 μg/ml in each case. Epitope sequence analysis of these resistant viruses demonstrated that seven of the eight viruses analyzed lacked a glycan residue at residues 295 or 332 or both (Table 6). These results indicate that the two PNGSs play an important role in 2G12 binding and neutralization. Each of the eight highly resistant viruses was shown to lack the glycan residue at position 397. Interestingly, CNAE17, which contained all of the major glycans apart from the glycan usually positioned at N397, showed high resistance to 2G12, suggesting that a mutation in the latter residue significantly impacts 2G12 binding.
bnmAbs, broadly neutralizing monoclonal antibodies.
Under 2G12, “+” indicates the presence of glycosylation sites, and “−” indicates the absence of glycosylation sites. Under PG9 and PG16, “−” indicates the amino-acid residue at this site was the same as the residue of the epitope recognized by PG9. Letters indicate that the amino-acid residue at this site was mutated to the residue represented by the letter. “Ref.” indicates reference strain.
The PG9 and PG16 mAbs have affinity for epitopes that are located in the conserved regions of the V2-V3 loop associated with the HIV-1 viral spike. Binding of PG9 and PG16 with the virus was largely dependent on the same residues, although PG16 was more sensitive to V3 loop substitutions than PG9. 11 The results of the neutralization assay showed that PG9 and PG16 neutralized a large proportion of the viruses (all except CNAE14 and CNAE31) and the IC50 of the two antibodies were less than 0.39 μg/ml (Table 5). Sequence analysis of PG9- and PG16-resistant viruses revealed complex mutation patterns associated with residues that are critical for PG9/PG16 binding. CNAE14 was shown to be resistant to both PG9 and PG16. It is likely that substitutions S158T, S162T, K305T, and I307T jointly contribute to this resistance phenotype.
Antibodies that target the CD4 binding site (CD4bs) on the HIV-1 spike protein, gp120, have been shown to have increased potency and viral subtype binding flexibility. These neutralizing monoclonal antibodies that facilitate targeting of CD4bs constitute one of the most effective antibody classes identified, with individual members capable of neutralizing up to 90% of HIV-1 strains with the mean IC50 ∼0.1 μg/ml. Previous studies pertaining to viruses displaying natural resistance to VRC01, in conjunction with mutagenesis analyses associated with clade B and CRF08_BC viruses, have suggested that mutations in either of the loop D or V5 regions (or both) may be critical for natural evasion of VRC01. 12,13 However, the resistance mechanisms utilized by CRF01 AE viruses are currently unknown. In our study, four CRF01 AE viruses, CNAE08, CNAE14, CNAE17, and CNAE31, were demonstrated to be resistant to VRC01 (IC50 6.43–26.92 μg/ml, Table 5). Protein sequence alignments associated with sections pertaining to loop D, CD4bs, and the V5 region suggest that the resistance phenotype of the four viruses could be due to the presence of two substitutions, that is, F277L and D279N in the loop D region (Fig. 3). Asn-461 in the V5 region is also a critical determinant of sensitivity to VRC01 in these resistant viruses. The side chain of Asn-461 is also a potential glycosylation site, which may create a steric hindrance effect preventing VRC01 binding. However, experimental analysis that resulted in exchanging the V5 region alone did not affect the sensitivity of the viruses to VRC01. CNAE09, CNAE10, and CNAE11 strains containing the asparagine residue at position 461 were still highly sensitive to VRC01. Of the pseudoviruses analyzed, CNAE17 demonstrated the highest levels of resistance to VRC01. This resistance phenotype may also be due to the presence of mutation S365P in the CD4bs.

Sequence alignment of VRC_01 binding sites in gp120. Yellow indicates sequences that were the same as the reference sequence SF162; “-” indicates loss of amino-acid residues; and red indicates that the mutation at this site might be associated with loss of sensitivity of the virus to antibodies. Blue indicates more than 50% sequences were the same. Green indicates sequence's similarity value was lower than 50%.
NIH45-46 is a more potent clonal variant of VRC01 and was isolated from the same donor patient. In our study, CNAE08, CNAE14, CNAE17, and CNAE31 were all shown to be highly resistant to this CD4bs-targeted antibody (Table 5).
Neutralization sensitivity of Env-pseudotyped viruses to subtype-specific plasma
To evaluate the neutralization susceptibility of CRF01_AE Env-pseudotyped viruses, 88 plasma samples from individuals infected by three viral subtypes or CRF groups were included in this study: 44 samples were extracted from CRF01_AE-infected patients, 26 samples were extracted from CRF07/08 BC-infected patients, and 18 samples were extracted from patients infected with subtype B. Each plasma sample was subsequently diluted (using a 1:50 dilution) due to variation effects associated with the neutralizing antibody (NAb) titers. At this dilution, the percentage of inhibition was reported to indicate the relative sensitivity of the pseudotyped viruses. The inhibition effect was considered negative when inhibition was shown to be less than 50%. Boxes are dark gray coded in Tables 7 –9 and indicate that the plasma showed a greater than 50% inhibition to the pseudotyped viruses.
The row entitled “summary” shows the number of samples where the plasma showed greater than 50% inhibition to the pseudotyped viruses and the percentage of these samples in relation to the total number of samples tested. Boxes are color-coded as follows: dark gray, greater than 50% inhibition; light gray, no inhibition but enhanced infection.
The row titled “summary” shows the number of samples where the plasma showed a greater than 50% inhibition to the pseudotyped viruses and the percentage of these samples in relation to the total number of samples tested. Boxes are color coded as follows: dark gray, greater than 50% inhibition; light gray, no inhibition but enhanced infection.
The row titled “summary” shows the number of samples where the plasma showed a greater than 50% inhibition to the pseudotyped viruses and the percentage of these samples in relation to the total number of samples tested. Boxes are dark gray coded and indicate that the plasma showed a greater than 50% inhibition to the pseudotyped viruses.
Each of the 44 CFR01_AE plasma samples was tested for neutralization susceptibility due to the presence of eight CRF01_ AE pseudoviruses. The data generated revealed that 203 out of the 352 tests performed (57.7%) showed inhibition ratios greater than 50% (Table 7). As few as 32 of the 352 tests performed (9.1%) showed strong neutralization sensitivity of the pseudoviruses to the CFR01_AE plasma samples with inhibition ratios greater than 80%. Five out of eight subtype AE pseudoviruses were shown to be sensitive to CFR01_AE plasma samples and were neutralized by more than half of the samples tested. However, CNAE08, CNAE14, and CNAE31 showed high resistance to subtype AE plasma samples.
Results of neutralization tests performed to assess potential neutralization of subtype AE pseudoviruses in the presence of subtype BC plasma samples (Table 8) showed that 117 of the 208 tests performed (56.3%) resulted in inhibition ratios that were greater than 50%. Indeed, 9.1% of the inhibition ratios were greater than 80%. Half of the eight subtype AE pseudoviruses that were tested (CNAE08, CNAE14, CNAE17, and CNAE31) were only minimally neutralized by the CRF07/08_BC plasma samples.
All eight pseudoviruses were also tested against 18 subtype B/B′ plasma samples (Table 9). Sixty-eight of the 144 neutralization tests (47.2%) demonstrated inhibition ratios of 50%. This suggested that there was minimal cross-reactivity between CRF01_AE and subtype B/B′ plasma samples. Along with CNAE08, CNAE14, CNAE17, and CNAE31, CNAE11 also demonstrated resistance to subtype B/B′ plasma samples.
Interestingly, antibodies from some of the plasma samples used in these tests did not prevent entry of the HIV-1 pseudoviruses, but conversely facilitated a two- to fourfold increase in infection (light gray highlight in Tables 7 and 8).
Discussion
In this study, we performed genetic and phenotypic analyses on a total of 60 full-length envelope gene clones that were isolated from the provirus of 14 HIV-1-infected individuals in Beijing, China. A phylogenetic tree generated as a result of this study suggested that all of the sequences analyzed demonstrated a close genetic relationship with the CRF01_AE strain. The analysis results demonstrated that the V1, V2, V4, and V5 regions of the cloned env genes exhibited substantial length variations. Conversely, the V3 regions of the env genes were relatively conserved. These results may have important implications for the design of anti-HIV-1 vaccines and therapeutic drugs.
These results also demonstrated that the majority of the PNGSs were located within the V1/V2, V4, C2, or C3 regions. On analysis of gp41, a large majority of the glycosylation sites were shown to be located in the ectodomain (outside the transmembrane). It was previously reported that the HIV-1 envelope protein contains more than 40% N-glycans. 14 Additional studies have shown that PNGSs play important roles in maintaining the tertiary structure of Gp120, 15 facilitating evasion of the host immune attack, 16 thereby potentially affecting the infectivity of HIV-1. 17 A recent study compared PNGS profiles from the CRF01_AE strain with the CRF07_BC, CRF08_BC, B, and B′ subtype Chinese isolates. The results demonstrated that the CRF01_AE subtypes contained significantly fewer PNGSs. Additionally, when a more specific analysis was performed, the CRF01_AE subtypes contained fewer PNGSs in the V2/V4 regions, but more PNGSs in the V5 region. Interestingly, this report showed that Gp120 from CRF01_AE had a unique PNGS distribution pattern. This pattern may reflect the prominence associated with the sexual transmission of this subtype in China. 18 These results differ from our findings that showed that most of the PNGS were located within the V1/V2, V4, C2, and C3 regions. However, our results showed that PNGS were distributed throughout the V5 region (Table 3). This is consistent with Shi's study, 18 where the numbers of PNGSs found to be associated with the V1, V2, V4, and V5 loops were found to be fewer in isolated sexually transmitted subjects when compared with intravenous drug users. The PNGS distribution in our isolated CRF01_AE strain may represent a unique pattern, and the cause of its prominence in sexual transmission in China remains elusive.
In this study, we generated Env-pseudotyped viruses using 32 cloned Env genes with intact ORFs. We isolated eight strains that demonstrated high titers from the pseudotypes analyzed. The results demonstrated that each of the eight strains analyzed used co-receptor CCR5 as the co-receptor for infection. This differed from previous results that showed that the CRF01_AE HIV-1 subtype (isolated in China) predominantly uses CXCR4 as the co-receptor for infection. 19 The genetic mechanisms that underpin this phenomenon remain to be elucidated.
The application of advanced technologies has resulted in the isolation of several new HIV-1 mAbs (PG9, PG16, VRC_01, and NIH45-46) that are more potent and broadly cross-reactive than were previous antibodies. It will be crucial to investigate whether the epitopes identified by these bnmAbs are also present on the envelope of HIV-1 CRF01_AE strains isolated in Beijing, China. Our results showed that each of the eight Env-pseudotyped strains analyzed were sensitive to neutralization by the bnmAbs, including VRC_01, PG9, PG16, and NIH45-46. The same strains were shown to be insensitive to 2G12. Notably, seven of the eight strains lacked glycan residues at residues 295 and/or 332, suggesting that these two PNGSs play an important role in 2G12 binding and neutralization. 2G12 has been shown to neutralize a wide spectrum of different HIV-1 isolates, including members from different clades, with the notable exception of clade E. 20 Previous studies revealed that the glycans at N295, N332, and N392 are critical in facilitating binding of 2G12 to gp120, 21 –23 whereas PNGSs at positions 339, 386, and 448 are reported to play an indirect role in the same process. 24,25 As shown in Table 5, 2G12 failed to neutralize each of the eight viruses tested, with the IC50 values exceeding 50 μg/ml. Epitope sequence analysis of these resistant viruses showed that seven of the eight viruses lack glycans at residues 295 and/or 332. These results confirm that these two PNGSs play an important role in 2G12 binding and neutralization. Each of the eight resistant viruses was shown to have lost the glycan at position 397. Interestingly, CNAE17, which contained all of the predominant glycans apart from N397, showed increased resistance to 2G12, suggesting that mutation of this residue significantly impacts 2G12 binding. The mAbs PG9 and PG16 have been shown to bind to a previously undefined site within the V2-V3 region of the HIV-1 viral spike. Crystal structure analysis suggests that VRC01 binds the CD4 binding loop within the gp120 loop D, and the V5 region is necessary for optimal VRC01 neutralization. 26 In addition, VRC01 neutralizes more than 90% of diverse HIV-1 primary isolates, 27 and NIH45-46 is a more potent clonal variant of VRC01. There is an increased interaction between NIH45-46 and the gp120 inner domain, leading to enhanced neutralization. 28 In the present study, CNAE08, CNAE14, CNAE17, and CNAE31 were all shown to be highly resistant to the CD4bs-targeted antibody, suggesting a unique resistance profile of the CRF01_AE isolate that circulates in China. Further studies will be needed to compare the resistance with bnmAbs of CRF01_AE isolates from different countries.
Additionally, the pseudoviruses were shown to be more sensitive to neutralization by plasma isolated from patients infected with subtype CRF01_AE and CRF07/08_BC, suggesting a cross-neutralization antibody profile between these two strains.
It is worthy to note that antibodies specific for viral proteins enhance, rather than inhibit, infection. This phenomenon, known as antibody-dependent enhancement, has been described for HIV. 29 There is increasing concern that passively infused or vaccine-induced antibodies might enhance the chance of infection. It suggests that there is a need for further studies to develop assays that can predict the likelihood that an antibody infusion or vaccine will lead to enhanced infection or pathogenesis.
In summary, this study facilitated a genetic and phenotypic analysis of CRF01_AE env clones isolated from HIV-1 patients in China. The results that were generated showed unique patterns in relation to genetic variation, PNGS distribution (in the V1, V2, V4, and V5 regions), co-receptor tropism, and neutralization antibody profiles. These findings have important implications for the design of potential HIV vaccines and therapeutic drugs.
Footnotes
Acknowledgments
This study was supported by China megaprojects in infectious diseases (2012ZX10001-005). The authors thank all laboratory members of the National Institute for Viral Disease Control and Prevention of China CDC for their support in carrying out this work and the Institute of Infectious Diseases of Beijing Ditan hospital for supplying the HIV-infected patients’ blood and plasma samples.
Author Disclosure Statement
No competing financial interests exist.