
Abstract
Persistent immune activation in human immunodeficiency virus (HIV) infection is responsible for alterations in immune system such as activation, apoptosis, and reduced frequencies. Reduced immune activation is known to be associated with virus control. Limited information is available on the influence of pan-immune activation on memory responses. Hence, we compared the T cell activation and memory profile in HIV-infected individuals exhibiting disease control such as long-term nonprogressors (LTNPs) and progressors. The activated CD4+ and CD8+ T cells were significantly lower and the CD4+ and CD8+ central memory T cell phenotypes were significantly higher in the LTNPs compared to the progressors. In addition, we observed significant inverse association between the T cell activation and frequencies of central memory T cells. Our findings indicate that patients with absence of disease progression have preserved central memory T cell population associated with lesser immune activation.
H
We used HIV-infected long-term nonprogressors (LTNPs) as a model for control over HIV disease progression. LTNPs represent a group of asymptomatic, HIV-infected individuals with stable CD4 count of ≥500 cells/mm3 for at least seven or more years of documented HIV infection in the absence of antiretroviral treatment (ART). 9 LTNPs are shown to have a variable degree of immune activation in different studies. 10 –12 It was reported that despite a high viral load, the lower immune activation and preserved immune responses result in long-term asymptomatic disease stage in patients of Amsterdam cohort, 10 while chronic immune activation has been shown to be associated with progressive HIV. 13,14 Furthermore, the persistent immune activation is shown to drive the T cell differentiation resulting in replicative senescence. 15 Hence, we hypothesize that lower immune activation of T cells in LTNPs may ultimately result in better preservation of memory T cell population compared to the chronic progressors having a higher percentage of activated T cells. To test this hypothesis, we assessed the frequencies of activated T cells expressing HLA-DR and CD38 as a marker of immune activation, as well as the frequencies of central (TCM) and effector memory (TEM) T cells in LTNPs.
Thirty-one (11M/20F) LTNPs were recruited from the LTNPs cohort who are under regular follow-up at National AIDS Research Institute along with 21 (11M/10F) chronic progressors (ART-naive HIV-infected patients with CD4 count <500 cells/mm3) attending the outpatient clinics of the institute. The median age was 38 years in both LTNPs (range 30–75 years) and chronic progressors (range 22–48 years). The median seropositivity period of LTNPs was 11 years (range 8–19 years). At the time of enrolment in this study, the median CD4 count was 630 and 300 cells/mm3 in LTNPs and progressors, respectively. The median plasma viral load (pVL) was significantly lower (5,321 copies/ml; range 40–770,523) in LTNPs than that in progressors (28,345 copies/ml; range 1,065–1,456,931) (p = .012). The participants were not tested for M. tuberculosis infection; however, none of them presented with any signs or symptoms of tuberculosis disease. In addition, five HIV-seronegative healthy individuals (healthy controls; range 28–33 years; 3M/2F) with self-reported history of no fever in last 2 weeks and no major systemic disease were also enrolled for comparison. The study was approved by the Institutional Ethics Committee and prior written informed consent was obtained from all the participants.
The frozen peripheral blood mononuclear cells (PBMCs) from the study participants were revived as described previously
16
and viability of the cells was assessed using trypan blue dye exclusion method. The samples showing >90% viability and >3 × 106 cell count were considered for further analysis. 1 × 106 PBMCs each were used to characterize activation status of T cells and determine frequencies of memory CD4+ and CD8+ T cells using multicolor flow cytometry. The following antibodies were used: anti-CD3-PE-Texas-Red-A (Invitrogen), anti-CD4-APC-Cy7, anti-CD8-PE-Cy7, anti-CD38-FITC, anti-HLA-DR-PerCP-Cy5.5, anti-CD45RA-APC, CD67L-PE, and CCR7-PE-Cy7 (all from BD Pharmingen). PBMCs were incubated with specific antibodies for 30 min in dark at RT, followed by washing with a wash buffer (phosphate-buffered saline +0.5% fetal calf serum). The cells were fixed in 3% formaldehyde, acquired, and analyzed on FACSAria (Becton Dickinson) using FlowJo software. A total of 50,000 gated events of lymphocytes were acquired. Appropriate cell and isotype controls were used for determination of CD38/HLA-DR/HLA-DR and CD38 coexpressing CD4+ and CD8+ T cells and to determine the frequencies of CD4 and CD8 central memory (CCR7+/CD62L+ and CD45RA−) and effector memory (CCR7−/CD62L− and CD45RA−) population. The gating strategy to identify the different populations is shown in Supplementary Figure S1 (Supplementary Data are available online at
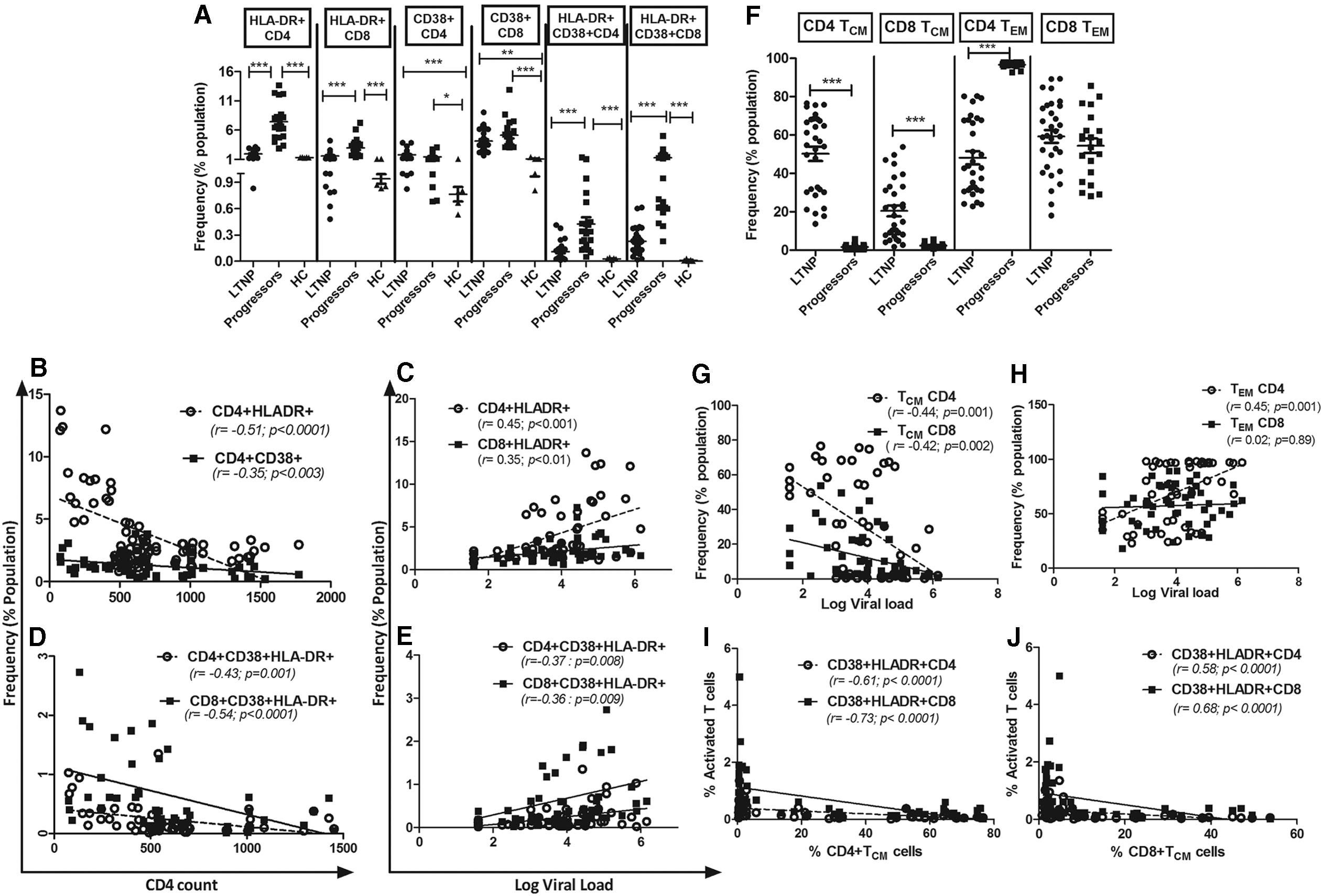
Characterization of CD4+ and CD8+ T cells among Indian LTNPs and chronic progressors.
The progressors had a significantly higher expression of activation markers, HLA-DR, CD38, and HLA-DR+CD38 on CD4 and CD8 T cells compared to the healthy controls (p < .05; Fig. 1A). Progressors also had a higher expression of HLA-DR and HLA-DR+CD38 on both CD4+ and CD8+ T cells compared to the LTNPs (p < .001; Fig. 1A), but not CD38-expressing T cells (p > .05; Fig. 1A). The LTNPs had similar frequencies of HLA-DR as well as HLA-DR+CD38-coexpressing CD4 and CD8 T cells compared with healthy controls (p > .05; Fig. 1A), except the expression of CD38, which was significantly higher in LTNPs than in healthy individuals (p < .05; Fig. 1A). We also determined whether the immune activation had any association with age within each study group and found that the frequency of only CD38-expressing CD4 cells was positively associated with age in LTNPs (p < .0001; r = 0.50), while the T cell activation (HLA-DR- and/or CD38-expressing T cells) was not associated with age among the progressors. The HLA-DR+ and CD38+ CD4 T cell frequencies were negatively associated with CD4 cell counts (p < .0001 and p < .003, respectively) (Fig. 1B). HLA-DR-expressing CD4+ and CD8+ T cells were associated with higher pVL (p < .001 and p < .01, respectively) (Fig. 1C). Higher pVL was also found to be associated with CD38+CD8+ T cells, close to the significance levels (p = .056; r = 0.268). Furthermore, the frequencies of T cells coexpressing HLA-DR+CD38 were negatively associated with CD4 cell count (Fig. 1D) and positively with pVL (Fig. 1E).
The frequencies of both CD4+ and CD8+ TCM cells were significantly higher in LTNPs than progressors (p < .0001) (Fig. 1F) and were associated with lower pVL (p = .001 and p = .002, respectively) (Fig. 1G), but not with CD4 count. In contrast, the frequencies of CD4+ TEM cells were significantly lower in LTNPs than in the progressors (p < .0001) (Fig. 1F) and significantly associated with higher pVL (p = .0009) (Fig. 1H). The CD8+ TEM frequencies were similar in progressors and LTNPs (Fig. 1F). We further assessed the association between the immune activation (activated T cells) and memory T cell population. A strong inverse correlation was observed between the activated CD4+ and CD8+ T cells expressing HLA-DR+CD38+ with CD4+ and CD8+ TCM (Fig. I, J respectively), while the CD4+TEM frequency was positively associated with HLA-DR+CD4+ (r = 0.594; p < .0001) and CD8+ T cells (r = 0.435; p = .0001).
Studies to identify correlates of immune mechanisms of HIV control are utmost essential for the development of new immune therapies as well as protection strategies. We, in this study, assessed the influence of immune activation on the memory T cell population that is important in mounting efficient recall responses. The study showed that the LTNPs exhibit lower T cell activation, higher CD4+ and CD8+TCM, and lower TEM populations compared to the chronic progressors. We also showed that the immune activation correlated with lower frequencies of TCM cells. A number of studies have shown that early immune activation causes damage to the immune system, which could result in chronic immune activation and further loss of immune function leading to disease progression. 4,10,13,14,17 The CD38- and HLA-DR-expressing T cells are being used as a marker of immune hyperactivation and disease progression. 4 These studies support our finding that LTNPs show less activation of T cells, indicating low inflammatory environment and thus preservation of CD4 count, and control over disease progression.
Strong TCM cell responses have shown to be associated with controlled disease progression. 10,18 –20 Our Indian LTNP population also showed higher TCM population, supporting these observations. We also confirmed the previous finding of association of TEM with faster disease progression. 6 The association of TEM frequencies with higher immune activation is also supported by previous observation in a mouse model where the immune activation showed a progressive conversion of naive T cells into effector memory cells. 20
The increased immune activation is known to drive T cell differentiation leading to replicative senescence and immunodeficiencies. 15,21 In HIV infection, this scenario in early course of HIV infection might result in loss of virus control leading to disease progression. During early HIV infection, higher TCM and TEM are associated with low and high viral load set point, respectively, indicating that early decrease in TCM cells might be contributing to high viral load and thus faster disease progression. 6 In support to these previous observations, our study has shown that preserved TCM frequencies and lower immune activation are associated with controlled disease progression, although the causal relationship could not be established. It might be possible that lower immune activation in LTNPs is responsible for reduced perturbation in T cell differentiation and further apoptosis of T cells, resulting in preserved central memory T cells, which can effectively respond to the antigenic stimulus. This scenario might be collectively and/or partially contributing to the sustained CD4 counts in these patients even in the presence of high viral load. Indeed, the natural host of SIV infection, the sooty mangabey monkeys, do not develop AIDS despite high levels of virus replication and exhibit very low levels of immune activation. 22
In summary, the low immune activation in HIV infection may contribute to preservation of memory cells and thus control over disease progression. The immune interventions to reduce the immune activation might be one of the useful anti-HIV strategies.
Footnotes
Acknowledgments
The authors would like to thank the study participants and the staff of clinic and immunology and virology laboratory of National AIDS Research Institute for their assistance.
Financial Support
This work was supported through institutional funding.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.