
Abstract
Antibodies to the carboxy-terminal constant (C5) region 5 of the HIV-1 envelope glycoprotein gp120 have previously been associated with slow disease progression. This is one of the regions on gp120 that interact with the transmembrane glycoprotein, gp41, anchoring it to the viral and infected cell membrane. This study analyzed humoral responses to a novel heterodimeric peptide construct comprising the C5501–512 region and a compatible region on gp41732–744. Antibody levels to C5501–512/gp41732–744 were associated with slow disease progression in a treatment naive historical longitudinal cohort from Norway (n = 32; p = .00001). Elevated anti-C5501–512/gp41732–744 antibody levels correlated with moderate viral load (VL) (50–10,000 copies/ml) in a cohort, including natural viral suppressors (NVS) in the Unites States (n = 58; p = .002). Analysis of HIV-positive sera from treatment naive patients in Estonia (n = 300) showed an inverse correlation between anti-C5501–512/gp41732–744 antibodies and VL when comparing VL 2,000–10,000 copies/ml with VL >10,000 (p = .050). Further mapping using peptide inhibition of antibody binding revealed that responses to the C5501–506 subdomain correlated with preserved CD4 counts (n = 55; p = .0012) irrespective of VL in this cohort. The C5 region encompassing C5501–506 shows sequence similarity to the shared epitope (SE) of certain HLA-DR associated with immune dysfunction. Partial antigenic cross-reactivity between SE and C5 is indicated by partial inhibition of NVS antibody binding using SE 15-mer peptide (median 65% inhibition), the C5501–506 6-mer peptide (79% inhibition), and binding of rheumatoid arthritis patient sera to both SE and C5 peptide sequences. The potential influence of these observations on HIV-1 pathogenesis remains to be determined.
Introduction
I
LTNP have been shown to have high titer antibodies to the fifth constant (C5) domain on the HIV surface glycoprotein gp120, 5 –7 whereas rapid progressors had high titers to a portion of the V3 loop. 5 Furthermore, loss of high titer antibodies to C5 immediately preceded progression to AIDS. 8 The inability of anti-C5 antibodies to neutralize HIV infection in standard biological neutralizing assays may explain why the C5 region has not been considered for vaccine development. 9
Previous studies that correlated the presence of antibodies to C5 with slow disease progression used a linear peptide antigen corresponding to the highly conserved carboxy-terminal 15 amino acids from the C5 region for antibody detection. In virions, however, the C5 region interacts with the transmembrane glycoprotein, gp41, to anchor gp120 to the membrane. 10,11
A compatible region on gp41 was identified leading to the design of a heterodimeric peptide construct incorporating the highly conserved C5 domain and this compatible region on gp41 (Vacc-C5). This peptide antigen was used to verify previous observations linking the presence of antibodies to C5 with slow disease progression and associated lower pan-immune activation using a historical longitudinal cohort from before cART was introduced (n = 32) and a second cohort of treatment naive natural viral suppressors (NVS) (n = 58). Analysis of a larger cohort of treatment naive HIV-positive sera (n = 300) revealed a correlation between antibody responses to a subdomain on C5 with preserved CD4 counts irrespective of VL.
Materials and Methods
Sequence analysis and peptide synthesis
Curated alignments of the Env gene (year 2010, M-group excluding recombinants) were downloaded from the HIV sequence database (
The two peptides contained within C5501–512/gp41732–744 are linked by a thioether bond between the side chains of the marked cysteine and lysine residues.
The bold underline indicates where the C5 and gp41 peptides are linked.
Serology cohorts
Sera from a cohort of HIV-1-infected individuals from the United States (n = 58), including NVS, were kindly provided by the Institute of Human Virology, University of Maryland, United States. Two blood donor sera were included in the analysis and the samples tested blind. Archival plasma samples from the Oslo cohort (time period 1989–1995 before the introduction of cART) were kindly provided by the Norwegian Institute of Public Health. From this large cohort, slow progressors were selected as subjects that survived for >10 years following diagnosis (n = 16). Fast progressors were selected as those subjects that died within 5–6 years of diagnosis (n = 16). Of the subjects included, the majority of fast progressors died within 2 years. Since these were archival samples, VL was retested using COBAS AmpliPrep/COBAS TaqMan HIV Test v2.0 (Roche) using a plasma dilution of 1/10 corresponding to a detection level of 400 copies/ml. Any degradation of viral RNA over time in these samples was considered to be similar between the two groups.
Serum samples from treatment naive HIV-positive subjects (n = 300) were kindly made available for testing on site at the HIV reference laboratory at West-Tallinn Central Hospital, Estonia. Sera were collected according to the following VL ranges: VL <10,000 copies RNA/ml (n = 100); VL 10,000–50,000 copies RNA/ml (n = 99); and VL >50,000 copies RNA/ml (n = 101). Information on VL, CD4, and CD8 counts was available for these patient sera.
Serum samples (n = 33) from a panel of HIV-negative sera with rheumatoid arthritis were obtained from Bionor Laboratories, Skien, Norway.
Blood donor sera were obtained from Telemark Central Hospital, Norway. Sera from chronically infected subjects were from Oslo University Hospital, Norway, and LTNP sera were provided by Dr. Dalgleish at St. George's, University of London, United Kingdom.
All human sera were analyzed anonymously. Serum collection was carried out according to the regulations at each institution, and permission to use the serum samples was obtained from the respective principal investigators. Voluntary blood donation includes consent to the potential use of blood for research purposes.
Rabbit immunizations to generate anti-C5501–512/gp41732–744 (Vacc-C5) antibodies
New Zealand White Rabbits (Oryctolagus cuniculus) were immunized subcutaneously at weeks 0, 2, 6, and 10 with 500 μg Vacc-C5 using Freund's adjuvant (50% v/v) in a volume of 1 ml/dose/rabbit. Blood was collected 2 weeks after each immunization. Serum was extracted from the blood using conventional techniques and stored frozen at −70°C until used. Immunization of rabbits was approved by the Norwegian Animal Research Authority (study number FOTS 442) and conducted according to national guidelines, which complied with the European Commission's animal research directive (86/609/EEC) and the Council of Europe ETS 123: European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes.
Serological analyses
Antibody responses to C5, gp41, and V3 loop antigens were determined using an ELISA. Blood donor sera were used as negative controls. Briefly, MaxiSorp 96-well ELISA plates were coated with Vacc-C5 or the C5 and gp41 peptide components separately as required, at a concentration of 1.6 μg/well in a volume of 100 μl, and incubated at 4°C for 4 days. After extensive washing, the plates were stored at 4°C until used. Five-fold dilutions of sera were added and incubated 2 h at 37°C. HIVLai V3 biotinylated peptides (0.25 μg/well in 100 μl) were coated on streptavidin strips (Thermo Scientific) for one hour at room temperature (RT) before use. Start dilution for detecting anti-V3 antibodies was 100-fold with incubation at RT.
After six washing steps a protein G conjugated Alkaline Phosphatase was added (0.3 μg/well) and incubated for 1 h at room temperature. The substrate (0.3% w/v Phenolphthalein Monophosphate, pH 8.2) was added for 15 min after which stopping solution was used that mediates a color change at 550/620 nm in proportion to the amount of bound antibody. This was converted to μg/ml by the use of a positive control sheep anti-C5 serum from Aalto, Dublin, Ireland (D7324). The positive control polyclonal antiserum to HIV-1 LAI V3 loop (Catalogue No. 0433) was purchased from NIBSC, United Kingdom.
For antibody blocking assays, rabbit or human sera at a titrated dilution were preincubated with excess peptide (100–500 μM) before plating and the percentage inhibition determined. For cross competition of antibody binding, the ELISA was carried out as described except that dilutions of pooled human sera (1:2–1:50) were mixed with rabbit or human serum (fixed 1:10 dilution) before plating, and the percentage inhibition of antibody binding was determined using Alkaline phosphatase–conjugated goat anti-rabbit IgG (6 μg/ml) (D-0487; DAKO) or rabbit anti-human IgG (D-0336; DAKO).
Thirty-three sera from a panel of HIV-negative sera with rheumatoid arthritis (Bionor Laboratories, Skien, Norway) were tested in peptide inhibition experiments measuring optical density (OD) using streptavidin plates with bound K(Biotin)-EQKRAA (corresponding to shared epitope [SE]) and sera optionally blocked with EQKRAA. The same sera were tested for binding to biotinylated C5501–506 and to streptavidin plates without peptide. Two sera were excluded because the OD for the blank plates exceeded cutoff for background, set to the medium +3 Sigma of OD from dilution buffer alone.
Results
Design of the Vacc-C5 peptide antigen
The C5 domain corresponds to the 41 carboxy-terminal amino acids following the fifth variable domain (V5) of gp120 where the last 15 amino acids are highly conserved. For the reference HIV-1 sequence, HXB2 (clade B), this sequence corresponds to: 497-APTKAKRRVVQREKR-511.
A region encompassing a stretch of highly conserved negatively charged residues on gp41, compatible with the conserved positively charged residues on C5, was identified (corresponding to aa's 732–744 on the reference strain HXB2). A complex peptide antigen incorporating the C5 sequences chemically linked to gp41732–744 and representative for the predominant clades A, B, C, and D was designed (Vacc-C5) (Table 1 and Fig. 1). This peptide antigen induced antibodies in rabbits that cross competed with HIV-positive human sera from LTNP and NVS showing that sera generated following natural infection recognized the C5501–512/gp41732–744 (Vacc-C5) peptide antigen (Table 2).
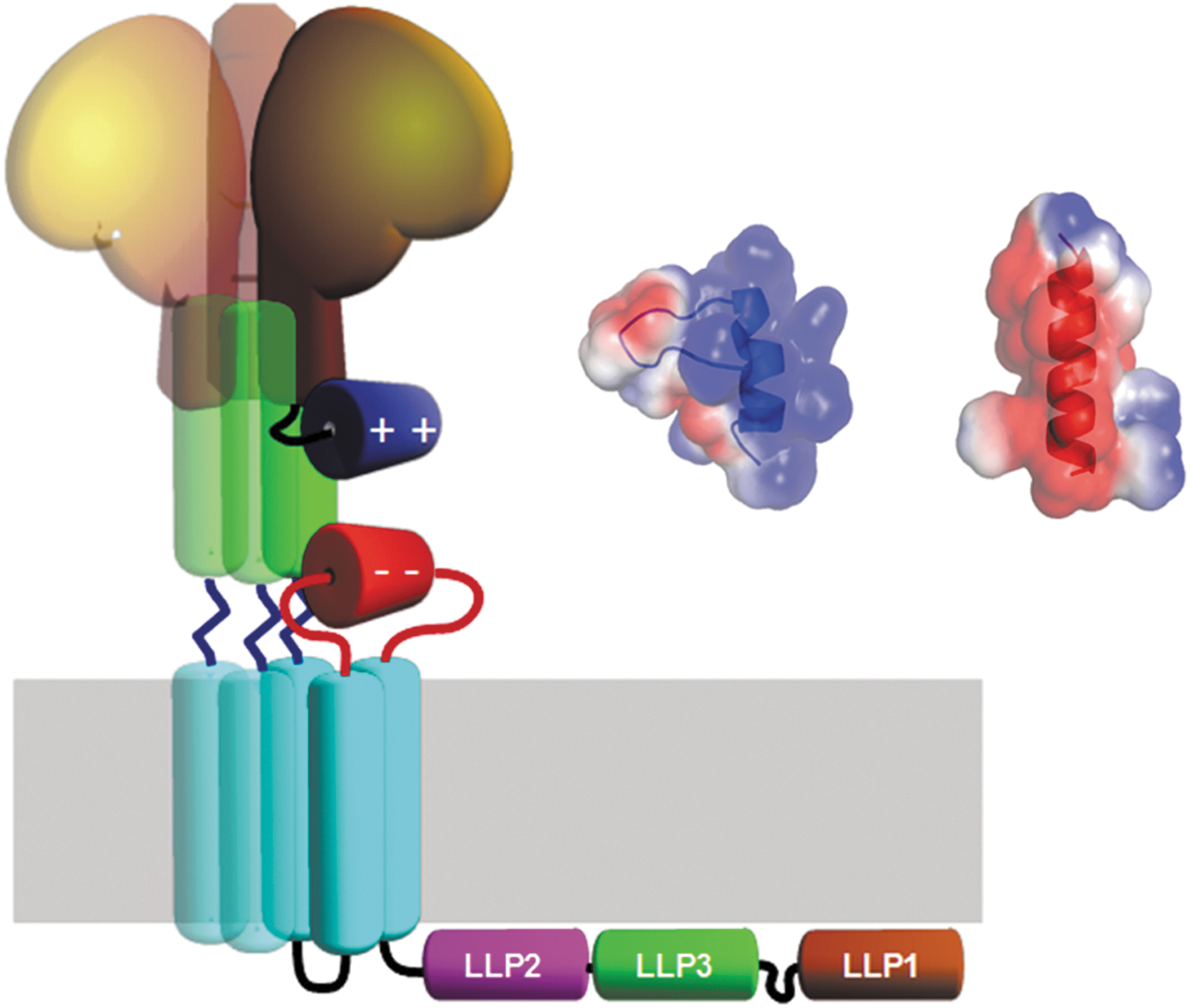
Illustration of gp120 (brown) and the interaction between the C5 and gp41 domains. The envelope glycoprotein gp120 (brown) with its C-terminal C5 region (blue) and its potential interaction with the gp41 C-helix (green) and the extracellular loop of gp41 (red). The trimeric nature of the complex is illustrated by shaded copies of the central parts. Inset shows electrostatic surface representation of the NMR structure of the C5-region (blue helix) and modeled helical structure of gp41 fragment 728–741 (red helix). NMR, nuclear magnetic resonance.
Peptides and human sera were used to inhibit rabbit anti-C5501–512/gp41732–744 antisera binding to C5497–511 and gp41728–745 peptide sequences. The data shown are for peptide inhibition at 500 μM concentration. LTNP serum pool from Ullevål University Hospital, Norway. NVS sera from University of Maryland and blood donor sera were provided by Telemark Central Hospital, Norway.
LTNP, long-term nonprogressors; NVS, natural viral suppressors; PBS, phosphate buffered saline.
Antibody responses to Vacc-C5 peptide and disease progression
Antibody levels to Vacc-C5 were investigated in relation to disease progression using a cohort of archival sera from subjects at the Norwegian Institute of Public Health, from the time period 1989–1995, before the advent of combination ART. 13 Slow progressors (n = 16) that lived for more than 6 years without ART had significantly higher levels of IgG (μg/ml) to Vacc-C5 than fast progressors, where the majority died within 2 years (Fig. 2). The level of anti-Vacc-C5 antibodies was found to be stable over time for each individual; however, the time period for the progressors was shorter (2 years compared to 4–5 years, data not shown).

Serological responses of a treatment naive historical longitudinal cohort. Shows data from a longitudinal cohort of 32 patients (16 in each group) with fast or slow progression of HIV-1 infection. Median anti-C5501–512/gp41732–744 (Vacc-C5) IgG concentration based on earliest available sample was 0.38 and 6.61 μg/ml, for fast and slow progressors, respectively. Similarly, median anti-V3 loop IgG concentrations were 1,569 and 2,591 μg/ml for fast and slow progressors, respectively. Box shows median, interquartile range, whiskers are 1.5 times interquartile range, and diamonds are measurements outside this range. p-Values are from a Student's t-test on log10-transformed concentrations.
The 16 patients in the fast progressors group had a median anti-Vacc-C5 IgG of 0.38 μg/ml (from the first available visit), whereas the 16 slow progressors had a median of 6.61 μg/ml. A level of 2 μg/ml was found to differentiate these two groups. Of the 16 fast progressors, only 2 out of 16 mounted an anti-Vacc-C5 IgG response above 2 μg/ml plasma in contrast to the slow progressors, of which 14 of 16 patients had an anti-Vacc-C5 IgG level above 2 μg/ml plasma.
VL levels were retested after anti-Vacc-C5 antibody status had been determined and remained relatively constant between the first and last time point tested. Median VL of the slow progressors for the first sample was 400 copies HIV-1 RNA/ml (detection limit) (n = 16) and 440 copies HIV-1 RNA/ml for the last sample (n = 15), whereas for the fast progressors median VL was 5,300 copies HIV-1 RNA/ml for the first sample (n = 16) and 13,000 copies HIV-1 RNA/ml for the last sample (n = 16). VL was significantly different between the two groups for the first plasma sample taken (p = .001, Student's t-test on log10-transformed values). The time span between the first and last samples in the slow progressors was 4–5 years, whereas for the fast progressors this was 2 years.
As a control, the level of antibody to the HIVLAI V3 loop (Table 1) was also determined since this was the strain used in earlier studies. 5 The 16 patients in the fast progressors group had a median anti-V3 loop antibody level of 1,569 μg/ml, whereas slow progressors had a level of 2,591 μg/ml (p = .21) (Fig. 2). This corresponded to an ∼2-fold difference in median antibody level between the groups compared to a 17-fold difference for median anti-Vacc-C5 antibody levels (p = .00001, Student's t-test on log10-transformed values).
These findings support a link between the level of anti-Vacc-C5 antibodies and VL/disease progression.
Antibody responses to Vacc-C5 inversely correlated with VL in sera from a cohort that included NVS
Sera from a U.S. cohort of HIV-infected patients that included NVS, that is, patients who have undetectable HIV VL without cART 14 were also tested for the presence of anti-Vacc-C5 antibodies. Patients with intermediate VLs (50–10,000 copies/ml) had high levels of anti-Vacc-C5 antibodies (Fig. 3), whereas subjects with high VL (>10,000 copies/ml) had low anti-Vacc-C5 antibody levels suggesting that a loss or reduction was associated with progression. NVS who had no detectable virus (<50 copies/ml) also had low anti-Vacc-C5 levels. This suggests that a low-viral antigen load provided negligible antibody stimulation. A statistically significant inverse correlation between anti-Vacc-C5 antibodies (IgG) and VL was observed (n = 58; p = .002).
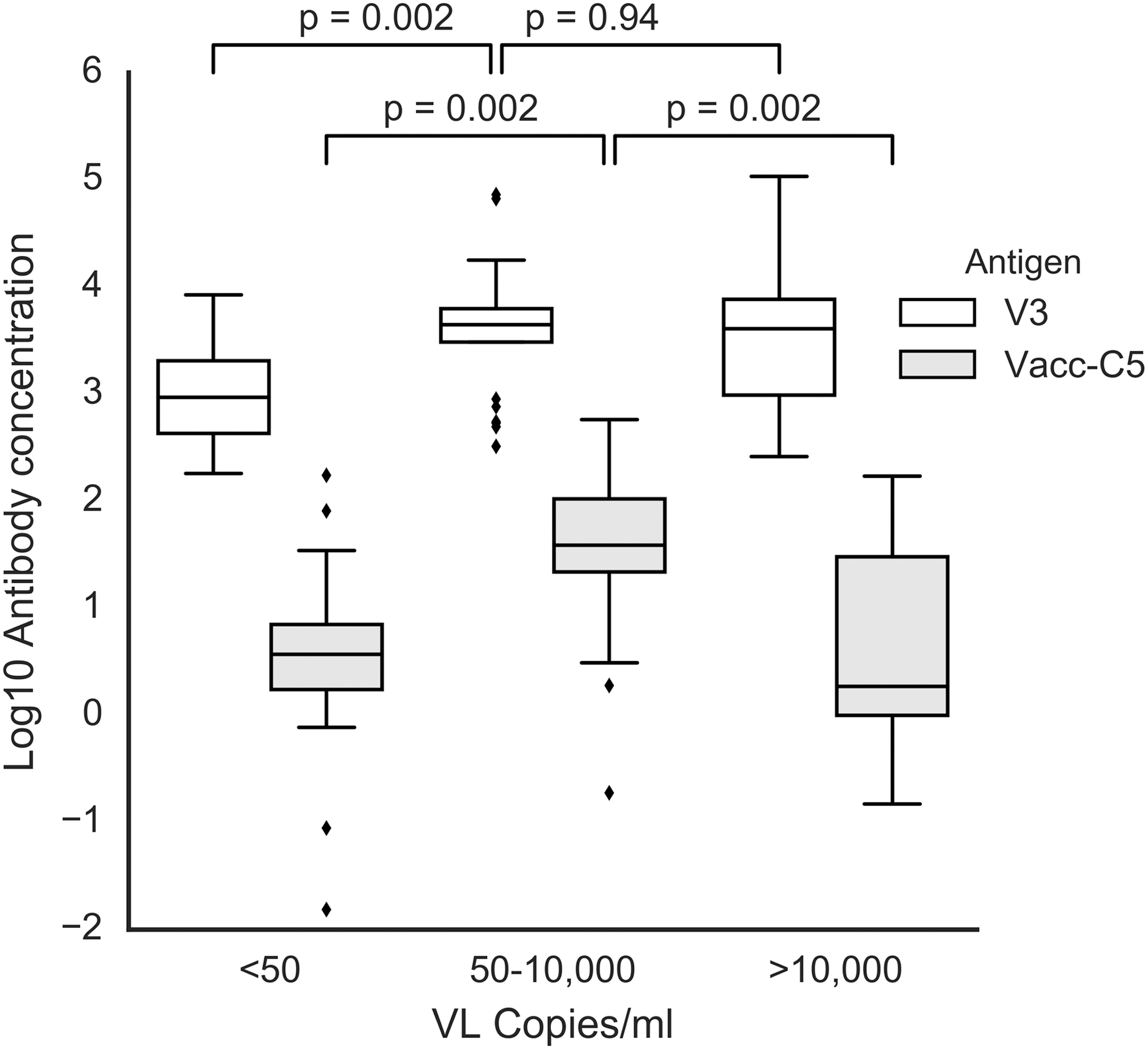
Serological responses in a cohort, including NVS. HIV-1-infected individuals (n = 58) from a cohort, including HIV-1 natural virus suppressors, were tested for anti-C5501–512/gp41732–744 (Vacc-C5) IgG concentration and correlated with individual viral load. The subjects were divided into three groups, subjects with <50 copies HIV-1 RNA/ml (n = 16), subjects with 50–10,000 copies HIV-1 RNA/ml (n = 25), and subjects with >10,000 copies HIV-1 RNA/ml (n = 17). Median anti-Vacc-C5 antibody levels were 3.7, 38.9, and 1.9 μg/ml, respectively. Median anti-V3-loop antibody levels were 943, 4,418, and 4,044 μg/ml, respectively. p-Values are calculated from a Mann–Whitney U test on antibody concentrations. Box shows median, interquartile range, whiskers are 1.5 times interquartile range, and diamonds are measurements outside this range. NVS, natural viral suppressors.
Levels of anti-V3 loop antibodies were orders of magnitude higher than anti-Vacc-C5 antibody levels regardless of VL. Furthermore, in contrast to anti-Vacc-C5 antibody responses where there was a significant difference in antibody levels in relation to highest VL (VL 50–10,000 and VL >10,000; p = .002), antibodies to the V3 loop did not show the same significant difference (VL 50–10,000 and VL >10,000; p = .94) (Fig. 3).
Antibody responses to a subdomain on C5 correlated with preservation of CD4 T cells
Treatment naive HIV-positive sera (n = 300) were tested for seroreactivity to Vacc-C5 with respect to VL. No statistically significant correlations were observed when the data were analyzed using different VL stratifications, although the difference between anti-Vacc-C5 antibody levels in patients with VLs between 2,000 and 10,000 copies RNA/ml (n = 69) compared to those in patients with VLs >10,000 copies RNA/ml (n = 200) approached significance (p = .050; Mann–Whitney) (Table 3).
Antibody responses to Vacc-C5 in treatment naive HIV-positive individuals (n = 300) were correlated with plasma viral load. A statistically significant correlation (bold) was only observed when comparing sera from individuals with plasma viral loads of 2,000–10,000 copies and >10,000 copies/ml (p = .05, Mann–Whitney test). “—” Indicates correlations not possible.
Peptides corresponding to subparts of the Vacc-C5 construct as shown in Table 1 were used to block binding of antisera to the Vacc-C5 antigen. Only sera with sufficient anti-Vacc-C5 antibody (>5 μg/ml) (n = 55) could be used for this analysis. The results (Table 4) showed a mean inhibition of 52.1% inhibition of serum binding for peptide C5501–506, whereas peptide C5504–510 showed only a mean of 18% inhibition. The linear C5501–512 and gp41732–744 showed mean inhibition of 82.8% and 67.6%, respectively. The longer peptides (C5501–512) may be more effective because they may help to maintain the helical conformation of the C5501–506 region.
A Pearson's correlation analysis (on log10-transformed data) between peptide inhibition of anti-Vacc-C5 binding and VL, CD4, and CD8 counts, respectively, is shown in Table 5. Peptide inhibition using C5501–506 showed a significant correlation with CD4 count (p = .0012) (Table 5 and Fig. 4).

Correlation between inhibition of anti-C5501–512/gp41732–744 (Vacc-C5) antibody binding by peptide C5501–506 and CD4 T cell counts in treatment naive HIV-infected individuals (n = 55) with anti-Vacc-C5 antibody levels >5 μg/ml.
Statistical significance p > 0.05.
The C5 domain has been shown to have sequence similarity to the third hypervariable region (HVR3) of the HLA-DR β chain. 15,16 The HVR3 is an allo-epitope, associated with antigen/peptide interactions, T cell regulation, and allo-recognition. These HLA-DR are characterized by the “shared epitope,” which comprises five amino acids in the HLA-DR sequence; QKRAA; QRRAA; and RRRAA (Table 6) are implicated in the susceptibility and/or severity of rheumatoid arthritis (RA). 17 These sequences show similarity to predominant C5 sequences (QRRVV, KRRVV, RRRVV), which correspond to the region covered by peptide C5501–506. Therefore the recognition of SE containing peptides was assessed.
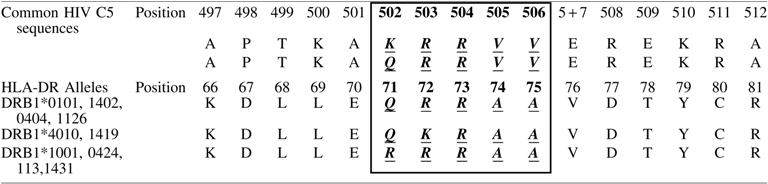
Shared epitope sequences are shown within the boxed section as bold, italic, and underlined.
The SE sequences were synthesized as a pool of 5 and 6-mers (DERAA, EDERAA, GQKRAA, EQKRAA) and a 15-mer sequence (KDLLEQKRAAVDTYC) to similarly block sera from the NVS cohort since no further serum was available from the Estonian cohort. Only the 15-mer SE sequence showed blocking (median 65%, n = 18) in the range of that shown by C5501–506 (median 79%, n = 18) (Fig. 5). Blocking using sheep control antisera (raised to a linear C597–511 peptide), a similar inhibition, was observed as for SE 15-mers as C5501–506 41% (n = 1).
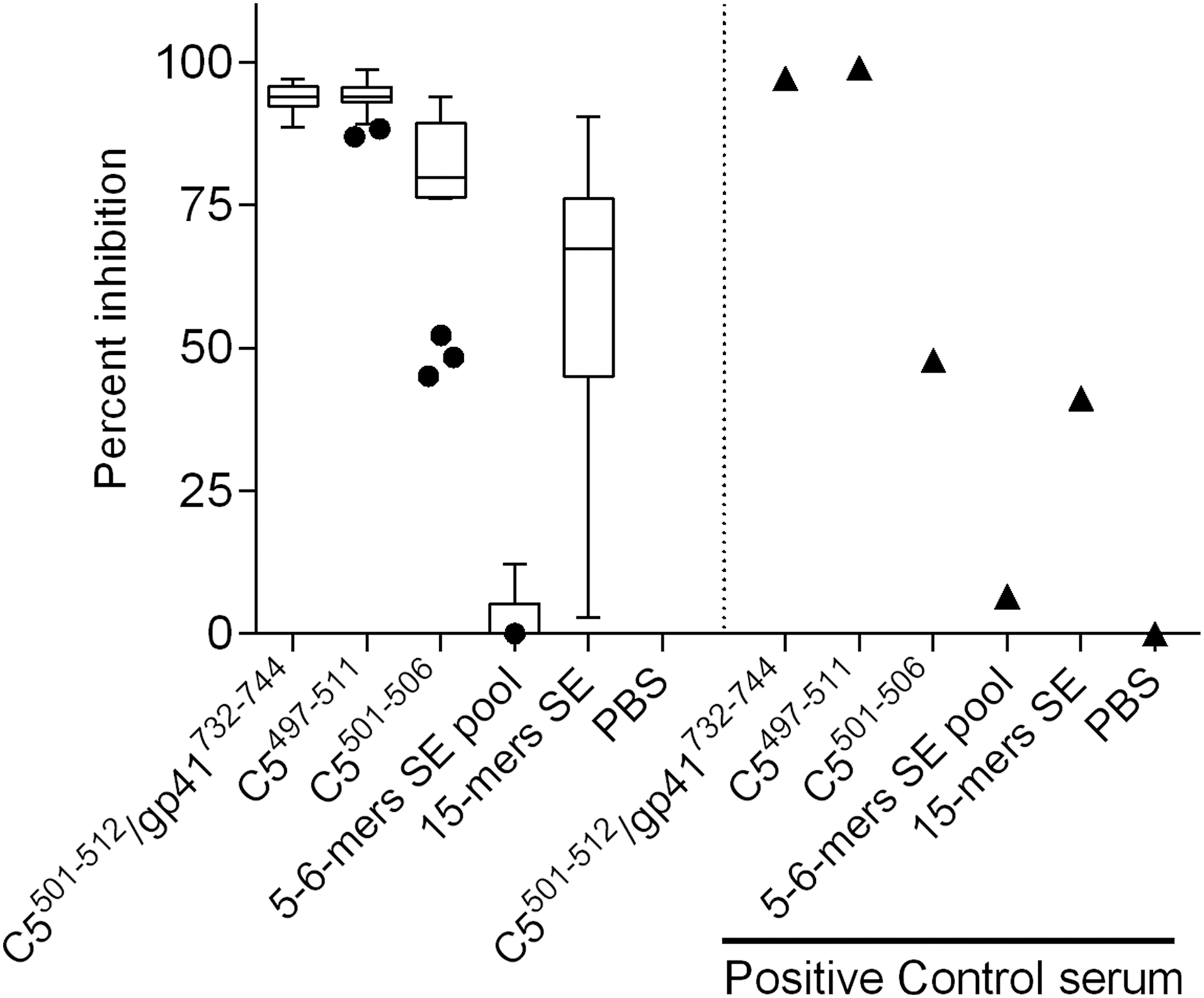
Peptide inhibition of antibody binding in subjects from the cohort, including NVS. Only sera that had anti-C5501–512/gp41732–744 levels >5 μg/ml (n = 18) using peptides corresponding to C5 subdomains, gp41 and SE. (1) C5501–512/gp41732–744; (2) C5497–511; (3) C5501–506; (4) 5–6-mer SE pool; (5) 15-mer SE. Positive control: sheep anti-C5497–511 antisera (n = 1). Box shows median, interquartile range, whiskers are 1.5 times interquartile range, and dots are outliers. The percent inhibition is relative to a negative control of no peptide (phosphate-buffered saline alone). SE, shared epitope.
Furthermore, when testing sera from HIV-negative individuals with rheumatoid arthritis, measurements of OD of binding to C5501–506 above background (blank streptavidin plates) showed significant Linear Regression (Adj. r 2 = 0.253, p = .002) on OD measurements of specific binding to the SE containing peptide EQKRAA (excess OD over sera blocked with peptide). There was insufficient serum available to test by blocking with scrambled peptides or other antibodies, for example, anti-gp120.
Discussion
In this study, previous observations of an association between detectable antibody responses to C5 and slowed disease progression were confirmed and extended using an antigen that incorporated not only C5 sequences but also a compatible region on gp41. The region on gp41 included in Vacc-C5 has not previously been implicated in C5 interactions.
According to traditional models of gp41, this region, which includes the Kennedy epitope 18 and the highly immunogenic region, lies beneath the membrane and would not be available for interaction with C5. 19,20 However, an alternate model for gp41 topology was proposed by Hollier and Dimmock in 2005, 21 which suggests that the cytoplasmic tail of gp41 traverses the membrane more than once thereby exposing the Kennedy epitope. Exposure of the C5501–512/gp41732–744 region would make it available for antibody binding to the surface of infected cells.
In 2012, Postler et al. provided evidence indicating that surface exposure of the Kennedy epitope was an artifact from nonspecific binding of HIV envelope glycoproteins at the cell surface. 22 Although the same authors in a later review stated that the existence of an alternate structure could not be excluded at this time. 23 Steckbeck et al. 19 addressed this potential artifact using cells transfected with gp41 and nontransfected cells as a control, in the presence of clarified supernatants from Env expressing cells. Antibodies to the Kennedy epitope only bound the gp41 transfected cells suggesting that under such conditions, the detection of the Kennedy epitope by antibodies to gp41 was not due to nonspecific binding of Env from other cells. More recently, Kyrychenko et al. undertook alternate computation and experimental approaches and proposed a different model to that of Hollier and Dimmock, but which nevertheless suggests that the gp41 cytoplasmic tail can traverse the membrane more than once thereby exposing the Kennedy epitope. 24 To date, the structure of the cytoplasmic tail of gp41 remains unresolved and controversial; however, the potential existence of multiple gp41 topologies at the cell and virion surface represents the current thinking.
Rapid endocytosis of HIV-1 envelope glycoproteins, probably to avoid antibody-dependent cell-mediated cytotoxicity (ADCC), 23 may explain why few envelope glycoproteins become incorporated in to virions. 21
Detectable anti-Vacc-C5 antibody responses correlated with moderate VL in the cohort, including NVS. In the historical longitudinal cohort, the observation that some fast progressors (2/16) produce >2 μg/ml anti-Vacc-C5 antibodies and some slow progressors (also 2/16) had <2 μg/ml anti-Vacc-C5 antibodies is consistent with the existence of other correlates of HIV-1 pathogenesis or protection, such as microbial translocation, type I interferon, or host expression HLA-B57, which demonstrate similar correlations and whose influences are not fully understood. 25 –29 This is indicative of a complex disease etiology involving a number of related factors which determine either control or progression of disease. 30 This is also emphasized by the observation that NVS patients with low VL had little production of anti-Vacc-C5 antibodies possibly due to other protective mechanisms such as the CCR5 delta-32 mutation resulting in low levels of circulating antigen in these individuals. Whether anti-Vacc-C5 antibodies exert a protective influence on HIV-1 disease is yet to be determined.
In the larger cohort from Estonia, the correlation between antibody responses to Vacc-C5 and VL was only statistically significant for the subpopulation of samples with VL (2,000–10,000) and VL (>10,000) (Table 3). The reason why anti-Vacc-C5 antibodies are lost on disease progression with a consequent increase in VL is not fully understood.
Preservation of antibody responses to p245, 31 and Tat 32 has also been shown to be associated with slowed disease progression, although the mechanism(s) involved remain to be determined. Loss of antibodies to HIV antigens such as p24, Tat, and antibodies to the V3 loop may, in part, be due to loss of CD4 help to B cells following CD4 T cell depletion upon disease progression. However, we observed that the level of anti-Vacc-C5 antibodies was much lower, in fast progressors, and the fold reduction in anti-Vacc-C5 antibodies between progressors and nonprogressors was much higher compared to that of anti-V3 loop antibodies. Furthermore, in contrast to antibodies to Vacc-C5, the level of anti-V3 loop antibodies did not show the same correlation with VL suggesting that the mechanism(s) associated with lower anti-Vacc-C5 antibody and reduction in anti-V3 loop antibody with increased VL (disease progression) may have differences.
Antibodies against Vacc-C5 are non-neutralizing. The potential role of such antibodies in HIV infection is gaining interest 33 particularly following the RV144 clinical study that showed a marginal statistical significance of protection, likely a consequence of non-neutralizing antibodies. 34 Antibodies to the C5 region of HIV-1 have been shown to participate in ADCC, 35 which could represent one mechanism by which these antibodies exert an effect on VL and disease progression.
The envelope glycoprotein gp120, including C5, 36 and the transmembrane glycoprotein gp41 37 show strong homology to HLA (particularly HLA-DR), which includes the “shared epitope (SE),” a five amino acid sequence within HLA-DR implicated in autoimmune disease such as rheumatoid arthritis (RA) 38 (Table 6). Although there was also no correlation between antibody responses to the complete heterodimeric C5501–512/gp41732–744 complex and CD4 counts, an interesting correlation was observed between antibody responses to a subpart of C5 (C5501–506) and CD4 counts (Fig. 5 and Table 5) that has not been described previously. This short peptide corresponded to the SE region of HLA-DR (Table 6).
A 15-mer peptide to SE showed inhibition of anti-C5501–512/gp41732–744 antibody binding (median 65% inhibition), whereas the shorter 5–6-mer peptide pool did not. Both SE and C5 have helical regions, which may not have been maintained as conformational epitopes in the shorter peptides. The observed inhibition of binding by the SE 15-mer and C5501–506 was greater for the cohort, including NVS (n = 18), which are generated against HIV natural infection and may include antibodies to C5 alone (from soluble gp120), gp41 alone (after gp120 shedding), or the C5/gp41 complex on the surface of infected cells. In contrast, the lower level of inhibition observed for the sheep control serum (n = 1) could be because the antigen used for immunization was a single linear peptide C5497–511. It also suggests that in the absence of a helical structure, the SE sequences may need to exhibit greater sequence identity to C5501–506 (AKRRVV—see Table 6).
The mechanism by which the SE exerts its influence on rheumatoid arthritis has been extensively studied, but is not fully understood. Several nonmutually exclusive hypotheses have been proposed, including antigen presentation, T cell selection, and innate immune activation. 39 –42 It is unclear whether gp120 could replicate these through C5 homology to the SE. Another possibility is that through this homology to the SE, anti-C5 immune responses could be subject to immune tolerance, which could, in part, explain the low level of anti-Vacc-C5 antibodies observed in fast progressors. Poor immunogenicity and autoreactivity are characteristics of antigens that can induce protective antibody responses to HIV-1. 43,44 It is therefore interesting to note that anti-C5 antibody detected in human colostrum was found to be highly autoreactive, 45 while SE-specific T cell responses are often absent from rheumatoid arthritis patients carrying HLA-DR harboring SE 46 indicating tolerance mechanisms to anti-SE immunity.
Antibodies to C5501–512/gp41732–744 are linked to slow disease progression and the associated low HIV-associated immune activation. In addition, antibodies to C5501–512/gp41732–744 (Vacc-C5) have recently been shown to be associated with improved VL outcome following therapeutic vaccination with the peptide-based candidate, Vacc-4x, and treatment interruption. 47
The potential influence of immunological cross-reactivity between C5501–506 and SE on HIV pathogenesis, immune activation, and disease progression remains to be determined. It is remarkable that HIV-2, which is associated with a milder disease course, lacks the level of similarity in its C5 domain to the conserved 15 carboxy-terminal amino acids of the HIV-1 C5 domain. This may, in part, explain the lower immune activation associated with HIV-2 infection. 48
Footnotes
Acknowledgments
This work acknowledges the contribution and memory of Kjell Skaug, Norwegian Institute of Public Health. The authors thank Jens Holmberg for his contribution to the animal immunogenicity studies, Tønnes Lange for his contribution to the peptide sequence analysis, Dr. Mohammed M. Sajadi for his work on the cohort at the University of Maryland, and Dr. Dag Kvale at the University of Oslo providing test sera. This work was supported by grants from the Research Council of Norway BIA (project no.: 187903) and GLOBVAC (project no.: 228323) programs.
Part of this work was presented as a poster at AIDS Vaccine 2013, Barcelona, Spain. The abstract is published in AIDS Res. Hum. Retroviruses 29 (11):142 Supplement. Abstract P13:19.
Author Disclosure Statement
M.A.S., G.S., and M. Ö. are employees of Bionor Pharma AS, and have shares in the company. B.S. and A.-O.H. were employees of Bionor Pharma AS when the work was carried out. B.S. has shares in the company. J.Z. was a member of the Bionor Pharma AS Board of Directors at the time of the study with shares in the company. Angus Dalgleish is a member of the Bionor Pharma Clinical Advisory Board (CAB) while Robert Redfield was a member of the CAB at the time of this study. The remaining authors G.H., P.L.S., Ø.J., and V.U. have no conflicts of interest.