
Abstract
The impact of HIV infection on the frequency and differentiation capability of CD34+ bone marrow hematopoietic progenitor cells (BM-HPCs) is still debated, having a possible primary role in antiretroviral-induced immunoreconstitution. We investigated the influence of HIV replication or proinflammatory cytokines on lymphopoietic capability of BM-HPCs from seven viremic (VR) and five nonviremic (NVR) HIV-infected patients. We found that BM-HPCs from VR patients were unable to differentiate in vitro toward T cells, and produced proinflammatory cytokines in the absence of viral replication. In contrast, the lymphoid differentiation potential of BM-HPCs was partially restored in successfully antiretroviral therapy-treated patients. We also showed that TLR8 triggering induced BM-HPCs from healthy donors to release proinflammatory cytokines affecting T cell differentiation. These data suggest that in HIV-infected patients, the lymphopoiesis capability of BM-HPCs may be modulated by a virus-driven autocrine mechanism involving proinflammatory cytokines.
Introduction
C
In fact, on one hand it has been reported that HIV disease progression is directly linked to alterations in the HPCs compartment, despite treatment-induced viral suppression 5 ; on the other hand, we previously showed that a very early introduction of cART may preserve lymphopoietic capability concomitantly to a rapid decrease of immune activation markers. 6 However, the effect of immune activation in HIV-infected patients on HPCs functionality remains a matter of debate.
For these reasons we were prompted to investigate the lymphocyte development capability of CD34+ BM-HPCs during HIV infection, and to analyze the influence of HIV replication on this process. With this aim, CD34+ BM-HPCs from viremic (VR) and nonviremic (NVR) chronically HIV-infected patients were induced to differentiate toward T cells in an in vitro culture system following a program that resembles the process normally occurring in the thymus. 7 Moreover, we analyzed the effect of inflammatory cytokines on T cell lineage commitment from CD34+ BM-HPCs during HIV infection.
Materials and Methods
Study population
Twelve chronically HIV-infected patients requiring bone marrow (BM) aspirate for diagnostic purposes (fever of unknown origin, pancytopenia, suspect of lymphoma) were enrolled in the study. The study was approved by the local Institute Ethics Committee (approval no. 4 dated January 22, 2013) and all enrolled subjects signed the informed consent. BM aspirates from 12 chronically HIV-infected patients were found normocellular in 9 and with a mild HIV-related dysplasia in 3 patients.
Ten out of 12 patients were receiving cART at the time of BM aspirate. Patients were classified in NVR (seven cART-treated patients with viral load <40 copies/ml) and VR patients (five patients with viral load ranging from 317 to 855,000 copies/ml, including two untreated and three cART-treated patients). Patients' features are summarized in Table 1. Three HIV-seronegative donor BM cells were used as a control and purchased from Stem Cell Technologies (Vancouver, Canada).
Gray shade identifies patients used to evaluate BM-HPC in vitro differentiation potential.
ART, antiretroviral therapy.
Bone marrow mononuclear cell separation and purification of CD34+ cells
Bone marrow mononuclear cells (BM-MNCs) were isolated from BM aspirate by density gradient centrifugation (Lympholyte-H; Cederlane, Canada). After separation, BM-MNCs were resuspended in phosphate-buffered saline for magnetic labeling. Freshly isolated CD34+ HPCs were purified from BM-MNCs by a two-step magnetic procedure involving lineage depletion, followed by CD34+ isolation (the CD34 Diamond Isolation Kit; Miltenyi Biotec, Calderara di Reno [BO], Italy). The purity of CD34+Lin− cells was always >98% as determined by flow cytometry.
Antibodies
The following panel of mouse anti-human mAbs was used: phyco-erythrin-Cy7 and allophycocyanin-conjugated anti-CD34, PE-conjugated anti-CD38, APC-H7-conjugated anti-CD45RA, V450-conjugated anti-CD7, fluorescein isothiocyanate (FITC)-conjugated anti-CD4, FITC-conjugated anti-Lin (CD3, CD14, CD16, CD19, CD20, and CD56), APC-H7-conjugated anti-CD8, and AmCyan-conjugated anti-CD45 (BD Bioscience, San Jose).
Flow cytometry
The frequency of CD34+Lin− cells and, among these, CD34+Lin−CD38−CD45RA+CD7+ lymphoid HPCs (L-HPCs) progenitors as well as the generation of CD4 and CD8 T cell progeny was determined by flow cytometry on 1 × 105 viable cells. In brief, 1 × 105 cells were incubated with antibodies mix for membrane staining at 4°C for 15 min, then washed and fixed with paraformaldehyde 1% (PFA; Sigma-Aldrich). Sample acquisition and data analysis were performed by a FACS Canto II Flow Cytometer (Becton Dickinson, San Jose, CA) by using Diva software.
Human OP9-DL1 cell coculture
BM-CD34+ HPCs differentiation toward T cells was induced by coculturing with OP9-DL1 stromal cells expressing the Notch 1 ligand Delta-like 1 (kindly provided by Dr. Zuniga-Pflucker) for 3 weeks, in the presence of interleukin (IL)-7 (1 ng/ml; Immunotools, Friesoyte, Germany), Stem Cell Factor (30 ng/ml; ENZO Life Science Farmingdale, NY), and FMS-like tyrosine kinase 3 (FLT3) ligand (5 ng/ml; PeproTech, Inc., Rocky Hill, NJ), as described. 7 Briefly, 5 × 104 BM-CD34+ cells were cultured on OP9-DL1 cells seeded in wells (50%–80% confluence) using a 24-well plate in alpha-minimal essential medium (Gibco, Grand Island, NY) supplemented of 20% fetal bovine serum and growth factors. Twice a week and at the end of culture supernatants were collected. After culture, the cells were filtered 40 μm (BD, Falcon) and stained for flow cytometry. In some experiments, proinflammatory cytokine production of healthy donors (HD) BM-CD34+ cells was evaluated upon TLR8 agonist (R848, 5 μM; InvivoGen, San Diego) response after 1 week of OP9-DL1 coculture.
HIV-RNA quantification
HIV-RNA in cultured CD34+ samples was quantified by using Abbott Real-time HIV-1 assay (Abbott Molecular, Inc., Des Plaines, IL), according to the manufacturer's instructions.
Cytokine detection
BM plasma samples were obtained after speed centrifugation for 10 min at 2,000 rpm and stored at −80°C. Supernatants on CD34+ cocultures were collected and stored at −80°C. Levels of proinflammatory cytokines (IL-8, IL-1β, macrophage inflammatory protein-1β, and granulocyte macrophage colony stimulating factor) on CD34+ coculture supernatants and plasma were analyzed by Bio-Plex Pro Assay (Bio-Rad Laboratories, CA) and Luminex platform (Luminex, Austin, TX), according to the manufacturer's instructions.
Statistical analysis
Results are expressed as median and range. Statistical significance was assessed by Mann–Whitney U-test by using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA). A p < .05 was considered statistically significant.
Results
Decreased frequency of CD34+Lin− cells from BM of HIV-infected patients
To analyze BM-HPCs compartment, we first assessed the frequency of HPC defined as CD34+Lin− cells (Fig. 1). The percentage of BM-HPC was found significantly lower in VR when compared with NVR patients: VR median 0.2% (IQR 0.16%–0.3%) versus NVR median 0.95% (IQR 0.8%–1.6%; p < .05). However, the percentage of BM-HPCs in NVR was lower than in HD (data not shown).

BM CD34+Lin− cells frequency from VR and NVR HIV+ patients.
BM-HPC lymphoid differentiation potential was partially restored in successfully ART-treated patients
The functionality of BM-HPC from HIV+-infected patients was analyzed in terms of lymphoid differentiation capability. To this aim, we cultured CD34+Lin− cells purified from 7 of 12 enrolled patients (4 NVR and 3 VR, Table 2, gray color) on OP9-DL1 stroma feeder cells for 16 days. We excluded two VR (No. 11, 12) for low viral load, cART exposure, and low yield of purified CD34+Lin− cells and three NVR (No. 5–7) for low yield of purified CD34+Lin− cells. The results indicated that the lymphoid differentiation potential of CD34+ BM-HPC was partially restored in successfully ART-treated patients. Indeed, BM-HPCs from VR patients (Fig. 2B, black bars) failed to differentiate toward T cells (CD4 and CD8). In contrast, in NVR patients (gray bars), the differentiation capability was improved, although remaining significantly lower than in HD (white bars). Specifically, the percentage of CD4 T cells mean was 48 ± 12 versus 4 ± 0.8 for NVR and VR, respectively, and the percentage of CD8 T cells mean was 26 ± 9 versus 2 ± 0.8 for NVR and VR, respectively.
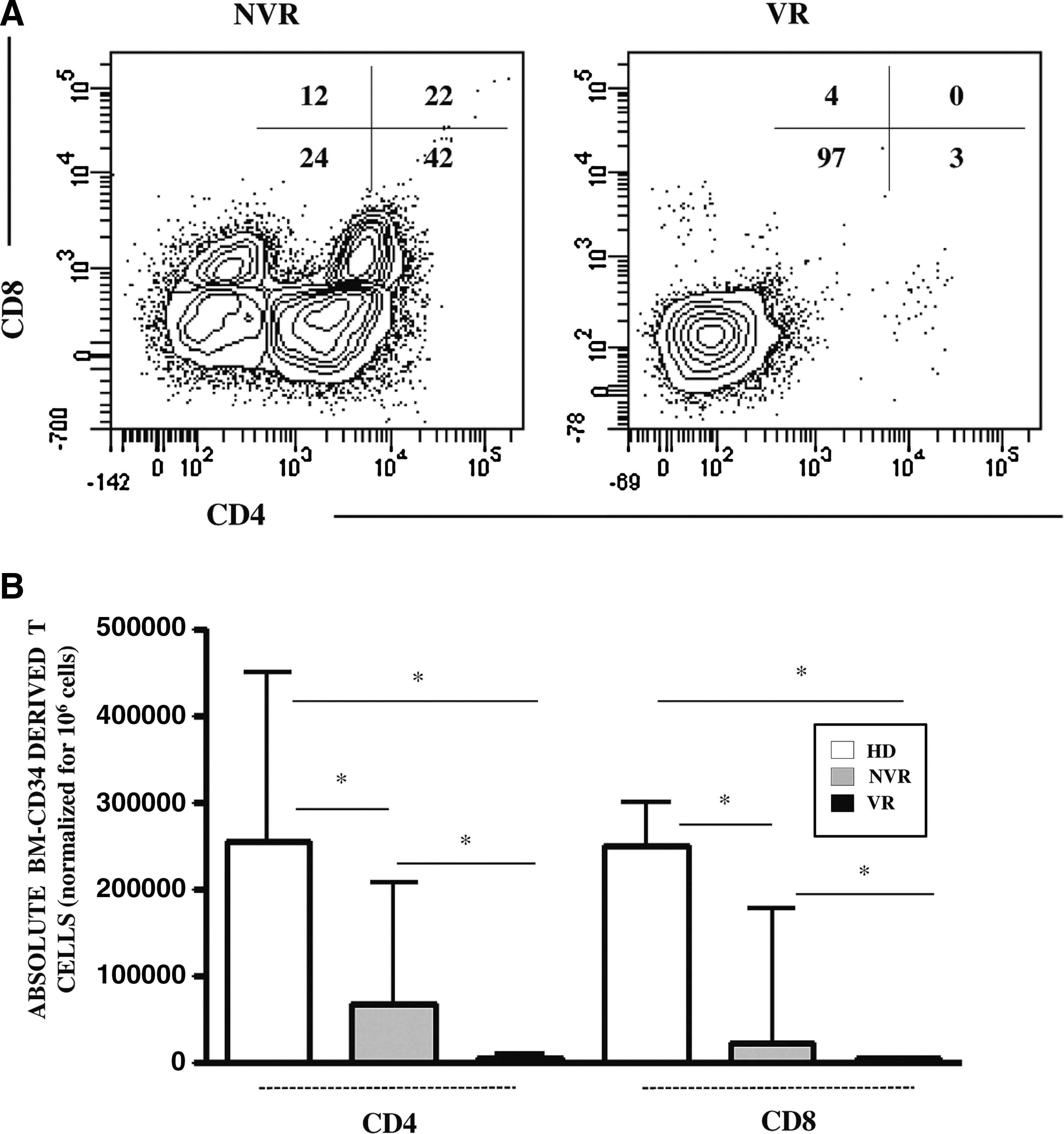
Lymphopoietic capability of BM-CD34+ cells isolated from HIV+ patients and HD after 16 days of coculture with OP9-DL1 cells.
Gray shade identifies patients used to evaluate BM-HPC in vitro differentiation potential.
BM, bone marrow; BMMCs, bone marrow mononuclear cells; nd, not done.
CD34+ cells from HIV-infected patients released proinflammatory cytokines during T cell differentiation
To evaluate the possible involvement of active HIV replication and/or of inflammatory profile in impairing CD34+ capability to differentiate toward T cells, HIV-RNA level and proinflammatory cytokines were quantified in BM-CD34+ HPCs cultures. Surprisingly, no active replication, measured as HIV-RNA, was detected in differentiating BM-CD34+ HPC cultures. In contrast, the ability of BM-CD34+ HPCs to produce proinflammatory cytokines was different in HD and HIV-infected patients. BM-CD34+ HPCs from HD produced very low levels of cytokines (Fig. 3). In contrast, in culture from HIV+ patients a significant increase in cytokines was observed in both NVR and VR patients: in particular, NVR patients presented a slight increase with respect to HD, whereas VR patients showed a massive increase when compared with both HD and NVR patients. We found a strict association between T cell differentiation capability and proinflammatory cytokine production. Indeed, we found a negative correlation between the number of differentiated CD4 or CD8 T cells and IL-8 (r = −0.86, p = .0023), GM-CSF (r = −0.80, p = .0036), and MIP-1β (r = −0.7, p = .01); no correlations was found with IL-1β.
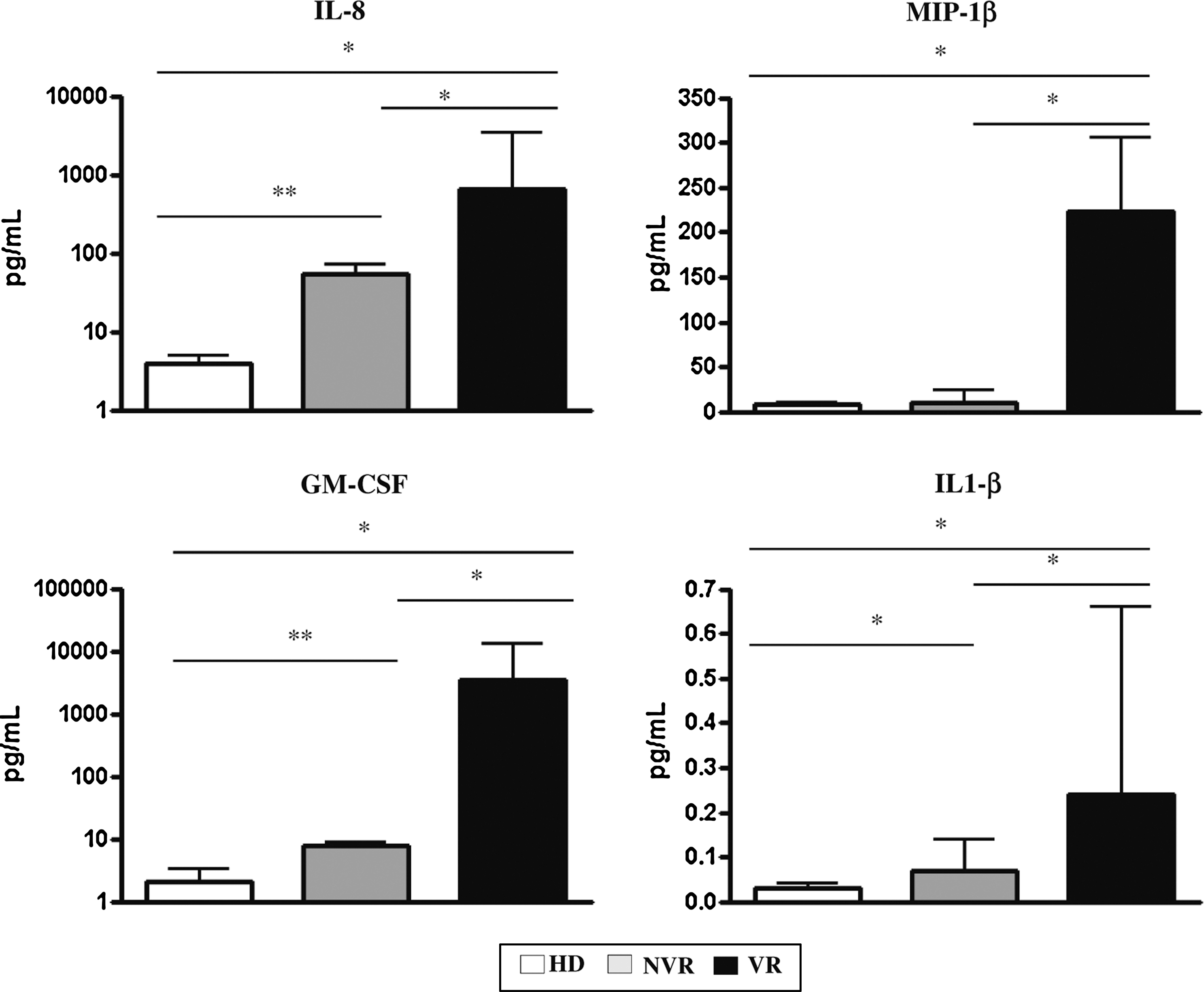
Proinflammatory cytokine production of BM-CD34+ cells during OP9-DL1 coculture. Supernatants were collected after 16 days from BM-CD34+ cells isolated from HD, NVR, and VR HIV+ patients cocultured with OP9-DL1 cells and assayed for cytokine concentration (IL-1β, IL-8, MIP-1β, GM-CSF) by Luminex assay. Histograms show the median and range of three HD, four NVR, and three VR samples in respective experiments. Mann–Whitney test, *p < .05, **p < .01. GM-CSF, granulocyte macrophage colony stimulating factor; IL, interleukin, MIP, macrophage inflammatory protein.
TLR8 triggering induced proinflammatory cytokine release by BM-HPC from HD affecting T cell differentiation
Therefore, we wondered whether the induction of proinflammatory cytokines by TLR8 triggering may impair T cell differentiation of BM-HPCs from HD. Thus, BM-HPCs from HD were stimulated with chemical TLR8 agonist R848, and after 1 week of OP9-DL1 coculture, the level of proinflammatory cytokines and the absolute number of T cell precursors were assessed. As reported in Figure 4A, TLR8 triggering significantly increased the release of IL-1β, MIP-1β, and GM-CSF from CD34+ BM-HPCs and determined a parallel and significant decrease of lymphoid commitment expansion (Fig. 4B, C, p < .05). Indeed, the percentage of L-HPC mean was 16 ± 3 versus 8 ± 1 for untreated and R848 stimulated, respectively, confirming a relationship between inflammatory microenvironment and different capability. In line with this finding, preliminary results obtained by triggering TLR8 on CD34 from one NVR patient confirms a relationship between proinflammatory cytokine production and CD4 and CD8 T cell differentiation impairment (data not shown).
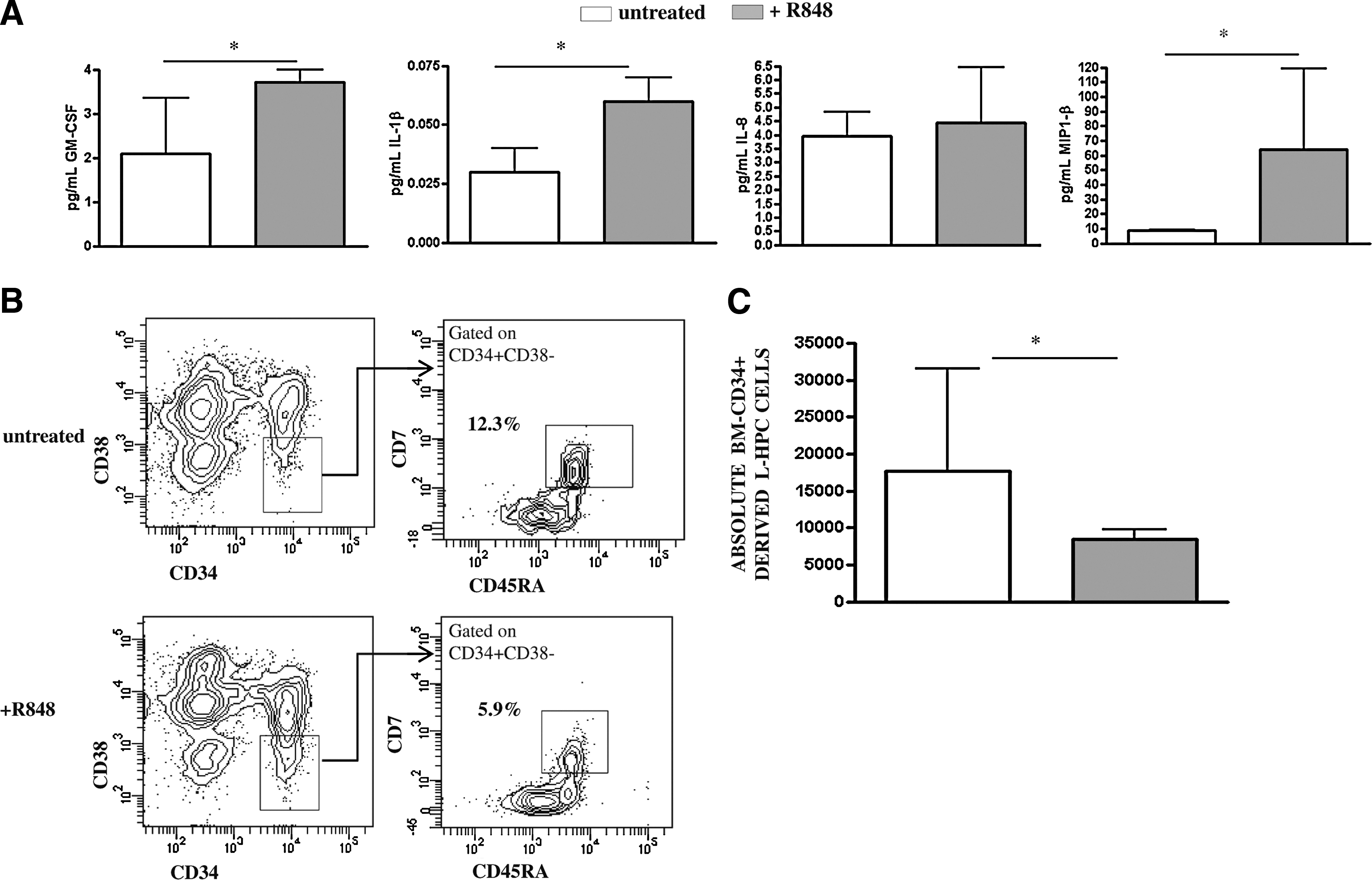
Relationship between proinflammatory cytokine production of BM-CD34+ cells by TLR8 triggering and T cell lineage differentiation potential.
Discussion
By using a previously described in vitro model of lymphopoiesis in vitro, 7,8 we were able to investigate for the first time the lymphoid differentiation potential of BM CD34+ cells isolated from VR and NVR HIV-infected patients. A number of studies have previously shown that BM-HPCs from HIV-infected patients have functional alterations 9 –11 and in HIV progressors display reduced capacity to generate CD4+ and CD8+ T cells in fetal thymic organ culture. 12,13 Moreover, it has been reported that, in an animal model, declining thymopoiesis in HIV infection may be related to impaired lymphopoiesis. 14,15 Overall, our results show that successfully ART-treated patients were able to promote an improvement in T cell commitment of BM-HPCs. These results agree with previous reports showing an improvement of BM-HPCs activity from cART-treated patients in clonogenic capability and cytokine production. 16
Several mechanisms may be taken into account in explaining the reduced lymphocyte differentiation of BM-HPCs from HIV-infected patients. Among these, viral replication as well as inflammatory microenvironment can play major roles. Recently we detected in BM-CD34+ cells HIV-DNA that correlated to plasma HIV viremia. 17 The absence of HIV-RNA in CD34+ cultures, as shown in the present study as well as in our previous report, 18 is in line with Durand et al., who did not find HIV-RNA in CD34+ cells upon cytokine-directed differentiation, 19 whereas is in contrast with the work of Carter et al. that revealed Gag protein expression in CD34+ cells upon differentiation. 20 There are several possible explanations for these conflicting data, such as CD34+ cells being infected at a level below the limit of detection in our system, and/or clinical features of patients determining different virus tropisms, and/or a possible contamination by other cell types. Further investigations seem necessary to clarify this issue.
Moreover, the absence of active HIV replication in CD34+ cultures from VR patients suggests that mechanisms other than those related to virus replication may affect the lymphopoietic capability of BM-HPCs. In fact, proinflammatory cytokines have been suggested as able to affect HPCs function, 21 but a correlation with HPC impairment has not been demonstrated. Cytokine-mediated selection of one HPC subset over another has been suggested as a clever way for the immune system to preferentially manufacture myeloid or lymphoid effector cells as appropriate. 21 Our results indicated that, during T cell differentiation, BM-HPCs from HIV+ patients produce proinflammatory cytokines, particularly higher in VR patients than in NVR. In particular, the release by CD34+ BM-HPC derived from VR patients of high levels of IL-8 and GM-CSF, cytokines able to promote the differentiation toward myeloid lineages, may explain their impairment to differentiate toward lymphoid lineage and T cells. 22,23 Moreover, it has been reported that the production of GM-CSF by CD34+ HPCs is in response to inflammatory stimuli. 24
It was reported that CD34+ HPCs can respond to several toll-like receptor agonist resulting in cytokine production and the generation of innate immune cells such as monocytes, macrophages, and dendritic cells. 25 –27 However, little is known about the response of HPCs to TLR agonist during T lineage differentiation. In this study, we showed that BM-HPCs from HD may respond to TLR8 triggering by increasing the production of proinflammatory cytokines, and decreasing T cell differentiation potential probably regulating myelopoiesis. Therefore, we can speculate that virus degradation products, able to trigger TLR8, 28 may induce BM-HPCs inflammatory response impairing their T cell differentiation potential.
Also if limited by a small sample size due to poor access to BM, our study depicts a new scenery on the mechanism possibly involved in the different lymphopoiesis capability of BM CD34+ HPC in NVR and VR patients. In our study, CD34+Lin− cells from BM of HIV-infected patients were able to produce proinflammatory cytokines in the absence of active viral replication, and this is associated with the reduced differentiation capability toward T cells. These data highlight a potentially new point of view on the multiple factors that may impair the immune reconstitution capability of treated HIV-infected persons.
Further critical issues in HIV-infected persons need to be evaluated, such as the molecular mechanisms triggering proinflammatory cytokine release by BM-HPCs, that may impair the immune reconstitution after antiretroviral treatment.
Footnotes
Acknowledgements
The authors gratefully acknowledge the patients who made this study possible. This work was supported by grants from Italian Ministry of Health [Ricerca Corrente] to INMI L. Spallanzani, IRCCS.
Author Disclosure Statement
No competing financial interests exist.