
Abstract
In the absence of an effective vaccine, strategies to prevent HIV transmission are urgently needed. Condomless receptive anal intercourse represents a major route of transmission, and efforts are being made to develop strategies, in which potent anti-HIV drugs are formulated for topical application to the rectum before sex. 5P12-RANTES is a promising candidate for such a purpose. It is an analog of the human chemokine RANTES/CCL5, which potently blocks CCR5, the principal coreceptor used by HIV to enter and infect target cells. As a protein, 5P12-RANTES is potentially vulnerable to attack by proteases in the rectal environment. In this study, we tested the stability of 5P12-RANTES on exposure to rectal lavage samples obtained from healthy volunteers, using a sensitive HIV entry inhibition assay as an indicator of stability. Varying levels of inactivating activity toward 5P12-RANTES were detected across the different lavage samples. Analysis of even the most aggressive samples indicated that protease activity in the rectal environment is unlikely to impact on the anti-HIV activity of 5P12-RANTES when applied pericoitally at the envisaged clinical dose (1 mM). This study indicates that 5P12-RANTES has adequate stability for further development as an HIV prevention drug for rectal use.
Introduction
N
Preexposure prophylaxis (PrEP) strategies are being developed, which involve delivering anti-HIV drugs to people at risk before sex. 4 PrEP products contain potent anti-HIV drugs formulated for either oral dosing or topical application to the mucosal tissues involved in sexual contact. Oral tenofovir disoproxil fumarate/emtricitabine was approved by the Food and Drug Administration for use in PrEP in 2012, 5 and a number of topical prevention products, including those specifically designed for pericoital rectal use, are in clinical development. 4
The chemokine receptor CCR5, which has a physiological role in the recruitment of leukocytes to sites of inflammation, is also the principal coreceptor used by HIV to enter and infect target cells. 6 Individuals homozygous for a null mutation in CCR5 are remarkably resistant to HIV infection, irrespective of the route of entry of the virus into the body. 7,8 Several studies using highly stringent models of mucosal viral challenge have shown that targeting of CCR5 is a valid strategy for topical HIV prevention. 9,10
Among the drugs targeting CCR5 that have been proposed for use in topical PrEP strategies are maraviroc, 11,12 a small molecule inhibitor approved for use in HIV therapy since 2007, 13 and 5P12-RANTES.14–-16 5P12-RANTES is a highly potent 14 and efficacious 15 69-residue analog of the human chemokine protein RANTES/CCL5, one of the natural ligands of CCR5. We have recently established a scalable low-cost cGMP manufacturing strategy 17 for producing clinical grade 5P12-RANTES in Pichia yeast to support the clinical development of 5P12-RANTES for use in both vaginal and rectal PrEP.
Stability both during storage and use is a key issue in evaluating protein-based topical PrEP strategies. We have previously shown that 5P12-RANTES is stable at elevated temperature, at vaginal pH, and in the presence of both human semen and human cervicovaginal lavage. 16 In this study, we investigated the stability of 5P12-RANTES in rectal lavage samples obtained from a panel of 25 healthy human subjects.
Materials and Methods
5P12-RANTES
We used clinical grade samples of recombinant 5P12-RANTES produced in Pichia yeast as described. 17
Rectal lavage samples
Lavage samples were obtained by infusing 50 mL of Normosol solution into the rectal vault via a flexible sigmoidoscope. 30 s later, 25–30 mL of lavage fluid was collected, filtered through a 0.2 μm filter, aliquoted, and stored at −80°C. In all cases, informed consent was obtained in writing from participants for biological sample collection, and the study had ethics committee approval.
Visual aspect, pH, and protein concentration
Aliquots of rectal lavage samples were thawed and assessed visually for suspended material and color intensity. Samples were taken for measurement of pH (pH strips, Machery-Nagel) and protein concentration (Quick Start Bradford protein Assay, Bio-Rad), and the remainder centrifuged for 3 min at 10,000 g.
Incubation of 5P12-RANTES with rectal lavage samples
5P12-RANTES was incubated with rectal lavage supernatants at concentrations of 10 μM and 1 mM. For the 10 μM 5P12-RANTES concentrations, 100 μL of rectal lavage supernatant was added to 25 μL of 5P12-RANTES diluted to 50 μM in PBS. For the 1 mM 5P12-RANTES concentrations, 1 mg of lyophilized 5P12-RANTES was added directly to 125 μL rectal lavage supernatant. Mixtures were incubated at 37°C for defined time intervals and then stored frozen at −80°C.
Cell fusion assay
An R5-tropic envelope-dependent cell-fusion assay was carried out as described 14,16,18 using HeLa-P5 L 19 and HeLa-Env-ADA 20 cell lines.
Fluorescein-casein protease assay
Protease activity of rectal lavage samples was assayed using a commercially available kit (Pierce) according to the manufacturer's instructions using a Flexstation microplate fluorimeter (Molecular Devices) with triplicate measurements taken every 20 s during a 20 min incubation at 37°C.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis and western blot analysis of 5P12-RANTES samples
Rectal lavage supernatants were incubated with 5P12-RANTES (10 μM) in PBS for 24 h at 37°C. Samples were separated on NuPAGE Novex 12% Bis–Tris gels. 5P12-RANTES was detected by western blot using a commercially available anti-RANTES/CCL5 biotinylated polyclonal antibody (R&D Systems) together with peroxidase-conjugated streptavidin (Roche), using SuperSignal West substrate (Pierce) with analysis performed using a Biomax MR system (Kodak).
Results
Initial screen
To favor detection of inactivating activity, our initial screen of each rectal lavage sample used 5P12-RANTES at a dilution 100-fold lower than the anticipated clinical concentration (10 μM instead of 1 mM): small losses represent a larger proportion of the initial amount and are thus more noticeable.
5P12-RANTES was incubated with rectal lavage samples for time intervals of 30 min, 1, 3, and 6 h at 37°C. Dilutions of these mixtures corresponding to initial 5P12-RANTES concentrations of 10−7.0 M, 10−7.7 M, 10−8.4 M, and 10−9.1 M were tested for anti-HIV activity using a highly sensitive cell-fusion assay 14,16,18 that is used as a potency assay for release of clinical grade batches of 5P12-RANTES 17 (Fig. 1).
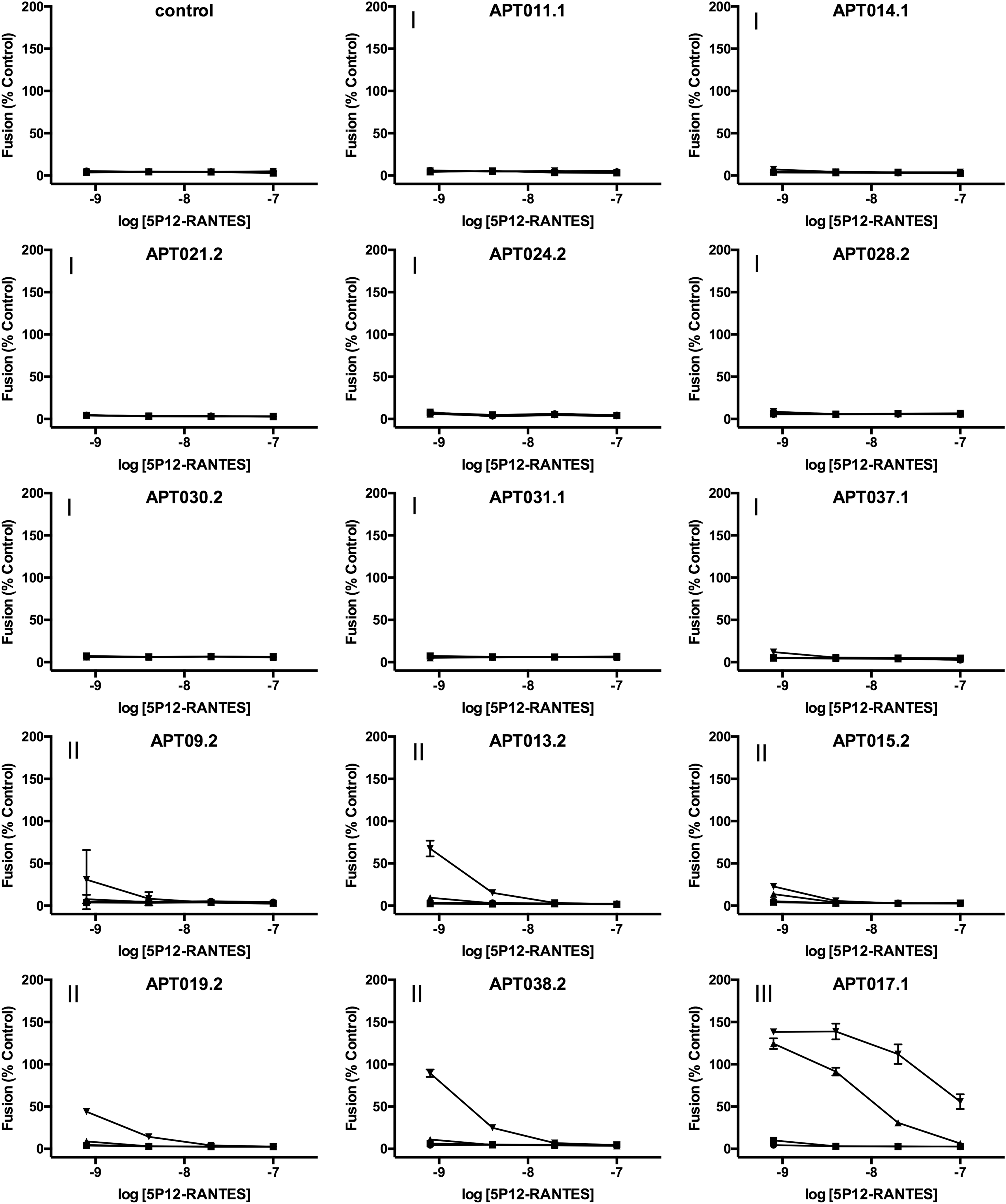
Inactivating activity of 26 rectal lavage samples against 5P12-RANTES. A cell-fusion assay was used to assess the anti-HIV activity of 5P12-RANTES following incubation (37°C) at 10 μM concentration with rectal lavage samples for 30 min (circles), 1 h (squares), 3 h (triangles), or 6 h (inverted triangles). Roman numerals indicate the group to which each lavage sample was assigned based on its activity (I = no effect, II = minor, III = intermediate, IV = aggressive).
Results obtained across the panel of lavage samples indicated striking heterogeneity, with inactivating activity ranging from entirely absent (e.g., sample APT011.1, no detectable loss of anti-HIV activity at any incubation time point) to aggressive (e.g., sample APT013.3, elimination of anti-HIV activity after only 30 min incubation).
To simplify further analysis, we divided the lavage samples into four different groups according to their levels of inactivating activity (see roman numerals in Fig. 1). The “No effect” group (I) contained lavage samples for which the 5P12-RANTES retained full inhibitory activity at all four incubation times. Lavage samples were assigned to the “Minor” group (II), when detectable loss of inhibitory activity was noted for the longest incubation time only (6 h), to the “Intermediate” group (III), when detectable loss of inhibitory activity was noted at both 3 and 6 h, and to the “Aggressive” group (IV), when loss of inhibitory activity was noted at 1, 3, and 6 h.
Characterization of lavage samples according to inactivating activity
Each lavage sample was next characterized for visual appearance, pH, and protein concentration to establish whether any of these parameters correlate with the inactivating activity of the samples toward 5P12-RANTES (Table 1).
Sample names (APTXXX.Y) indicate encoded donor identity (XXX) and sequence of sample donation (Y) if more than one sample was donated. The date of sample collection is indicated in brackets. Samples were grouped according to the level of inactivating activity toward 5P12-RANTES (Fig. 1), assessed visually (appearance) for yellow/brown color intensity (−colorless to +++most intense yellow/brown color), for pH using pH strips and for protein concentration by Bradford assay.
None of the rectal lavage samples showed any evidence of precipitated or suspended material. The color of the samples ranged from colorless to yellow/brown of varying intensity, presumably due to differences in levels of fecal-derived material. The pH of the samples lay in the range 6.0–8.8, consistent with values determined in the descending colon in previous studies. 21 Protein concentrations ranged between 0.08 and 1.59 μg/mL. The two lavage samples that had much higher protein concentrations than the others (APT032.1 and APT013.3) were also among the intensely colored samples and showed aggressive inactivating activity against 5P12-RANTES. However, other intensely colored lavage samples (e.g., APT024.2) had low protein concentrations, and across the whole group of samples, no obvious correlation was apparent between inactivating activity against 5P12-RANTES and color, or protein concentration, or pH (Fig. 2).
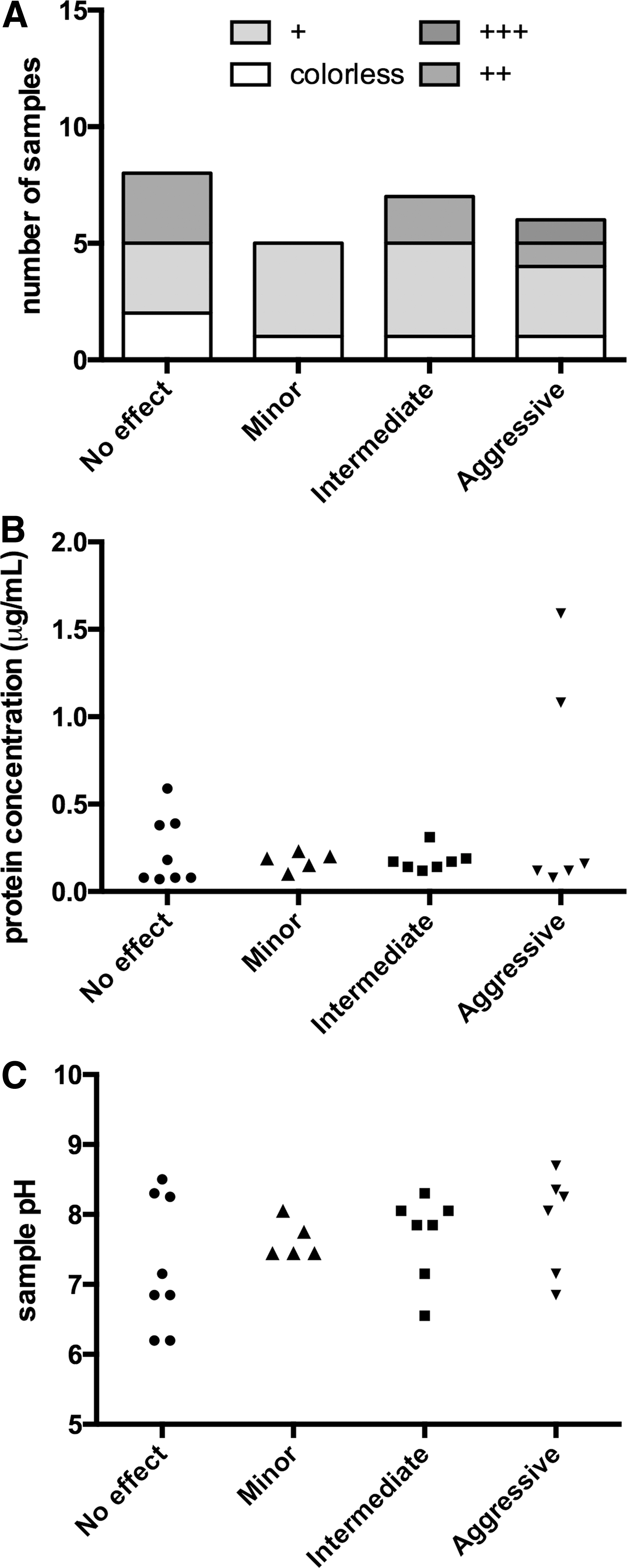
Color, protein concentration, and pH of rectal lavage samples.
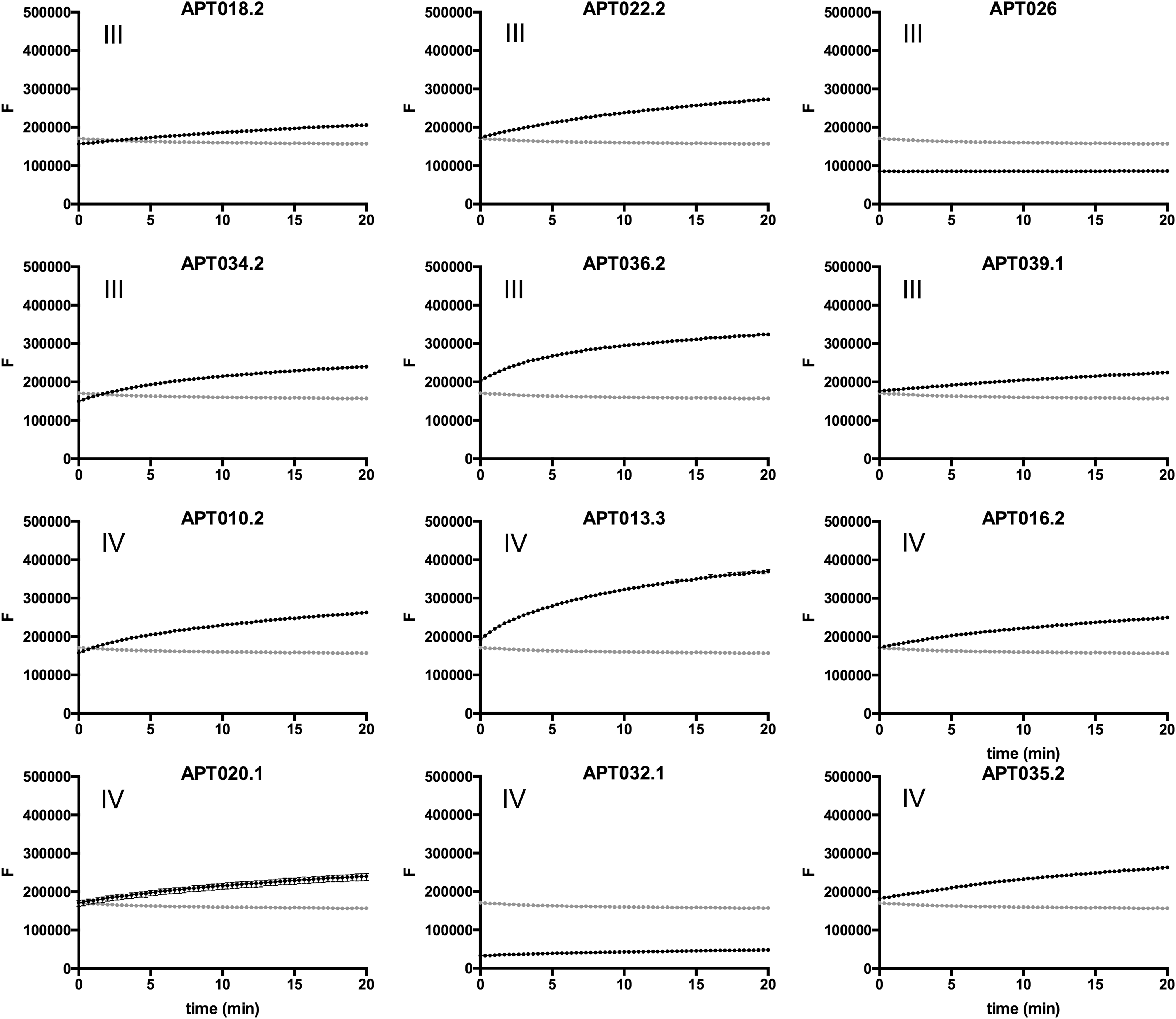
We next assessed the samples for overall protease activity in an assay using fluorescein-conjugated casein as a substrate (Fig. 3). A clear correlation emerged between the signals obtained in this assay for each lavage sample and corresponding inactivating activity against 5P12-RANTES (Fig. 1). Samples in the “no effect” group (I) exhibited undetectable levels of protease activity, samples in the “minor” group (II) showed low but detectable levels of protease activity, and samples in the more aggressive groups (III and IV) yielded the highest levels of protease activity. Hence, the differences in inactivating activity toward 5P12-RANTES observed across the panel of rectal lavage samples can reasonably be attributed to differences in protease activity.

Overall protease activity of rectal lavage samples. Samples or trypsin controls were added to fluorescein-casein substrate in 96-well plates and then transferred to a fluorescence plate reader, where measurements were taken every 20 s for 20 min. Baseline fluorescence (signal obtained when substrate is incubated with buffer alone) is indicated in gray. Typically, samples identified as having intense color (Table 1) gave signals partly or entirely below baseline; this presumably due to interference of sample components with the fluorescence signal. Roman numerals indicate to the 5P12-RANTES inactivating activity groups to which each sample was assigned (Fig. 1).
Protease-mediated inactivating activity against 5P12-RANTES could occur either through complete destruction of the protein or through truncation of sequences crucial for anti-HIV activity located at the N-terminus. 14 To investigate which of these processes is responsible, we incubated samples of 5P12-RANTES with either control buffer or selected rectal lavage samples and analyzed the resulting material by both sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and western blot using a polyclonal antibody capable of detecting not only RANTES/CCL5 but also 5P12-RANTES 17 and a range of N-terminally modified variants (unpublished data) (Fig. 4). As expected, bands corresponding to intact 5P12-RANTES were observed after incubation with samples previously shown to have no detectable inactivating activity (samples APT024.2 and APT037.1). Less intense bands of apparently intact protein were apparent after incubation with one sample with “minor” inactivating activity (APT015.2) as well as with one of the five samples with “intermediate” inactivating activity (APT026). No detectable bands of intact protein remained after incubation with the other four out of five samples with “intermediate” inactivating activity (APT017.1, APT022.2, APT036.2, and APT039.1) or with the most aggressive sample (APT013.3). Hence, the predominant route of protease-mediated inactivation of 5P12-RANTES appears to be via total destruction of the protein except, possibly, for sample APT026. Despite its relatively high inactivating activity, it shows relatively low levels of overall protease activity (Fig. 3) and moderate reduction of the intensity of the band corresponding to 5P12-RANTES in SDS-PAGE and western blot and may involve N-terminal truncation.
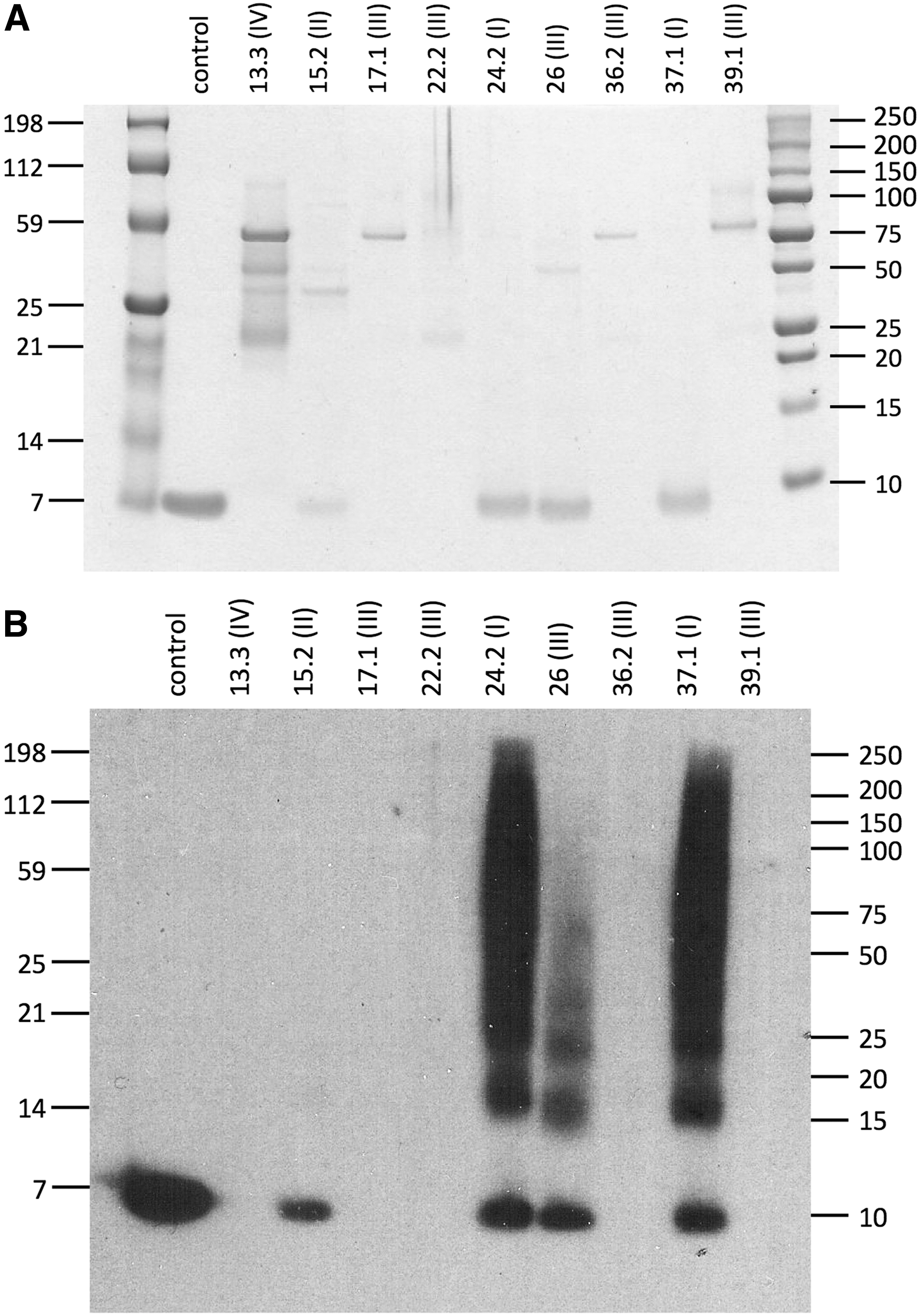
Effects of rectal lavage protease activity on the integrity of 5P12-RANTES. Samples of 5P12-RANTES were incubated (37°C) at 10 μM for 24 h with the indicated rectal lavage samples, and then subjected to analysis by SDS-PAGE.
Inactivating activity of the aggressive rectal lavage samples against 5P12-RANTES at the envisaged clinical dose
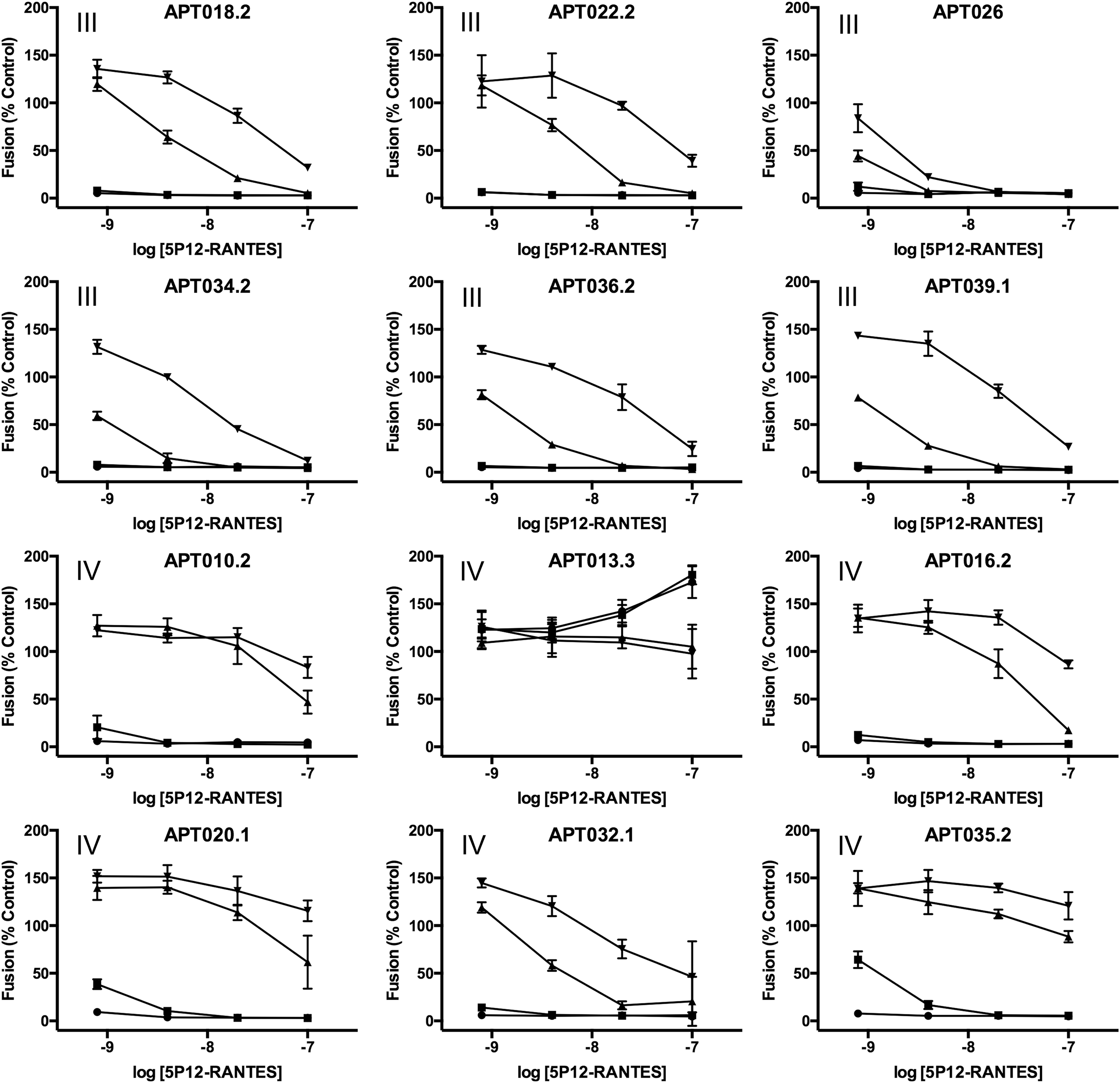
We next chose four samples from the “Aggressive” group (APT013.3, APT016.2, APT020.1, and APT035.2) and incubated them with 5P12-RANTES at the envisaged clinical dose, 1 mM. Incubation mixtures were sampled at defined time points and assessed for anti-HIV activity in the cell fusion assay over a full concentration range (nine dilutions spanning the range 10−7 to 2.6 × 10−13 M). The incubation time points chosen for rectal lavage samples APT016.2, APT020.1, and APT035.2 were 3 and 12 h, while those chosen for rectal lavage sample APT013.3, which exhibited particularly aggressive activity in the preliminary screen, were 30 min and 1 h.
To take account of the intrinsic intra-assay variability characteristic of cell-based assays in general and of our cell fusion assay in particular, we set up 12 independent dose–response experiments for reference standard samples in parallel (Fig. 5A). In accord with results previously obtained with 5P12-RANTES in this assay, 14 the IC50 values lay in the low picomolar range (upper and lower limits of potency 7 pM and 26 pM, respectively).

Effect of 5P12-RANTES concentration on inactivation by aggressive rectal lavage samples. Anti-HIV activity of 5P12-RANTES using a cell-fusion assay
For the test incubations (Fig 5B), none of the IC50 values for samples incubated at the shorter of the two incubation times chosen lay outside the range determined for the reference standard samples (fitted IC50 values [pM] of 16, 26, 22, and 22, respectively, for lavage samples 13.3, 16.2, 20.1, and 35.2). However, there were signs of degradation on longer incubation (fitted IC50 values [pM] of 46, 23, 33, and 61, respectively, for lavage samples 13.3, 16.2, 20.1, and 35.2).
Discussion
Heterogeneity in inactivating activity across the panel of rectal lavage samples
Under conditions chosen to favor detection of low levels of degradation, we noted a wide range of inactivating activity toward 5P12-RANTES across the panel of rectal lavage samples (Fig. 1): eight samples showed no detectable inactivating activity, twelve samples showed minor/intermediate levels, and six showed the highest levels of inactivating activity. These differences did not correlate with color, pH, or protein content of the samples (Fig. 2), but strongly correlated with their overall protease activity (Fig. 3). Analysis of the structural integrity of 5P12-RANTES by SDS-PAGE and western blot following incubation with different rectal lavage samples indicates that inactivation occurs mostly via total destruction of the protein rather than truncation of its N-terminal anti-HIV pharmacophore 14 (Fig. 4).
The most likely explanation for this heterogeneity across the panel of donors would be differences in their rectal microbiomes. The rectal environment contains proteolytic enzymes of both host and microbiotal origin, with the latter contributing the majority of protease activity. 22 Human gut microbiomes typically contain several thousand different detectable bacterial species, 23 with even more variation detectable both between individuals 23,24 and within an individual over time. 25 Large differences in fecal proteolytic activity across groups of individuals have been noted 22 and explained in terms of differences in gut colonization by bacterial taxa that produce high levels of active proteases. In this regard, it is interesting to note that in a previous study, 26 evaluation of the same panel of rectal lavages revealed quite striking differences in levels of nuclease activity against anti-HIV RNA aptamers, and it was proposed that these were attributed to microbiotal differences in the rectal lavages of the different samples.
A single subject in the study, APT013 donated two different samples (APT013.2 and APT013.3), the first preceding the second by ∼12 weeks. These two samples showed striking differences in both overall protease activity (low for APT013.2 and among the highest for APT013.3, Fig. 3) and inactivating activity toward 5P12-RANTES (minor for APT013.2 and aggressive for APT013.3, Fig. 1), indicating that proteolytic activity in the rectal environment of a given individual can vary significantly over time, most likely due to changes in the intestinal microbiota.
Implications for use of rectal gel containing 5P12-RANTES
The aggressive protease activity shown by certain rectal lavage samples analyzed in this study would be unlikely to impact on the efficacy of 5P12-RANTES in its intended clinical use. When these samples were tested on 5P12-RANTES at the envisaged clinical concentration (1 mM) and at time points compatible with pericoital use (30 min to 3 h), none of the cell-fusion assay curves lay outside the range of the assay curves for untreated reference standard samples of 5P12-RANTES (Fig. 5). It should, however, be noted that (i) proteolytic enzymes responsible for degradation would be expected to be found at a higher concentration in undiluted rectal fluid than in the lavage samples, (ii) 5P12-RANTES would be diluted from its initial concentration of 1 mM following rectal application. Furthermore, the gel formulation used could have an impact (probably negative, but possibly positive) on the access of the proteolytic enzymes to 5P12-RANTES, and their cleavage activity on it as a substrate. Hence, it will only be possible to obtain certainty as to the stability of 5P12-RANTES in the rectal environment during Phase 1 clinical studies, where pharmacokinetic and pharmacodynamics studies would be carried out following rectal administration of gel-formulated 5P12-RANTES to healthy volunteers. If a sustained release rectal formulation were to be explored, given that we noted signs of degradation after longer incubations, incorporation of a protease inhibitor such as aprotinin would be a potentially useful strategy.
Importantly, however, even in the presence of the most aggressive rectal lavage samples, 5P12-RANTES shows encouraging stability with respect to its envisaged pericoital formulation.
Footnotes
Acknowledgment
This work was supported by funding from the Swiss National Science Foundation (310030_143789 to O.H.).
Author Disclosure Statement
No competing financial interests exist.