
Abstract
We assessed CD8+ T cell reactivity to human herpesvirus 8 (HHV-8; Kaposi's sarcoma [KS]-associated herpesvirus) and the role of CD4+CD25hiFoxP3+ regulatory T cells (Treg) in HHV-8- and HIV-coinfected participants of the Multicenter AIDS Cohort Study who did or did not develop KS. There were similarly low CD8+ T cell interferon-γ responses to MHC class I-restricted epitopes of HHV-8 lytic and latent proteins over 5.7 years before KS in participants who developed KS compared to those who did not. T cell reactivity to HHV-8 antigens was low relative to responses to a combination of cytomegalovirus, Epstein–Barr virus and influenza A virus (CEF) peptide epitopes, and dominant HIV peptide epitopes. There was no change in %Treg in the HHV-8- and HIV-coinfected participants who did not develop KS, whereas there was a significant increase in %Treg in HHV-8- and HIV-coinfected participants who developed KS beginning 1.8 years before development of KS. Removal of Treg enhanced HHV-8-specific T cell responses in HHV-8- and HIV-coinfected participants who did or did not develop KS, with a similar pattern observed in response to CEF and HIV peptides. Thus, long-term, low levels of anti-HHV-8 CD8+ T cell reactivity were present in both HHV-8- and HIV-coinfected men who did and did not develop KS. This was related to moderately enhanced Treg function.
Introduction
H
There are several lines of evidence that support a central role for CD8+ T cell immunity in the control of KS, including the high risk of KS in immunocompromised persons due to HIV infection or immunosuppressive therapy for prevention of organ transplant rejection. 4 There is, however, limited information directly linking anti-HHV-8 CD8+ T cell reactivity to the development of KS. 5 Cross-sectional studies show that in vitro CD8+ T cell reactivity to peptides of HHV-8 lytic and latent cycle proteins is demonstrable in HHV-8 seropositive persons with or without HIV infection in the absence of KS, and that these T cell responses are usually greater than those in HIV-infected persons with KS. 6 –10
Regardless of immunosuppressive HIV coinfection, CD8+ T cell responses to HHV-8 are relatively nonrobust compared with those of other human herpesviruses such as Epstein–Barr virus (EBV) and cytomegalovirus (CMV). 9,11 –13 The basis for this weak T cell reactivity to HHV-8 is usually attributed to viral proteins found in KS tumor cells. 1 There is little evidence, however, showing a direct role of these viral proteins in inhibiting host immunity during natural HHV-8 infection and development of HIV-related KS. Alternatively, anti-HHV-8 CD8+ T cell immunity could at least, in part, be dampened by abnormally over-reactive regulatory CD4+ T cells (Treg), which are known to play a role in downregulation of immune function during mouse gamma herpesvirus infection 14 as well as the other human gamma herpesvirus, EBV. 15
To further assess CD8+ T cell immunity in development of KS, and a possible role for Treg in this host response, we conducted a nonconcurrent, prospective study of anti-HHV-8 T cell responses in HHV-8- and HIV-coinfected men who have sex with men (MSM) who did (HIV+HHV-8+ KS+) or did not (HIV+HHV-8+KS−) develop KS, in the Multicenter AIDS Cohort Study (MACS). These responses were compared to T cell reactivity in HIV−HHV-8+, HIV+HHV-8−, and HIV−HHV-8− men.
Materials and Methods
Study cohort
We studied five HIV- and HHV-8-coinfected MSM who developed clinically diagnosed KS before the availability of ART (HIV+HHV-8+KS+) and 10 HIV- and HHV-8-coinfected MSM who did not develop KS (HIV+HHV-8+KS−) in the University of Pittsburgh clinical MACS site. Control groups were five HIV-negative and HHV-8-positive men (HIV−HHV-8+), five HIV-positive and HIV-8-negative men (HIV+HHV-8−), and five HIV-negative and HHV-8-negative (HIV−HHV-8−) men enrolled in the MACS. HHV-8 infection was defined as positive for HHV-8 immunofluorescent antibody specific for lytic antigens. 16 Fourteen of the 15 HIV+HHV-8+ participants were Caucasian and one was African American; the average age was 35 (range 22–64) at study entry, and all had an HLA A*0201 haplotype. Only HLA A*0201-restricted viral epitopes were tested due to the predominance of these anti-HHV-8 T cell epitopes compared to other MHC class I alleles. 5
Archived, cryopreserved peripheral blood mononuclear cells (PBMC) and serum and plasma samples from the five HIV+HHV-8+KS+ MSM were chosen for testing at four time points before and near the date of KS clinical diagnosis [median of 5.7 years (range, 4.5–7.0 years), 3.55 years (3.0–4.0 years), 1.8 years (1.6–2.0 years), and 0 year (0–0.1 year)]. Clinical samples from matching, longitudinal time points were chosen for the 10 HIV+HHV-8+KS−, 5 HIV−HHV-8+, 5 HIV+HHV-8−, and 5 HIV−HHV-8− men. Archived samples from an additional time point ∼1 year after beginning of ART were included for the HIV+HHV-8+KS− and HIV+HHV-8− participants, with samples from a temporal corresponding time point for HIV−HHV-8+ and HIV−HHV-8− men. This ART time point was not testable for the five HIV+HHV-8+KS+ participants because they expired before availability of effective, combination ART.
Plasma viral loads
HHV-8, CMV and EBV DNA loads in plasma were measured using laboratory-developed quantitative real-time polymerase chain reaction (PCR) assays. 17 –19 HIV viral RNA plasma loads were determined using a commercial real-time PCR assay (Roche Ultrasensitive RNA PCR assay).
T cell phenotyping
The number of CD3+, CD4+, and CD8+ T cells was determined by flow cytometry. 20 To determine the frequency of Treg, PBMC were fluorescently labeled using monoclonal antibodies and assessed for surface expression of CD4-V450, CD8-PerCyCy5.5, CD3-APC-H7, and CD25-APC (BD Biosciences), and intracellularly for FoxP3-PE (eBiosciences) by flow cytometry. Events were acquired with an LSR-II 12-color flow cytometer (BD) and then analyzed using FlowJo V 7.8 (TreeStar, Inc.). 21 The cytometer performance and optimal PMT voltages for individual channels were assessed by CS&T beads (BD) following the manufacturer's recommendation.
Treg depletion
PBMC were left untreated or depleted of Treg by magnetic bead separation according to the manufacturer's protocol (Miltenyi). The efficiency of Treg depletion was determined by flow cytometry, as described above.
HHV-8-specific T cell responses
To investigate the effect of Treg on CD8+ T cell function, PBMC were depleted of Treg 21 and compared to nondepleted PBMC after stimulation with viral peptide pools. Interferon-γ (IFN-γ) production was measured by ELISpot assay. 22 The HHV-8 lytic peptide antigen pool consisted of 6 HLA A*0201 epitopes: gB159–168, 12 gB492–500, 22a gB736–745, 12 K8.173–81, 12 K8.1135–143, 23 and K8.1209–217 23a (Sigma). The HHV-8 latent antigen peptide pool consisted of 9 HLA A*0201 epitopes: LANA-1140–148, 12 LANA-1238–246, 8 LANA-1281–289, 12 LANA-1417–425, 12 LANA-1688–697, 12 LANA-1920–928, 12 LANA-11116–1124, 8 K1217–25, 23a and K1223–32 12 (Sigma). Responses were compared to a pool of 12 HLA A*0201 immunodominant epitopes from HIV p1777–85, p2p7p1p670–79, p2419–27, PR76–84, RT33–41, RT179–187, RT309–317, Vpr59–67, gp160311–320, gp160813–822, Nef136–145, Nef180–189 24 (Sigma), and a CEF (CMV, EBV and influenza A virus [Flu]) peptide pool consisting of 32 MHC I-restricted, CD8+ T cell epitopes from CMV, EBV, and Flu 25 (NIH AIDS Reagent Program). Cells treated without peptides in RPMI 1640 medium (GIBCO) supplemented with 10% heat-inactivated FCS were used as a negative control. PBMC or PBMC depleted of Treg (100,000 cells/well) were stimulated with peptides (10 μg/mL) as previously described. 26 The results were a net number of virus-specific spot-forming cells per 106 PBMC in peptide-treated cell cultures minus control cells, with positive responses defined as greater than the mean number of spots in the control wells plus one SD.
Statistics
Data were analyzed by two-way ANOVA and Student's t-test using GraphPad Prism software v6.5.
Results
T cell numbers and viral load in relation to KS
Participants who were coinfected with HIV and HHV-8 and developed KS (HIV+HHV-8+KS+) had a downward trajectory of CD3+ (Fig. 1A), CD8+ (Fig. 1B), and CD4+ (Fig. 1C) T cell counts beginning 5.7 years before development of KS, reaching significance in the HIV+HHV-8+KS+ men for CD4+ T cell counts at 1.8 years before KS (Fig. 1C) and for CD3+ and CD4+ T cell counts at the time of KS diagnosis (Fig. 1A and 1C; 0.1 year). T cell counts for the HIV+HHV-8− participants were comparable to those of the HIV+HHV-8+KS− participants over the 5.7 years of the study. CD4+ T cell counts improved after ART in the HIV+HHV-8+KS− and the HIV+HHV-8− groups, although they do not recover to the levels seen in the HIV−HHV-8+ or HIV−HHV-8− groups taken at a contemporaneous time point. None of the KS+ subjects survived to receive ART, and thus, the effects of ART on these participants were not assessable in this study.
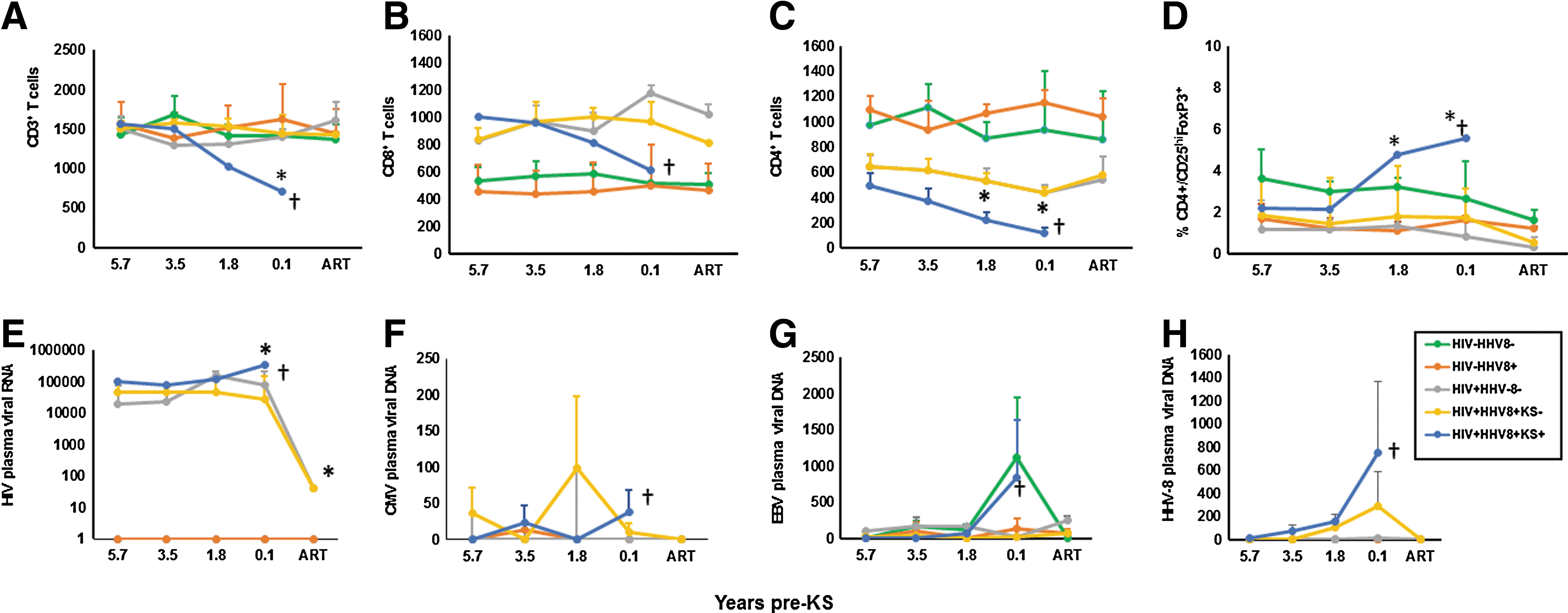
T cell counts and viral loads in study participants. Five HIV−/HHV-8− (green lines), 5 HIV−HHV-8+ (orange lines), 5 HIV+HHV-8− (gray lines), 10 HIV+HHV-8+KS− (yellow lines), and 5 HIV+HHV-8+KS+ (blue lines) participants were selected from the Pittsburgh MACS clinical site. Five visits were chosen based on specimen availability at the time of clinical diagnosis of KS in the HIV+HHV-8+KS+ participants and the corresponding visits in the HIV−HHV-8−, HIV−HHV-8+, HIV+HHV-8−, and HIV+HHV-8+KS− participants (X axis = median years pre-KS). Data are mean (±SE)/mm3 CD3+
Percentages of Treg in blood (CD4+CD25hiFoxP3+) were similar in all participants at 5.7 and 3.5 years of study, with a significant increase at 1.8 and 0.1 years (p < 0.05) before KS in the HIV+HHV-8+KS+ participants (Fig. 1D). Initiation of ART had no significant effect on the percentage of Treg in the HIV+HHV-8+KS− and HIV+HHV-8− participants (Fig. 1D).
Significant increases in plasma HIV viral load were noted in those men who developed KS compared to those who did not, at the time of KS diagnosis (0.1 year; p < 0.05) (Fig. 1E). The HIV viral load significantly decreased in the HIV+HHV-8− and HIV+HHV-8+KS− participants when they were on ART.
Low plasma viral loads for CMV (Fig. 1F) and EBV (Fig. 1G) were periodically detectable over the time course of this study in the various participants, with no discernable significant trends.
HHV-8 was not detected in plasma of the HIV−HHV-8+ participants (Fig. 1H). HHV-8 viral load trended higher in the HIV+HHV-8+KS+ and HIV+HHV-8+KS− participants over the pre-ART course of the study, but was not significantly different among the pre- and post-KS time point values. HHV-8 in the blood of the HIV+HHV-8+KS− participants decreased to undetectable levels while on ART.
CD8+ T cell reactivity to HHV-8 in relation to KS: cumulative responses
We first assessed the cumulative, composite results for CD8+ T cell responses to HHV-8 lytic and latent peptides in the three HHV-8+ groups. We found that without removing Treg from the PBMC, CD8+ T cell responses to both types of HHV-8 antigens were relatively low and did not significantly differ (Fig. 2A, HHV-8 latent and lytic peptides; black bars). Removal of Treg resulted in significantly higher responses to both lytic and latent HHV-8 peptides only in the HIV+HHV-8+KS− group (Fig. 2A, HHV-8 latent and lytic peptides, gray compared to black bars). When we measured the cumulative T cell responses to pooled HHV-8 latent and lytic antigens, these responses were lower in PBMC of HIV+HHV-8+ participants who did or did not develop KS compared to the HIV−HHV-8+ controls, although this was not significant (Fig. 2B, black bars).
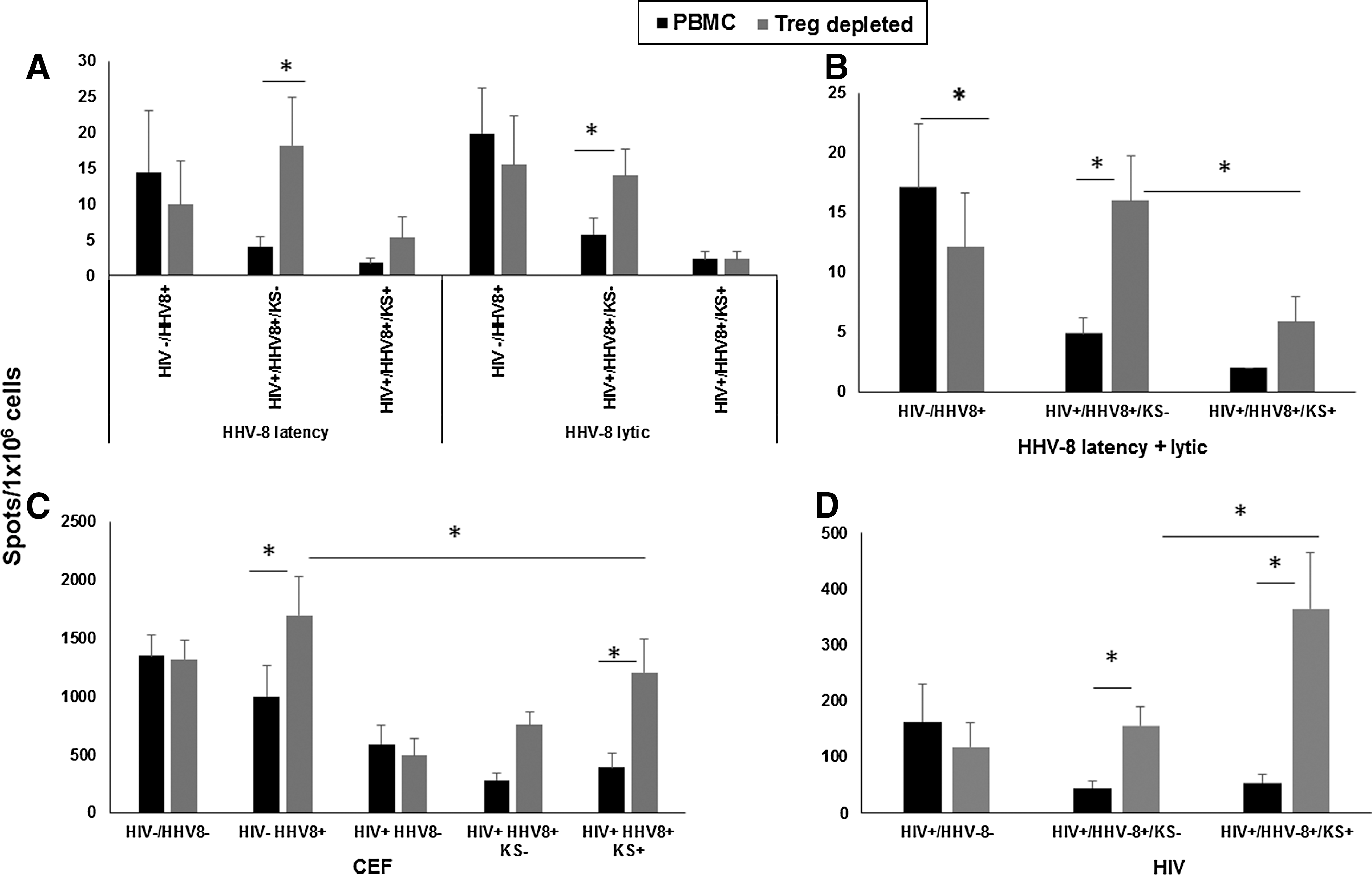
Effect of Treg on cumulative anti-HHV-8 CD8+ T cell responses. Cells were tested in an ELISpot assay against pools of HHV-8 latency or lytic peptides, CEF peptides, and HIV peptides. Data are mean IFN-γ spots (±SE)/1 × 106 cells.
Removal of Tregs from the PBMC resulted in a significant increase in the cumulative T cell responses to both HHV-8 latent and lytic antigens compared to non-Treg-depleted responses in HIV+HHV-8+KS− participants, but not in HIV−HHV-8+ controls or HIV+HHV-8+KS+ participants (Fig. 2B, gray compared to black bars). T cell responses measured in the Treg-depleted samples were also significantly higher in the HIV+HHV-8+KS− when compared to the same treatment in the participants who developed KS (Fig. 2B, gray bars).
PBMC from the 3 HHV-8+ study groups responded 32- to 228-fold higher to CEF antigens than to the HHV-8 antigens (Fig. 2C, CEF, black bars). T cell responses in the Treg depleted cultures to CEF in the HIV+HHV-8+KS+ group were significantly lower than in the HIV−HHV-8+ control group (Fig. 2C, CEF, gray bars).
T cell reactivity after removal of Treg was significantly higher to CEF in HIV−HHV-8+ controls and in the HIV+HHV-8+KS+ group (Fig. 2C, CEF, gray compared to black bars). Removal of Treg in both the HIV+HHV-8+KS− and HIV+HHV-8+KS+ groups did not restore the T cell responses to CEF to the same levels seen in the HIV- HHV-8 + control group.
T cell reactivity to the HIV peptide pool in PBMC was ∼10-fold greater than reactivity to HHV-8 antigens (Fig. 2D, HIV peptide pool, black bars). Removal of Treg enhanced T cell responses to HIV in the both the HIV+HHV-8+KS− and HIV+HHV-8+KS+ groups (Fig. 2D, black compared to gray bars), and was higher in the HIV+HHV-8+KS+ compared to the HIV+HHV-8+KS− groups (Fig. 2D, gray bars).
Taken together, these cumulative results indicate that CD8+ T cell reactivity to HHV-8 lytic and latent peptide epitopes was lower compared to CEF and HIV antigens, that is, an average of 32- and 10-fold lower, respectively, in the HIV+HHV-8+KS− and an average of 288- and 23-fold lower, respectively, in the HIV+HHV-8+KS+. Moreover, enhanced Treg activity was associated with lower CD8+ T cell reactivity to HHV-8 in the HIV+HHV-8+KS− group but not in the other HHV-8-positive groups.
CD8+ T cell reactivity to HHV-8 in relation to KS: longitudinal responses
We next analyzed the longitudinal T cell responses to HHV-8 based on the time of KS development. There was no difference between T cell responses to lytic and latent HHV-8 antigens in relation to development of KS (data not shown); therefore, these results were combined for further analysis. T cell reactivity to HHV-8 antigens without removal of Treg was low throughout the median 5.7 years of follow-up in the three HHV-8+ groups, that is, HIV−HHV-8+ controls and HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants (Fig. 3B, D, and E, black bars). Although before ART, increases in T cell responses to HHV-8 were consistently noted after removal of Tregs in the HIV+HHV-8+KS− participants (Fig. 3D), and also at all time points in HIV+HHV-8+KS+ participants (Fig. 3E), this enhancing effect was only significant at the earliest, 5.7-year time point in the HIV+HHV-8+KS− participants (Fig. 3D). Staining for CD4+ and CD8+ T cell frequency confirmed that this effect was not attributable to a proportional increase in the number of CD8+ T cells after removal of Treg (data not shown).
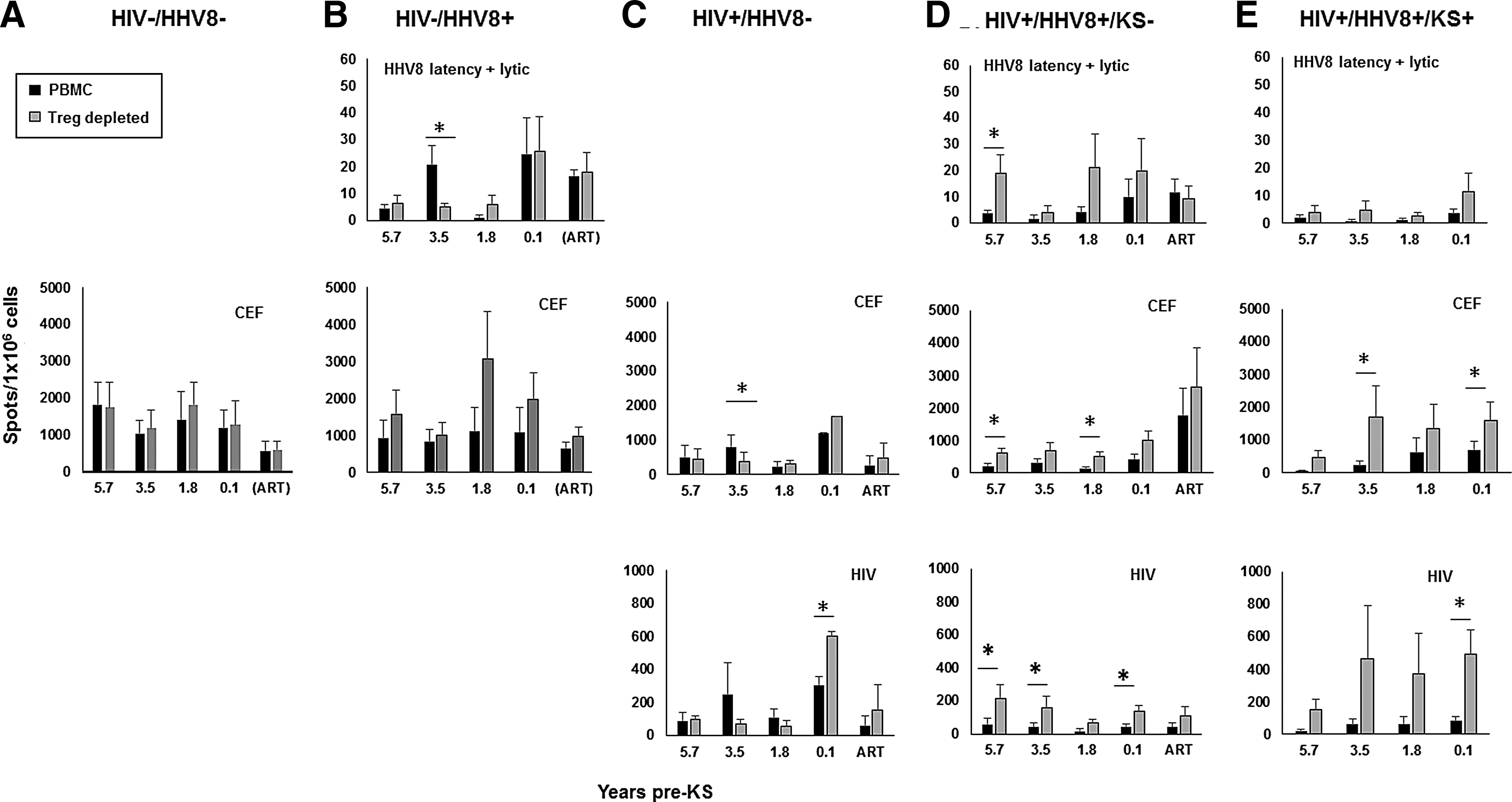
Effect of Treg on nonconcurrent, longitudinal antiviral CD8+ T cell responses. Study participants and visits were selected as detailed in Figure 1 legend (X axis = median years). Cells were tested in ELISpot assays in response to pools of HHV8 latency or lytic peptides, CEF peptides, and HIV peptides. Data are mean IFN-γ spots (±SE)/1 × 106 cells in 5 HIV−HHV8− participants
T cell reactivity to CEF antigens in PBMC without removal of Treg was an average of 177-fold higher than that to the HHV-8 antigens over the 5.7 years, with a consistent, upward directionality at each matched time point in the HIV−HHV-8+ (Fig. 3B, CEF) and to some extent in the HIV− HHV-8− controls (Fig. 3A, CEF), HIV+HHV-8+KS− participants (Fig. 3D, CEF), and HIV+HHV-8+KS+ participants (Fig. 3E, CEF). Removal of Treg had a consistent directionality pattern of increasing T cell responses to CEF in the HIV−HHV-8+ controls and HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants, reaching significance at several time points in the latter 2 groups (Fig. 3D and E; CEF). This consistent enhancing effect on CEF responses was not observed in the HIV+HHV-8− participants (Fig. 3C). A similar pattern of T cell reactivity in PBMC with and without Treg was noted to HIV peptides in the HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants (averaging 31-fold greater than the response to HHV-8 antigens), with consistent increases in directionality after removal of Treg (Fig. 3D, E).
Discussion
CD8+ T cell responses to HHV-8 lytic and latent proteins are believed to play a major role in controlling HHV-8 infection and preventing KS. 5,8,27 In the present study, there were no differences in CD8+ T cell reactivity to six HHV-8 lytic or nine HHV-8 latent MHC class I epitopes among the three HHV-8-infected groups, that is, HIV−HHV-8+ controls and HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants. Notably, mean levels of HHV-8-specific CD8+ T cell production of IFN-γ to both lytic and latent peptides were low in each of these three groups of HHV-8-positive participants. Indeed, the T cell response to these HHV-8 peptide epitopes was many fold lower than that to CEF and HIV peptide epitopes throughout the 5.7 years of study in the HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants. Furthermore, we did not observe significant changes in these CD8+ T cell responses to HHV-8, CEF, or HIV antigens during ART in the two groups that were testable while on ART, that is, HIV+HHV-8+KS− and HIV+HHV-8−. Thus, our results support that CD8+ T cell reactivity is relatively low against both lytic and latency antigens of HHV-8, regardless of HIV-1 coinfection or KS development, using conventional, short-term, antigen-stimulation methods. 13,28
We hypothesized that the low CD8+ T cell immune response to HHV-8 antigens could, in part, be due to overly responsive Treg. These cells normally function to maintain peripheral T cell homeostasis, 29 but pertinent to our study, are known to suppress CD8+ T cell responses to a murine gammaherpesvirus 14 and to the other human gammaherpesvirus, EBV, 15 and have been linked to enhancement of EBV-associated neoplasias. 30 –32 Thus, we postulated that excessively reactive Treg could be inhibiting anti-HHV-8 CD8+ T cell reactivity during HHV-8 infection. In fact, after removal of Treg, cumulative T cell IFN-γ responses were significantly increased in HIV+HHV-8+KS− participants, but not in HIV+HHV-8+KS+ participants, to both HHV-8 lytic and latent antigens. The consistently low anti-HHV-8 CD8+ T cell reactivity in the HIV+HHV-8+KS+ participants following Treg depletion over the 5.7 years of follow-up was not related to differences in the percentages of Treg, which significantly increased above the levels in the HIV+HHV-8+KS− participants at 1.8 years before and at the time of diagnosis of KS. Moreover, there was no discernable link of anti-HHV-8 CD8+ T cell responses to either HHV-8 or HIV viral loads in blood, with only the latter reaching significantly higher levels in the HIV+HHV-8+KS+ participants, compared to the HIV+HHV-8+KS− and HIV+HHV-8− groups, at the time of KS diagnosis. Removal of Treg consistently enhanced T cell responses to the CEF and HIV peptide pools in the HIV+HHV-8+KS− and HIV+HHV-8+KS+ participants. This enhancement was not found to CEF in the HIV−HHV-8− and HIV+HHV-8− controls, or to HIV in the HIV+HHV-8− controls. These results suggest that Treg can selectively dampen anti-HHV-8 responses in HIV- and HHV-8-coinfected individuals who did or did not develop KS. In addition, although we could not assess the effects of ART on our participants who developed KS, a pause in ART and rebound in HIV-1 replication are related to augmentation of the Treg subpopulation. 33 Therefore, a shift of the declining CD4+ T cell population to a more prominent Treg phenotype in KS cases before ART could be a final attempt to dampen exaggerated immune activation on the increase in HIV production associated with the onset of AIDS.
A limitation in our study is that we only assessed five men who developed KS and were available for analysis. This work was also restricted by our use of HHV-8 T cell epitopes associated only with HLA A*0201 and not other MHC class I alleles, due to their previously established antigenic specificity. Finally, as this study examined responses over many years, the immunologic patterns could, in part, be an effect of aging, which alters the function and phenotype of T cells. 34
In conclusion, our study of CD8+ T cell reactivity in HHV-8- and HIV-coinfected participants indicates that there were persistently low CD8+ T cell IFN-γ responses to HHV-8 lytic and latent proteins both in those men who developed KS and those who did not, beginning 5.7 years before KS. Although there was an increase in the percentage of Treg in the blood late in the course of HHV-8 and HIV coinfection in those men who developed KS, removal of Treg had a mild enhancing effect on HHV-8-specific T cell responses in HHV-8- and HIV-coinfected participants who did and did not develop KS. Regulatory factors in addition to this form of Treg could be effectively suppressing anti-HHV-8 CD8+ T cell reactivity preceding HIV-associated KS.
Footnotes
Acknowledgments
We thank A. Bullotta, Dr. B. Macatangay, L. Borowski, Dr. Y. Chen, E. Knowlton, Dr. R. Mailliard, K. Hartle, B. Zeher, and M. Jais for technical assistance, the Center for Analysis and Management of MACS Data, Johns Hopkins University, for data management, and the Pitt Men's Study MACS staff and volunteers for their dedication and support. This work was supported by the National Institutes of Health grants R01 CA82053, U01 AI35041, and T32 AI065380.
Author Disclosure Statement
No competing financial interests exist.