
Abstract
Carriage of alleles encoding certain inhibitory natural killer (NK) cell receptor/HLA ligand KIR3DL1/HLA-B combinations is associated with protection from HIV infection and slow time to AIDS, implicating NK cells in HIV control. NK cells also mediate antibody-dependent cellular cytotoxicity (ADCC). ADCC has been identified as a correlate of protection in secondary analyses of the modestly protective RV144 Thai HIV vaccine trial. In ADCC, HIV envelope (Env)-specific antibodies (Abs) bridge HIV-infected or gp120-coated target cells and NK cells expressing CD16 receptors for Ab Fc domains. CD16 engagement activates NK cells to secrete cytokines/chemokines, degranulate, deliver granzyme B (GrB) to target cells, and cytolysis. A subset of HIV+ subjects, known as slow progressors (SPs), maintains low-level viremia without treatment. HIV+ SPs versus progressors have higher titers and/or a greater breadth of ADCC-competent Abs. Investigations of the functional capacity of NK effector cells following CD16 engagement in HIV+ subjects are lacking. We used the ADCC-GranToxiLux (ADCC-GTL) assay to assess the frequency of GrB+ (%GrB+) cells generated by effector cells from 37 HIV+ SPs and 15 progressors to gp120-coated CEM.NKr.CCR5 target cells in the presence of anti-Env Abs. Subject groups were stratified according to whether or not they carried educating KIR3DL1/HLA-B combinations able to confer NK cells with functional potential. No differences were observed in %GrB+ target cells generated by effector cells from carriers of educating versus noneducating KIR3DL1/HLA-B pairs. The absence of an effect of NK cell education on this readout may be due to loss of the ability of educated NK cells from SPs to respond to Ab-dependent stimulation and/or the lower frequency of KIR3DL1+ than KIR3DL1− NK cells that coexpress CD16. That KIR/HLA genotypes have minimal impact on interindividual differences in ADCC potency has relevance for therapeutic interventions that target ADCC for HIV control.
Introduction
I
ADCC activity depends not only on the presence of ADCC-competent Abs but also on the ability of effector cells to mediate this function. 13 By using a single source of HIV Env-specific Ab in ADCC assays with effector cells from different donors, it is possible to focus on factors that affect ADCC effector function and their consequences on ADCC targets. Several flow cytometry-based assays have been developed for this purpose. Some measure the activation of NK cells to secrete chemokines/cytokines and/or express CD107a following stimulation with either HIV-infected CD4+ T cells or infected or recombinant HIV Env gp120 (rgp120) or gp140-coated CEM.NKr.CCR5 (CEM) target cells in the presence of HIV-specific Abs. 14 –19 Others measure the delivery of granzyme B (GrB) to target cells or their lysis by NK cells activated in an Ab-dependent manner. 18,20 –22
In uninfected persons, NK cell education influences the level to which NK effector cells are activated by Ab-dependent stimuli to produce CD107a and IFN-γ. 14,18,19 On the other hand, the frequency of GrB+ target cells generated in an ADCC-GranToxiLux (ADCC-GTL) assay was independent of NK cell education through KIR3DL1 (hereafter 3DL1). 18 In HIV-infected individuals, chronic progressive infection abolished Ab-dependent NK cell responsiveness, which may be partly due to the reduction of Fc receptors on effector cells. 14,23 –27
A subset of HIV-infected subjects experiences viral control and a slow disease course without treatment. 28 These slow progressors (SPs) include elite controllers (ECs) with VLs below 50 HIV RNA copies/ml of plasma detectable by standard VL assays and viral controllers who maintain low, but detectable, HIV VLs. 29,30 While several reports have found higher titers and/or a greater breadth of ADCC-competent Abs in plasma from HIV-infected SPs than typical progressors (TPs), 21,31 –34 less is known regarding the functional capacity of NK effector cells from SPs with respect to their ability to respond to Fc-dependent stimuli leading to the delivery of GrB to target cells.
In this study, we used the ADCC-GTL assay to investigate the delivery of GrB by effector cells from HIV-infected SPs and progressors in the chronic phase of infection to gp120-coated CEM cells in the presence of HIVIG, a pool of plasma from asymptomatic HIV-infected donors. We assessed whether NK cell licensing through the 3DL1 inhibitory receptor had an impact on the delivery of GrB to target cells as well as on the Fc-dependent activation of NK cells. We also examined the frequency of CD16+ cells among NK cells expressing or not the inhibitory 3DL1 receptor.
Materials and Methods
Study population
The study population included 52 treatment-naïve HIV-infected individuals in the chronic phase of infection, of which 37 were SPs and 15 TPs. The SP subjects were from the Canadian Cohort of HIV-Infected SPs recruited from sites across Canada, including Montreal, Toronto, and Vancouver. 30 All SPs maintained VLs below 4,000 copies/ml of plasma. Of these, 12 were ECs with VLs below the limit of detection throughout follow-up using the real-time HIV-1 assay (Abbott Molecular, Des Plaines, IL). Both SPs and TPs were followed longitudinally. Information on CD4 and CD8 counts and VL at the time point used to generate the results presented in this article is provided for each subject in Table 1. TPs were enrolled in the Montreal Primary Infection (PI) cohort, which follows individuals who are recruited within 12 months of infection for up to 4 years, an average of once every 3 months. All were followed long enough to be able to calculate a VL set point based on at least three observations during chronic phase infection; their VL at the time point tested was within the range of their VL set point. Table 1 also provides information on the HLA type, Bw4/Bw6 genotype category, and 3DL1 allotype of each study participant. Individuals were assigned to the Bw4 group if they carried at least 1 Bw4 allele. Those belonging to the Bw6 group carried no Bw4 alleles at either the HLA-A or -B loci. Table 2 shows the median and interquartile range (IQR) for the CD4 and CD8 counts of the SP and TP populations and of the VL for the 26 VC and 15 TP subjects.
*h/*y: 3DL1 hmz genotype with no low-expression 3DL1 alleles; *l/*x: 3DL1 hmz genotype with at least 1 low-expression 3DL1 allele.
CD4 and CD8 counts are reported in cells/mm3.
VL, viral load, reported as RNA copies/ml of plasma.
Null refers to carriage of 2 copies of the unexpressed 3DL1 allele *004.
VLs >40 copies/ml plasma were below the limit of detection of the assay used.
This subject had a copy number variation at the 3DL1 locus and expressed *001, *004, and *022 alleles.
HTZ = Heterozygous for a 3DL1/3DS1 generic genotype.
Results are presented as median (IQR).
IQR, interquartile range.
Genotyping
All individuals were typed for MHC-I alleles by sequence-based typing using kits from Atria Genetics, Inc. (South San Francisco, CA). Assign 3.5+ software was used to interpret sequence information for allele assignment (Conexio Genomics, Perth, Australia). Genotyping and allotyping of the KIR3DL1/S1 locus were performed as previously described. 35,36 3DL1 allotyping was done by sequencing 3DL1 exons. 35 Small nucleotide polymorphisms corresponding to 3DL1 alleles were identified by aligning the sequenced DNA to a reference consensus sequence consisting of a contig of 3DL1 cDNA sequences.
Cells
Peripheral blood mononuclear cells (PBMCs) were isolated from blood by density gradient centrifugation (Ficoll-Paque; Pharmacia, Uppsala, Sweden) and cryopreserved in 10% dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO) with 90% fetal calf serum (FCS; Wisent, Inc., St. Bruno, QC, Canada). After thawing, cells were maintained in RPMI media containing 2 mM L-glutamine; 100 IU/ml penicillin; 100 μg/ml streptomycin; and 10% FBS (all from Wisent) (R10) at 37°C in a 5% humidified CO2 incubator at a concentration of 1 × 106 cells/ml. CEM cells were obtained through the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH, from Dr. Alexandra Trkola.
ADCC-GTL assay
The ADCC-GTL assay was performed in triplicate as previously described on ex vivo frozen and thawed overnight-rested PBMCs. 18 Briefly, recombinant gp120 (rgp120, HIV-1BaL NIH AIDS Research and Reference Reagent Program, Division of AIDS, DAIDS, NIAID)-coated CEM target cells were labeled with TFL4 (OncoImmunin, Inc., Gaithersburg, MD) and a viability marker (NFL1; OncoImunnin) for 15 min at 37°C. Effector and target cells were cocultured with the GrB substrate at a ratio of 30:1. Following incubation for 5 min at RT, HIVIG, purified IgG isolated from pooled plasma obtained from HIV-infected donors (NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH: Catalog #3957, HIV-IG from NABI and NHLBI), was added (or not, as a control) at a dilution of 1:1,000 and incubated for 15 min at RT. The samples were then pelleted by centrifugation and incubated for 1 h at 37°C in a humidified 5% CO2 incubator. After washing, the cells were resuspended in wash buffer and acquired with a calibrated LSRFortessa flow cytometer (BD Biosciences, San Jose, CA) using a high-throughput system (HTS) (BD Biosciences). CEM cells alone were used to determine the position of the target cell population within the forward scatter (FSC) and side scatter (SSC) plot. The percent of GrB-positive (%GrB+) cells was then determined by gating on the live (NFL−) target cell (TFL+) population. Data analysis was performed using FlowJo software version 9.8 (Tree Star, Inc., Ashland, OR).
Ab-induced NK cell activation assay
PBMCs isolated as described above were rested overnight in R10. The following day, PBMCs and rgp120-coated CEM cells (prepared as described above) were cocultured at a 10:1 ratio in 96-well plates in the presence or absence of a 1:1,000 dilution of HIVIG for 6 h at 37°C in a humidified 5% CO2 incubator. Anti-CD107a-PE-CF594 (BD) was added at the start of the stimulation; brefeldin A (5 mg/ml; Sigma-Aldrich) and monensin (6 mg/ml, Golgi Stop; BD) were added 30 min after the initiation of the coculture. Cells were stained for viability using the UV Live/Dead fixable dead cell stain kit (Invitrogen, Burlington, ON, Canada) and for surface markers using the following Abs: CD3-BV785 (OKT3), CD56-BV711 (NCAM16.2), and KIR3DL1-BV421 (DX9) (all from BioLegend). Samples were washed twice with phosphate-buffered saline (PBS) containing 1% FBS (Wisent), fixed and permeabilized using a Fix/Perm kit (Invitrogen) according to the manufacturer's directions, and stained for intracellular IFN-γ and CCL4 using anti-IFN-γ-BV605 (B27; BD) and anti-CCL4-APC (24006; R&D Systems, Minneapolis, MN) Abs.
Samples were washed, fixed with 1% paraformaldehyde (PFA, Santa Cruz Biotechnology, Inc., Dallas, TX), and between 300,000 and 1,000,000 total events were acquired for each sample on a calibrated LSRFortessa flow cytometer (BD). Single-stained control beads (CompBead; BD) were used in every experiment to calculate compensation. Flow cytometry analysis for NK cell activation following stimulation was performed using FlowJo software. The results for the percent of functional NK cells stimulated by rgp120-coated CEM cells in the presence of HIVIG reported were background corrected using samples to which no HIVIG was added. Boolean gating was used to identify the frequency of NK cells with seven possible functional profiles: trifunctional (CD107a+IFN-γ+CCL4+), bifunctional (any combination of two functions), and monofunctional. Total responsiveness was defined as the sum of frequencies of tri-, bi-, and monofunctional NK cells. Total CD107a, IFN-γ, or CCL4 expression was defined as the sum of all functional subsets that included these functions.
Phenotypic staining
PBMC samples from 13 subjects whose cells were used as effector cells in ADCC-GTL assays were also cell surface stained to determine the frequency of Boolean-gated 3DL1+/− CD16+/− subsets among the CD3−CD56dim NK cell population. The surface staining was performed on frozen and thawed overnight-rested PBMCs on an aliquot of cells separate from the one used as effectors cells for the ADCC-GTL assay. Staining was done in the dark on ice on 1 × 106 PBMCs per individual with the following Abs: CD3-PerCP (OKT3), CD56-APC (NCAM16.2), CD158e1/KIR3DL1-PE (DX9), and CD16-pacific blue (all from BioLegend, San Diego, CA). Stained cells were then washed and fixed with 1% PFA (Santa Cruz). Between 300,000 and 400,000 events per sample were acquired using the FACSCanto flow cytometer (BD). Data analysis was performed using FlowJo software. Unstained and single-color controls were used for each subject for multicolor compensation and gating purposes.
Statistical analysis
Statistical analysis and graphical presentation were performed using GraphPad InStat 3.05 and GraphPad Prism 6. Wilcoxon matched pairs tests were used to compare the effect of a condition on two matched groups. Mann–Whitney and Kruskal–Wallis tests with Dunn's post-test comparisons were used to compare the significance of differences between two or more than two unmatched groups, respectively. p-values less than .05 were considered significant.
Results
ADCC activity of cells from HIV-infected SP and TP subjects
The gating strategy used to determine the %GrB+ cells generated in the ADCC-GTL assay is shown in Supplementary Figure S1 (Supplementary Data are available online at
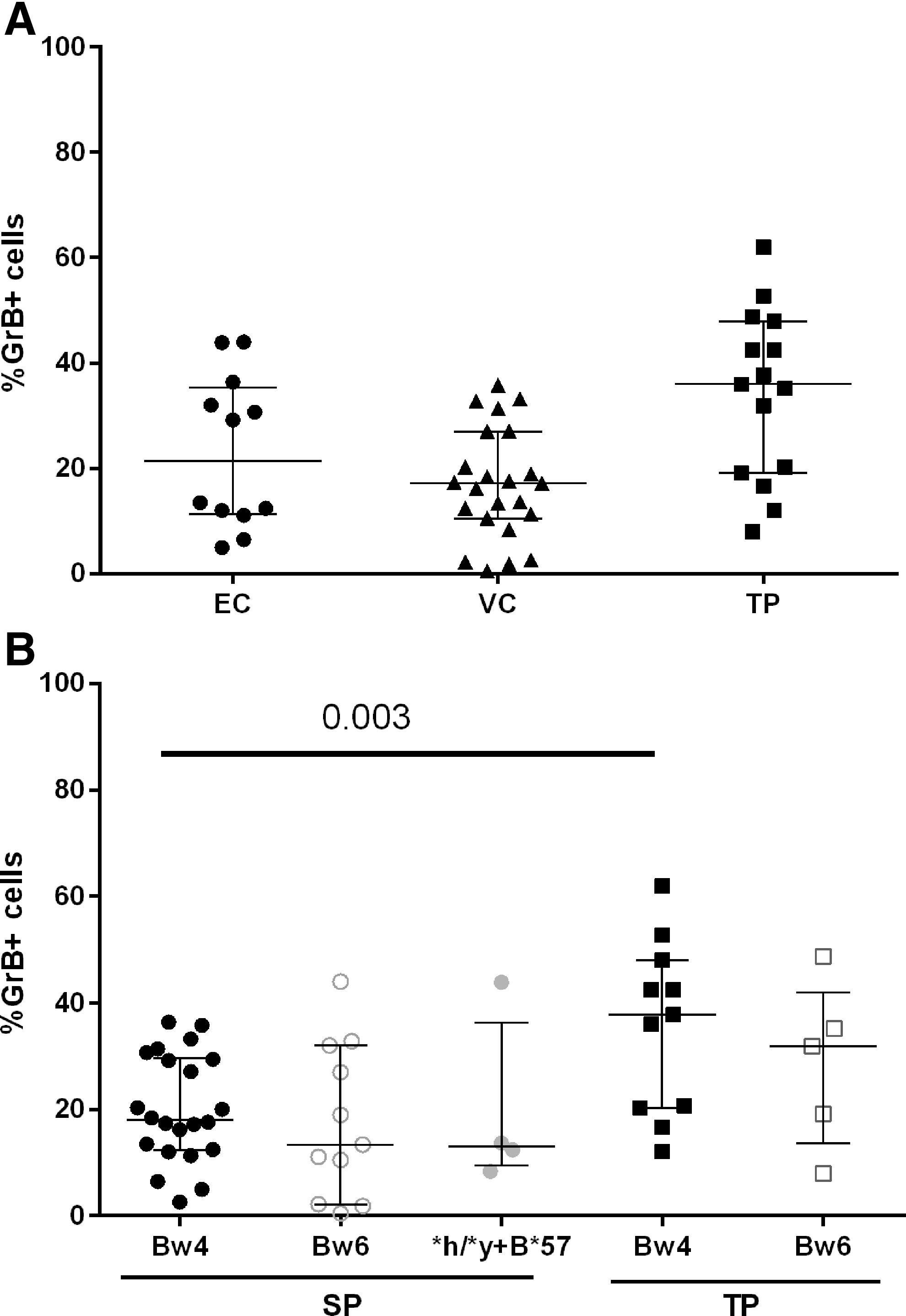
The influence of NK cell education through 3DL1 receptors on the frequency of granzyme B-positive (%GrB+) CEM target cells generated in the ADCC-GTL assay. The y-axis shows the %GrB+ cells generated in the ADCC-GTL assay by effector cells from three 3DL1 hmz populations: HIV-infected ECs (n = 12), VCs n = 25, and TPs (n = 15). HIV-infected SP (n = 37, subject 1 through 37 in Table 1) and TP (n = 15) individuals
To assess the impact of education through 3DL1-Bw4 on ADCC responses generated by effector cells from HIV-infected subjects, we stratified the results for %GrB+ CEM targets according to whether effector cells were from study subjects belonging to the Bw4 or Bw6 groups. Since we previously showed that the *h/*y+B*57 3DL1 homozygous (3DL1 hmz)/HLA-Bw4 genotype supported particularly potent education, we also considered results from carriers of this KIR/HLA combination separately from those generated by carriers of other 3DL1 hmz/HLA-Bw4 combinations. 37 –39 For this analysis, we combined the EC and VC groups into one SP population since PBMCs from both groups had generated similar results in the ADCC-GTL assay. There were no significant differences for the %GrB+ cells generated by effector cells from SP individuals belonging to the Bw4, Bw6, or *h/*y+B*57 groups (p = .6, Kruskal–Wallis) or for the TPs belonging to the Bw4 or Bw6 groups (p = .36, Mann–Whitney test) (Fig. 1B). Comparison of results from SPs and TPs who belonged to the Bw6 groups did not differ significantly from each other (p = .41, Mann–Whitney test). Cells from Bw4 TPs generated higher frequencies of GrB+ CEM cells than did cells from Bw4 SPs (p = .003, Mann–Whitney). We also investigated the amount of GrB delivered to CEM cells depending on whether the effector cells were from SP Bw4 versus Bw6 groups by ascertaining the mean fluorescence intensity of GrB staining and found no between-group differences (not shown). In summary, these results show that education through 3DL1/Bw4 pairs influenced neither the frequency of GrB+ target cells generated by effector cells from SPs and TPs nor the intensity with which GrB is delivered to target cells.
Comparison of the frequency of NK cells from SPs responding to Ab-dependent stimulation
HLA-Bw4 is a ligand for the 3DL1 receptor on NK cells involved in licensing 3DL1+ NK cells. Others have shown that 3DL1+ and 3DL1− NK cells from HIV-uninfected persons carrying this licensing pair were differentially stimulated by HLA-null cells and by Fc-dependent stimuli such as rgp120-coated CEM in the presence of anti-gp120 Abs. 14,18,19 The effect of NK cell education on NK cell activation by HLA-null cells was abolished when the responding cells were from HIV-infected individuals in chronic phase infection. 14 We questioned whether 3DL1+ NK cells from SP Bw4 carriers retained their functional advantage over 3DL1− NK cells in terms of their responsiveness to Fc-dependent activation. The gating strategy used to detect NK cells, 3DL1+/− cell populations, and their functional subsets is shown in Supplementary Figure S2. The frequency of 3DL1+ and 3DL1− NK cells from five ECs responding to Fc-dependent activation did not differ significantly for total responsiveness (p = 1.0, Wilcoxon test) (Fig. 2), so too the frequency of 3DL1+ and 3DL1− NK cells secreting IFN-γ, CCL4, or expressing CD107a did not differ significantly (p = 1, .81 and .19, respectively, Wilcoxon) (Supplementary Fig. S3). Thus, the effects on NK cell functionality of education through 3DL1 engagement were abolished in HIV-infected SPs as has been reported for HIV-infected progressors in chronic phase infection. 14

NK cell functional responses to antibody-dependent activation. The y-axis shows the frequency of total functional NK cells within the KIR3DL1+ (3DL1+) and KIR3DL1− (3DL1−) NK populations from five elite controllers who are 3DL1 homozygotes and cocarry an HLA-Bw4 allele following stimulation with rgp120-coated CEM.NKr.CCR5 (CEM) target cells and HIVIG. The SP study subjects included in this analysis correspond to subjects 9, 11, 25, 26, and 37 in Table 1. The frequency of functional NK cells between NK subjects was not significant.
ADCC activity inversely correlates with VL in TPs, but not SPs
We next investigated correlations between the %GrB+ cells generated in the ADCC-GTL assay and either CD4 counts or VL in HIV-infected subjects. We found that the %GrB+ cells did not correlate with the CD4 counts in either SPs or TPs (Fig. 3A, C). Although no correlation was observed between %GrB+ cells and VL for effector cells from SPs (Fig. 3B), there was a significant inverse correlation between these parameters for effector cells from TPs (Fig. 3D, Spearman's r = −0.72, p = .02). Thus, at least within TP subjects, the frequency of GrB+ target cells generated in the ADCC-GTL assay was negatively associated with VL.

The %GrB+ cells generated in the ADCC-GTL assay is negatively correlated with HIV VL in TP subject. Correlation analyses of %GrB+ cells generated in the ADCC-GTL assay by effector cells from 37 HIV-infected SP
A lower frequency of 3DL1+ compared with 3DL1− NK cells are CD16+
CD16 is an activating receptor present on CD56dim NK cells, which is crucial for activating NK cells to mediate ADCC. The gating strategy used to compare the coexpression of CD16 on 3DL1+ versus 3DL1− NK cells is shown in Supplementary Figure S4. When we compared the frequency of 3DL1+CD16+ and 3DL1−CD16+ CD56dim NK cell populations in 13 SPs who belonged to the Bw4 group, we found that 3DL1+ cells coexpressed CD16 at a significantly lower frequency than did 3DL1− NK cells. (Fig. 4, p = .0002, Wilcoxon). Thus, 3DL1+ NK cells that have the potential to be educated through this receptor are less likely to be CD16+ cells than their 3DL1− counterparts.
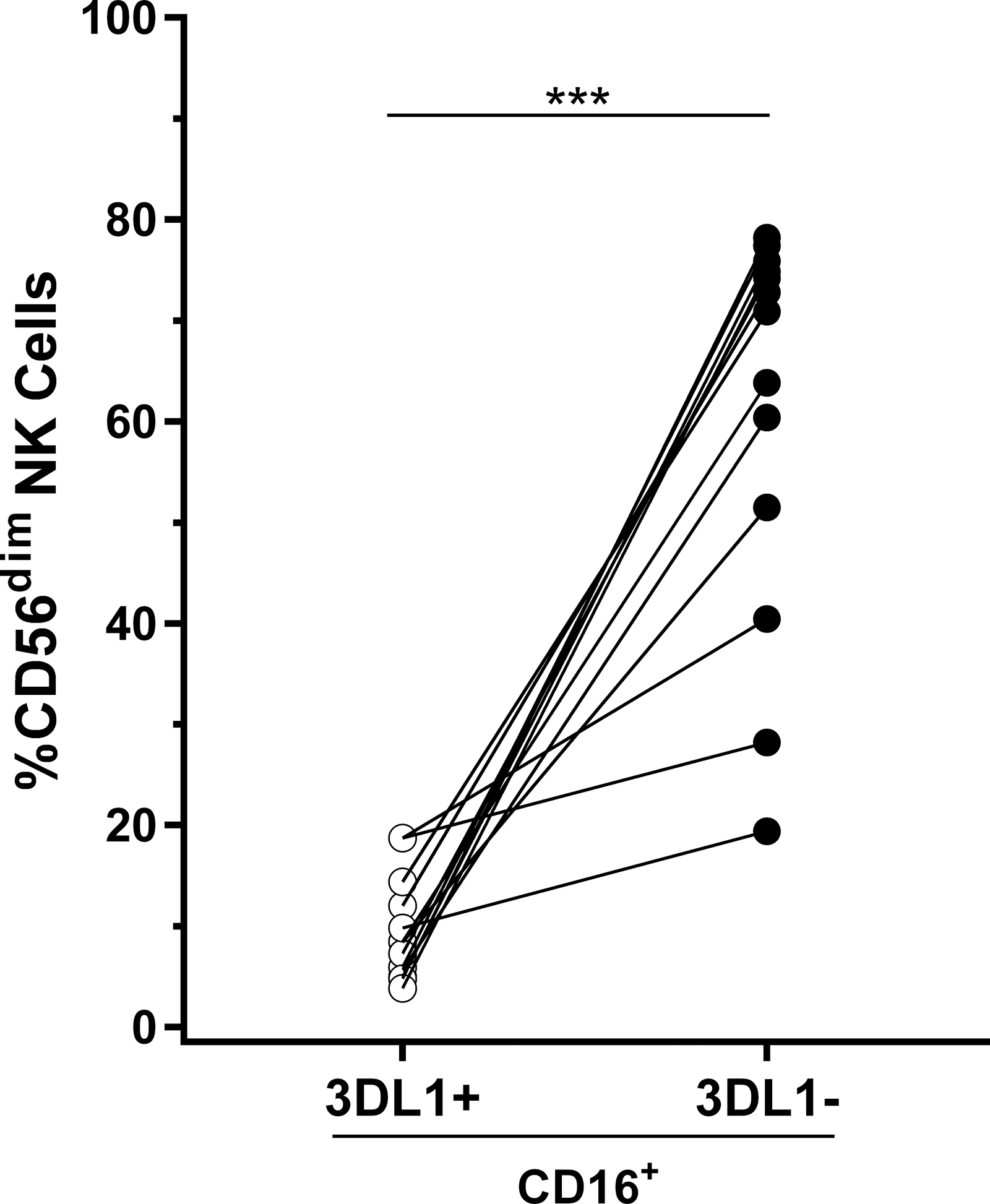
Frequency of CD16+ 3DL1−/+ NK cells among CD3−CD56dim NK cells. Shown on the y-axis is the frequency of CD16+ cells among 3DL1+ and 3DL1− CD56dim NK cells from 13 SPs belonging to the Bw4 group. Wilcoxon tests were used to assess the significance of within-individual differences between 3DL1+ and 3DL1+ NK cell populations. p-values for between-NK cell population differences are shown over the line linking the two groups. *** p < .001.
Discussion
In this article, we show that effector cells from treatment-naïve HIV-infected individuals categorized as ECs, VCs, and TPs generate percentages of GrB+ gp120-coated CEM target cells in an ADCC-GTL assay that are not significantly different from each other. Although there was variation in the %GrB+ cells generated in this assay within each subject group, differences were not due to the effect of NK cell licensing through 3DL1 receptors on effector cells. The %GrB+ cells generated by effector cells from the Bw6 TP and SP groups were similar, while effector cells from Bw4 TPs generated higher frequencies of %GrB+ cells than those from SPs. There was a significant negative correlation between %GrB+ cells and VL in TPs. There was no effect of NK cell education through the 3DL1 receptor on the extent to which NK cells, even when from ECs, responded to Fc-dependent activation by rgp120-coated CEM cells. We also observed that there was a lower frequency of CD16+ NK cells among 3DL1+ compared with 3DL1− NK cells from SPs.
GrB is a serine protease present within the lysosomal granules of effector cells such as cytotoxic T lymphocytes and NK cells. When effector cells are induced to degranulate, the granule contents are taken up by target cells. In the case of the ADCC-GTL assay, a GrB substrate is added that fluoresces when cleaved by GrB. By gating on live, labeled target cells, it is possible to enumerate the percentage of target cells, which have taken up GrB in the presence versus absence of ADCC-competent Abs. As GrB is one of the first active enzymes delivered to target cells and is part of the pathway leading to apoptosis, the GTL-ADCC assay measures an early marker for cytotoxicity. 20,40
A screen of a panel of Abs to various HIV Env epitopes on HIV-infected cells found that primary and laboratory-adapted HIV isolates differ in their susceptibility to neutralization and ADCC. It is notable that these Ab-mediated activities overlap extensively. 41 Primary isolates tend to be more resistant to neutralization and ADCC than the laboratory-adapted HIV-1NL4-3. This is likely related to the adaptations that HIV Env from primary isolates undergo to allow for viral replication under pressure exerted by virus-specific immunity. 42 –44 For example, the downmodulation of CD4 on HIV-infected CD4 T and CEM cells by HIV Nef and Vpu prevents the generation of the CD4-induced (CD4i) Env epitope recognized by some ADCC-competent Abs. 45 –47 Recently, shed gp120 released from infected cells and binding to uninfected bystander cells was shown to be recognized by the A32 Ab, which recognizes the CD4i epitope. 48 In this report, we were interested in investigating the responsiveness of NK cells to Fc-dependent activation and the consequences of this on the delivery of GrB to target cells. For this reason, we used rgp120-coated rather than HIV-infected CEM or autologous HIV-infected CD4+ T cells as target cells. The gp120 on the surface of coated CEM cells is recognized by HIVIG and plasma from HIV-infected subjects and by other ADCC-competent Abs, including those to CD4i epitopes. 18,21,45,46 Using a single source of pooled polyclonal anti-Env Abs in this assay eliminated the effect of Ab on intersubject variability, allowing us to focus on the impact of NK cell licensing on the delivery of GrB to CEM targets. Furthermore, the CEM target cell line expresses no Bw4 antigens, which can interact with 3DL1 on NK cells to suppress their function. 18,49 In summary, the ADCC-GTL assay parameters selected for this study were appropriate for investigating the influence of NK cell education through 3DL1 on the delivery of GrB to target cells by effector cells from HIV-infected individuals as well as on the activation of NK cell, cells from HLA-Bw4 carriers expressing or not 3DL1.
PBMCs from ECs, VCs, and TPs generated frequencies or GrB+ target cells that did not differ significantly from each other. Although others have noted that chronic untreated HIV infection has a negative impact on NK cell activity, the TP population studied here was from subjects in the 2nd or 3rd year postseroconversion. 50 In a previous study, we examined the %GrB+ cells generated using effector cells from 50 HIV-uninfected subjects using the same ADCC-GTL assay format as used here. 18 Comparisons of this measure for uninfected and SP subjects were not significantly different (not shown). The %GrB+ cells generated in the ADCC-GTL assay by cells from TPs were higher than those generated by cells from uninfected subjects (not shown). Together, these results indicate that for the population studied here, including the TPs, HIV infection did not have a negative impact on the %GrB+ CEM target cells generated in the ADCC-GTL assay.
In uninfected subjects, Fc-dependent activation by rgp120-coated CEM cells stimulated a higher frequency of functional 3DL1+ than 3DL1− NK cells. This pattern was not seen for NK cells from ECs. Although we did not perform any NK cell activation assessments on NK cells from TPs, Parsons et al. showed that licensing relevant activation of NK cell by HLA-null cell stimulation was abrogated in NK cells from chronically infected subjects. 14 We did, however, observe that %GrB activity correlated negatively with VL in TPs, suggesting that viremia has a negative impact on delivery of GrB to target cells. Overall, these results indicate that even in subjects with well-controlled viremia, HIV infection has a negative impact on the education relevant responsiveness of NK cells to stimulation by Ab-dependent activation.
Comparison of the %GrB+ cells generated in the ADCC-GTL assay within SPs belonging to the *h/*y+B*57, Bw4, and Bw6 groups and TPs belonging to the Bw4 and Bw6 groups revealed no significant differences. The lower frequency of CD16+ cells within the 3DL1+ NK population that can be educated through engagement of this receptor compared with 3DL1− cells may contribute to the lack of impact of education through 3DL1-Bw4 pairs on the readout of the ADCC-GTL assay. A similar finding was observed when PBMCs from uninfected individuals were used. 18 The low frequency of CD16 + 3DL1+ NK cells would make 3DL1+ NK cells poorer ADCC effectors than their 3DL1− counterparts. Supporting this, there was no correlation between the frequency of CD16+ NK cells and %GrB+ cells for 3DL1+ NK cells, whereas the frequency of CD16 + 3DL1− and %GrB+ CEM cells was positively correlated in uninfected subjects. 18
One interpretation of these results is that 3DL1− NK cells and/or other cell types are mediating effector function in the ADCC-GTL assay. 9 Monocytes and NK cells have been reported to mediate similar levels of ADCC at equivalent effector:target ratios against HIV-infected cells, while neutrophils are not as potent ADCC effector cells. 9 Fc-dependent activation of monocytes and neutrophils for ADCC activity would be independent of KIR/HLA genotype. Furthermore, 3DL1− NK cells activated to mediate ADCC would do so in a manner independent of NK cell education through 3DL1, which could explain the absence of an effect of 3DL1-Bw4 genotype on the %GrB+ cells generated in the ADCC-GTL assay.
To ensure that NK cells were indeed the effector cells in these ADCC-GTL assays, we performed paired tests using PBMCs and PBMCs from which NK cells were depleted by magnetic beads. NK depleted, compared with undepleted PBMCs, generated significantly lower %GrB+ target cell levels (p = .03, Wilcoxon matched pairs test, Supplementary Fig. S5A). On the other hand, %GrB+ cells generated by PBMCs, compared with NK cells isolated from PBMCs, were either equivalent or lower (Supplementary Fig. S5B). Thus, it is unlikely that either neutrophils or monocytes are acting as effector cells in the ADCC-GTL assay. Together, these results suggest that the effector population in the ADCC-GTL assay includes CD16+ NK cells that are mostly 3DL1−.
It should be noted that both 3DL1+ and 3DL1− NK cell populations likely also include NK cells educated through other iNKR/HLA combinations, such as NKG2A and KIR2DL1/L2/L3; 3DL1− NK population may also include hyporesponsive NK cells. 51 However, the impact of education through these other receptor–ligand combinations would be expected to be similarly distributed among 3DL1+/− NK cells. Previous reports show that a higher frequency of educated 3DL1+ than uneducated 3DL1− and educated KIR2DL1+ than uneducated KIR2DL1 NK cells from HIV-uninfected subjects responded to Ab-dependent stimulation by gp120-coated CEM cells. 18,52,53
To our knowledge, this report is the first that focuses on Ab-dependent activation of effector cells from HIV+ subjects and the delivery of GrB by NK cells from HIV+ subjects to CEM targets. It should be noted, however, the stimulation of NK cells from HIV-infected individuals by HLA-null cells has been investigated. Parsons et al. showed that chronic untreated HIV infection abolished the functional advantage of educated 3DL1+ compared with 3DL1− NK cells. Korner et al. showed that educated KIR2DL1/L2/L3+ NK cells from HIV+ subjects responded better than their uneducated counterparts to HLA-null stimulation and that this functional advantage waned from acute infection to early chronic infection time points. 54 This was the case for functions such as IFN-γ and TNF-α secretion, but not for expression of CD107a. 54
In summary, the results presented here indicate that the 3DL1-Bw4 genotype does not influence the ADCC potency of NK cells from SP HIV+ individuals. This may be due to an HIV infection-related loss in the ability of educated 3DL1+ NK cells from Bw4 carriers to respond to Ab-dependent activation and the reduction of CD16-expressing NK cells among 3DL1-coexpressing NK cells.
Interest in ADCC activity and other Ab-dependent functions has increased as evidence implicating these activities in protection from HIV infection has accumulated. 1,2,11,12 The availability of new monoclonal Abs with broad neutralization specificity that also mediate Ab-dependent functions has renewed interest in their use to treat HIV infection. 41,55 Passive transfer experiments using such Abs in humanized mouse models and rhesus macaques are generating a body of evidence that they can protect against challenge with SIV, simian/human immunodeficiency virus, and HIV and/or suppress viral rebound. 56 –60 The finding that KIR-HLA genotypes have a minimal impact on inter-individual differences in ADCC potency among HIV-infected individuals has relevance for the design of therapeutic interventions that may target ADCC for HIV control.
Footnotes
Acknowledgments
The authors wish to acknowledge Drs, Ken Logue, Sylvie Trottier, Benoit Trottier, Sylvie Vézina, Danielle Legault, Jean-Guy Baril, Bernard Lessard, Danielle Rouleau, Claude Fortin, Norbert Gilmore, Bertrand Lebouché, Marina Klein, Edward Ralph, Anita Rachlis, Rupert Kaul, Mona Loutfy, Brian Conway, and Alexandra de Pokomandy for recruiting and following study participants. The authors thank Ms. Stephanie Matte and Mr. Mario Legault for contacting study participants and coordinating their recruitment to this study. The authors are grateful to Ms. Pascale Arlotto and Josée Girouard for expert nursing skills in obtaining leukophoresis samples from participants and to Ms. Tsoarello Mabanga and Ms. Xiaoyan Ni for expert technical support. They also acknowledge the contribution of the study participants.
This study received support from the Canadian Institutes for Health Research (CIHR) MOP-111155 and THA-118628 and the Fonds de Recherche du Québec-Santé (FRQ-S) AIDS and Infectious Diseases Network. G.I. was supported by a postdoctoral scholarship from Canadian HIV Trials Network and CIHR, I.L. was supported by a PhD scholarship from FRQ-S and CIHR. A.T.M. was supported by a Canadian Association for HIV Research Abbvie Master's Award in Basic Science, and N.F.B. is a member of the RI-MUHC, an institution funded, in part, by the FRQ-S.
Author Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.