
Abstract
Macrophages are targets of HIV-1 infection, and control of viral replication within these cells may be an important component of a T-cell-based vaccine. Although several studies have analyzed the ability of CD8+ T cells to inhibit viral replication in monocyte-derived macrophages, the effect of T cells on HIV-1-infected tissue macrophages is less clear. We demonstrate here that both CD4+ and CD8+ T-cell effectors from HIV controllers are capable of suppressing viral replication in bronchoalveolar lavage-derived alveolar macrophages. These findings have implications for HIV-1 vaccine and eradication strategies.
M
In this study, we examined the effector immune responses to infected MDMs and AMs in three HIV controllers: two were ESs who maintained viral loads below 50 copies/ml without antiretroviral therapy and one was a viremic controller who had low level viremia (<200 copies/ml). All three patients were positive for the protective HLA-B*57/5801 alleles and ES31 was also HLA-B*27 positive. These patients are known to have very effective HIV-specific T-cell responses. 10 All protocols were approved by the Johns Hopkins University institutional review board and written consent was obtained from the study subjects. AMs were obtained through bronchoalveolar lavage and isolated based on adherence to the cell culture plate following a 4 h incubation at 37°C and vigorous pipetting to remove nonadherent cells. To generate MDMs, peripheral blood mononuclear cells isolated from whole blood through Ficoll-Paque PLUS gradient centrifugation were subjected to positive magnetic selection for CD14+ monocytes (Miltenyi Biotech). These monocytes, plated at 105 cells per well, were differentiated for 7 days at 37°C in RPMI 1640 supplemented with 20% human AB serum (GemCell, US origin), 1% HEPES, and 50 ng/ml human recombinant M-CSF (R&D Systems). Fluorescence-activated cell sorting analysis revealed that MDMs generated by this method were greater than 95% CD14+ and contained less than 0.5% CD3+ T cells. On the day of macrophage spinoculation with laboratory strain HIV-1BaL (500 ng p24 of virus per 106 cells), fresh PBMCs underwent positive magnetic selection for CD8+ T cells and negative magnetic selection for CD4+ T cells. Autologous CD4+ or CD8+ effectors were added to 105 target AMs or MDMs at a 1:1 ratio; the cells were then cocultured in RPMI 1640 with 10% FBS for 7 days. Culture supernatant harvested at various time points was measured with an ELISA for p24 content. Supernatant from MDMs and AMs that were not superinfected with HIV-1BAL had p24 values that were below the limit of detection. Infected target cells of the same number as in the experimental conditions were included to determine the maximum amount of virus production for calculating percentage inhibition (% Inhibition = 100% × [1 − experimental value/maximum virus production]). Statistics were calculated through a bootstrap resampling method due to low sample size (n = 3) by employing 1,000 samples of size 3 with replacement of the pairwise differences within subjects.
We first examined the susceptibility to infection of AMs and MDMs in each of the three individuals and found that there was no statistical difference between the two macrophage types (Fig. 1A); however, the impact of these results is limited by the small sample size. To validate the cell adherence method used, we compared responses against HIV-infected macrophages obtained by this method with responses against HIV-infected AMs that were isolated by fluorescence-activated cell sorting based on cell size and HLA-DR expression. The sorted AMs were greater than 92% HLA-DR+ and contained less than 0.05% CD3+ T cells. The HIV-1 replication and the CD4+ and CD8+ T cell responses to infected cells were similar enough to suggest that the cell adhesion method used for isolating AMs was valid (Fig. 1B). CD8+ T-cell responses were generally more effective than CD4+ T-cell responses in inhibiting viral replication within AM with an average of more than 90% suppression seen with the CD8+ T-cell response by day 5 onward compared with an average between 75% and 80% suppression seen for the CD4+ T-cell response (Fig. 1C). Interestingly, the degree of suppression of viral replication in MDMs induced by CD4+ T cells from VC11 was markedly lower than that induced by CD4+ T cells from the two ESs. The suppressive effect of the CD4+ T cells and that of the CD8+ T cells were significantly greater for viral replication in AMs than in MDMs on day 5 (p = .038 and p = .0041, respectively; Fig. 1D). The CD8+ T cells also had a significantly greater suppressive effect on replication in AM on day 7 (p = .021; Fig. 1D). Overall, both primary CD4+ and CD8+ T cells from all three HIV controllers efficiently suppressed viral replication in AMs. This is consistent with a prior study where we showed that both effector cells were capable of inhibiting viral replication: CD8+ T cells through a contact-dependent mechanism and CD4+ T cells through a mechanism that was partially mediated by soluble factors. 5
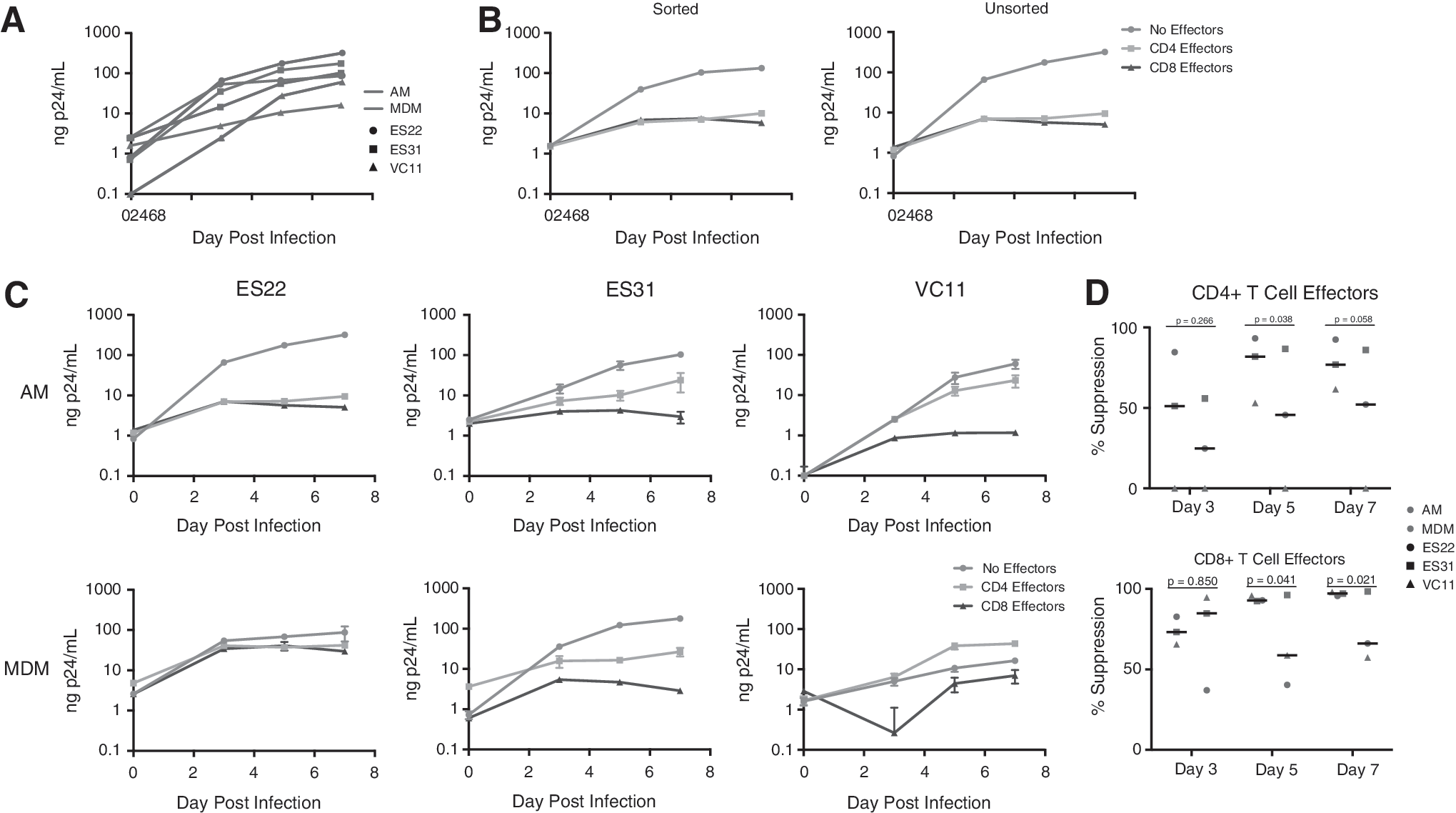
CD4+ and CD8+ T cells are capable of suppressing HIV-1 replication in AMs.
Although a prior study suggested that CD8+ T cells from patients with lymphocytic interstitial pneumonitis could kill AMs at high effector-to-target ratios, 8 our results suggest that T cells from individuals capable of controlling HIV infection can suppress HIV-1 replication in AMs at a 1:1 effector-to-target ratio. This is consistent with the fact that HIV-specific CD8+ T cells from HIV controllers are generally more effective than CD8+ T cells from chronic progressors. 10 This study is limited by the small number of subjects, the lack of chronic progressors, and the fact that we did not determine the mechanism of control of viral replication. In a prior study with 9 HIV controllers and 10 chronic progressors, we showed that CD8+ T cells from controllers were significantly more effective at controlling viral replication in MDMs, and determined that CD8+ T cells were capable of killing infected MDMs. 5 We extend these findings by showing here that CD8+ T cells from three HIV controllers can also control viral replication in AMs. Given that infected cells must be killed during curative strategies, further proof that T-cell-mediated suppression may be capable of eliminating infected tissue macrophages will be important for eradication trials.
Footnotes
Acknowledgments
The study was funded by NIH grants T32HL007534 (A.L.), K23HL103192 (M.B.D.), U01HL121814 and R01HL126549 (G.D.K.) and R21AI096948 (J.N.B.) and the Johns Hopkins University Center for AIDS Research (P30AI094189). We thank Antonietta D'Addio for patient recruitment.
Author Disclosure Statement
No competing financial interests exist.