
Abstract
Both Caspase 1-induced cell death and Caspase 3-induced cell death were reported to be the causes of CD4+ T cell depletion in HIV infection. We measured by flow cytometry the expression of key proteins associated with pyroptosis (Caspase 1), apoptosis (Caspase 3, Caspase 8, Caspase 9), and immune activation in peripheral T cells. The percentages of CD4+ T cells that expressed Caspase 1 and Caspase 3 were significantly higher in untreated human immunodeficiency virus 1 (HIV-1) patients compared with healthy control (Caspase 1: 19.40% vs. 4.65%, p = .006; Caspase 3: 12.75% vs. 4.18%, p < .001). However, the percentages of Caspase 3 in CD8+ T cells increased significantly, while the percentages of Caspase 1 in CD8+ T cells did not change significantly (Caspase 1: 3.33% vs. 1.99%, p = .821; Caspase 3: 20.35% vs 4.74%, p < .001). The percentages of HLA-DR+ CD38+ CD8+ T cells were positively correlated with those of Caspase 1+ CD4+ T cells, but not with those of Caspase 3+ CD4+ T cells. After highly active antiretroviral therapy, the percentages of Caspase 1, but not of Caspase 3, -expressing CD4+ T cells decreased to a level comparable with those of healthy controls (Caspase 1: 6.05% vs. 4.65%, p = .514; Caspase 3: 9.67% vs. 4.18%, p < .001). Our study indicated that CD4+ T cells experience both pyroptosis and apoptosis, while CD8+ T cells undergo only apoptosis in HIV-1 infection. Pyroptosis, but not apoptosis, in CD4+ T cells may be inhibited by effective antiretroviral therapy.
Introduction
T
Bystander CD4+ T cells are abortively infected with HIV-1, and short HIV DNA transcripts accumulate in the cytosol. 10 The DNA sensor, IFI16, detects these viral DNAs and activates Caspase 1 to induced pyroptosis characterized by plasma membrane pore formation and cytoplasmic content extrusion. 8,11 –14
Apoptosis can be initiated through two distinct pathways. In the intrinsic apoptosis pathway, HIV-1 infection leads to the release of cytochrome c from CD4+ T cell mitochondria, leading to activation of Caspase 9 and subsequent Caspase 3 activation. 15 In the extrinsic pathway, Fas receptor ligand (FasL) combined with Fas triggers Caspase 8 activation, resulting in subsequent Caspase 3 activation. 16,17 Caspase 3 activated by either pathway induces apoptosis in CD4+ T cells. 18
Apoptosis and pyroptosis may be the two main causes for decreased CD4+ T cells during HIV-1 infection. In this study, we showed that the percentages of T cells undergoing pyroptosis and apoptosis in HIV-1-infected patients were elevated before combination highly active antiretroviral therapy (HAART) and that Caspase 1 activity, but not Caspase 3, was significantly downregulated following HAART.
Materials and Methods
Participants
Forty HIV-1-infected patients naïve to antiretroviral therapy, 40 HIV-1-infected patients with undetectable plasma viral loads for more than 1 year after HAART, and 16 healthy controls were enrolled at Shanghai Public Health Clinical Center, China. HIV-1-infected patients with opportunistic infections, hepatitis B virus (HBV), or hepatitis C virus (HCV) infection were excluded. Healthy controls with HIV-1, HBV, or HCV infection or with medicine were excluded. Ethical approval was granted by the Ethics Committee of Shanghai Public Health Clinical Center (approval number: 2013-K006). Informed written consent was obtained from each volunteer.
Peripheral blood mononuclear cell preparation
Peripheral blood mononuclear cells (PBMCs) were isolated from 20 mL heparinized peripheral blood by centrifugation over a Ficoll-Histopaque gradient (Fresenius Kabi, Halden, Norway) and washed with phosphate-buffered saline.
T cell immunophenotyping and intracellular staining
The following monoclonal antibodies were used for T cell immunophenotyping: CD3-PerCP-Cy5.5 (Biolegend, London, United Kingdom); CD4-BV605, CD8-APC-H7, CD45RA-APC, CD38-PE-Cy7, HLA-DR-PE, Fas-PE-Cy7, FasL-PE, Annexin V-fluorescein isothiocyanate (FITC), 7-AAD, IgG2a-PE, and IgG-PE-Cy7 (BD Biosciences, San Jose, CA). The following antibodies were used for intracellular staining: CaspGLOW Fluorescein Active Caspase 3 Staining Kit, CaspGLOW Fluorescein Active Caspase 8 Staining Kit, and CaspGLOW Fluorescein Active Caspase 9 Staining Kit (eBioscience, San Diego, CA); and Caspase 1 (activated P10)-FITC (Bioss, Inc., Woburn, MA).
PBMCs were incubated with antibody cocktails (cocktail 1: CD3, CD4, CD8, CD45RA, CD38, and HLA-DR; cocktail 2: CD3, CD4, CD8, CD45RA, Fas, and FasL; cocktail 3: CD3, CD4, CD8, and CD45RA; cocktail 4: CD3, CD4, CD8, and CD45RA; cocktail 5: CD3, CD4, CD8, and CD45RA; and cocktail 6: CD3, CD4, CD8, CD45RA, IgG2a, and IgG) for 20 min at 4°C. After washing, 2 × 106 cells in cocktails 1, 2, 3, and 4 were further stained with the Active Caspase 3 Staining Kit, Active Caspase 8 Staining Kit, Active Caspase 9 Staining Kit, and Caspase 1 (activated P10)-FITC in accordance with manufacturer instructions to detect Caspase 3, Caspase 8, Caspase 9, and Caspase 1. Cells were then washed and fixed with 1% formaldehyde and stored at 4°C. Cells (2 × 106) in cocktail 5 were further stained with Annexin V and 7-AAD, followed by analysis using an LSRFortessa flow cytometer (BD Biosciences). The data were analyzed using FlowJo, version 7.6.4 (FlowJo, LLC, Ashland, OR), and two populations were identified: naive (CD4+CD45RA+ and CD84+CD45RA+Tn) and memory (CD4+CD45RA− and CD4+CD45RA−Tm). Expression of Caspase proteins was determined based on fluorescence minus one, while CD38, HLA-DR, Fas, and FasL expression was determined based on matched isotype controls. The flow cytometry gating strategy for Caspase-1- or Caspase-3-positive cells is shown in Supplementary Figure S1 (Supplementary Data are available online at
Measurements of HIV-1 RNA in peripheral plasma
HIV-1 RNA was quantified using the plasma COBAS AmpliPrep/COBAS TaqMan HIV-1 Qualitative Test, version 2.0 (Roche, Basel, Switzerland), according to manufacturer instructions. The lower limits of detection of the assays were 20 copies/mL.
Statistical analyses
The Mann–Whitney U test was used to compare statistical significance between groups. Spearman's rank correlation coefficient was used to assess associations among CD4+ T cell counts, HIV-1 RNA, and immunologic measurements. Normally distributed data are shown as mean ± standard deviation, while non-normally distributed data are shown as median (interquartile range). Analyses were two-sided, and p < .05 was considered significant. All statistical analyses were performed with SPSS, version 16.0 (SPSS, Inc., Chicago, IL), and GraphPad Prism, version 5.0, software (GraphPad Software, San Diego, CA).
Results
Patient characteristics are shown in Table 1.
HAART, highly active antiretroviral therapy; IQR, interquartile range; N/A, not available; NA, not applicable; NNRTIs, non-nucleoside reverse transcriptase inhibitors; NRTIs, nucleoside reverse transcriptase inhibitors; PIs, protease inhibitors.
Caspase 1 expression increased in CD4+ T cells, but not in CD8+ T cells, of HIV-1-infected patients
The percentage of CD4+ T cells expressing Caspase 1 was significantly higher in untreated HIV-1-infected patients relative to that of healthy controls [19.40% (6.36%–27.00%) vs. 4.65% (3.85%–6.88%); p = .006]. However, the percentage of CD8+ T cells expressing Caspase 1 was unchanged in HIV-1-infected patients compared with that observed in healthy controls [3.33% (1.20%–6.91%) vs. 1.99% (1.58%–5.56%); p = .821] (Fig. 1A, B).
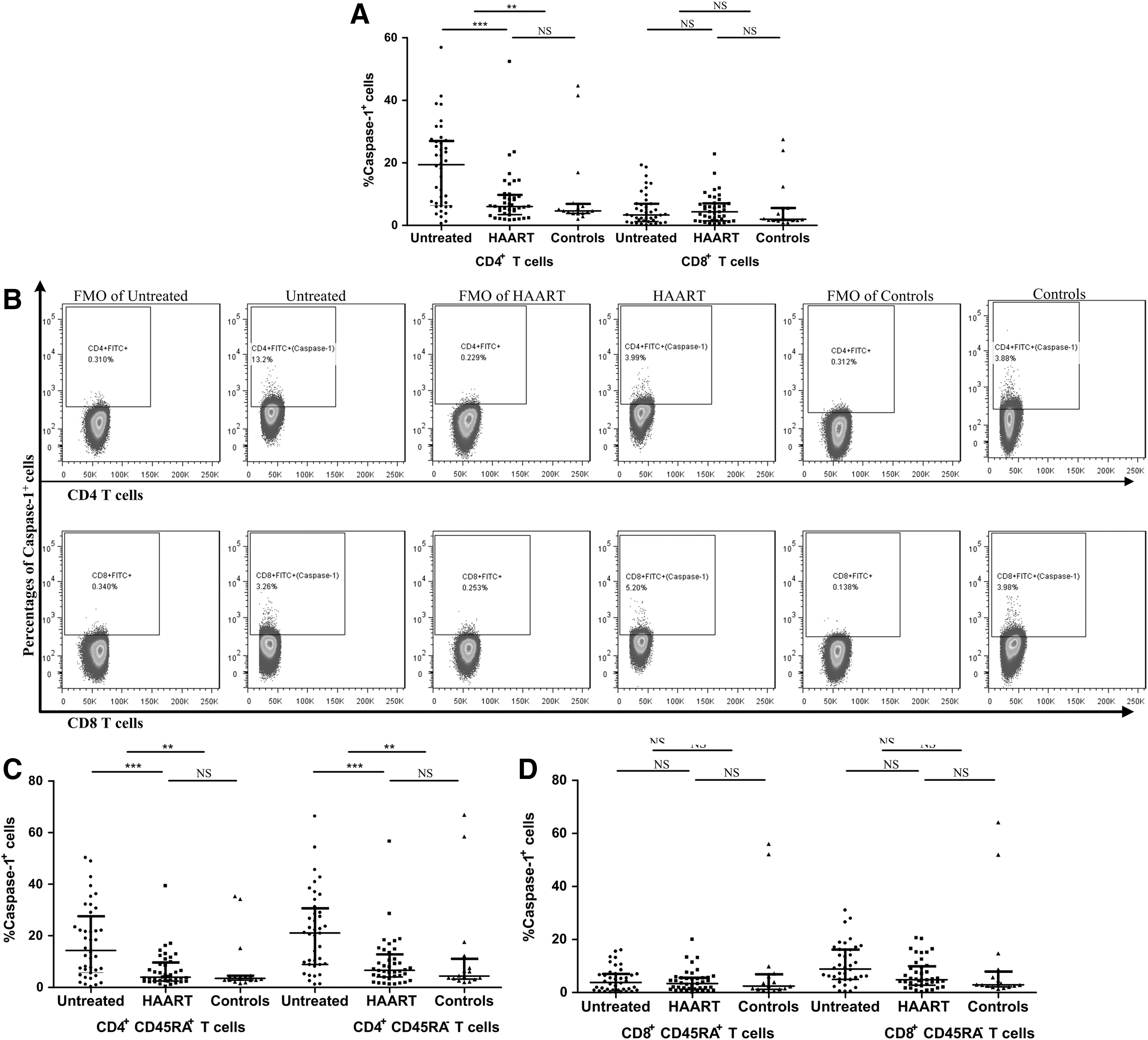
Percentages of CD4+ and CD8+ T cells expressing Caspase 1 from untreated HIV-1-infected patients, HAART patients, and healthy controls. Caspase 1 expression increased in CD4+ T cells, but not in CD8+ T cells, of HIV-1-infected patients
Similar results were observed in both memory CD4+ T cells and naive CD4+ T cells and memory CD8+ T cells and naive CD8+ T cells (Fig. 1C, D).
Caspase 3 expression increased in both CD4+ and CD8+ T cells in HIV-1-infected patients
The percentages of CD4+ and CD8+ T cells expressing Caspase 3 were both significantly higher in untreated HIV-1-infected patients compared with those of healthy controls [CD4+: 12.75% (9.11%–24.20%) vs. 4.18% (3.42%–5.52%), p < .001; CD8: 20.35% (11.48%–31.25%) vs. 4.74% (3.46%–6.18%), p < .001] (Fig. 2A, B).

Percentages of CD4+ and CD8+ T cells expressing Caspase 3 from untreated HIV-1-infected patients, HAART patients, and healthy controls. Caspase 3 expression increased in both CD4+ and CD8+ T cells in HIV-1-infected patients
Similar results were observed in both memory CD4+ T cells and naive CD4+ T cells and memory CD8+ T cells and naive CD8+ T cells (Fig. 2C, D).
No correlations between CD4+ T cell counts, HIV-1 RNA levels, and percentages of Caspase-1+CD4+ T cells and Caspase-3+CD4+ T cells
Among the untreated HIV-1-infected patients, neither CD4+ T cell count nor HIV-1 RNA levels were correlated with the proportions of CD4+ T cells expressing Caspase 1 (Fig. 3A, B); however, the percentage of HLA-DR+CD38+CD8+ T cells was positively correlated with CD4+ T cells expressing Caspase 1 (Fig. 3C).
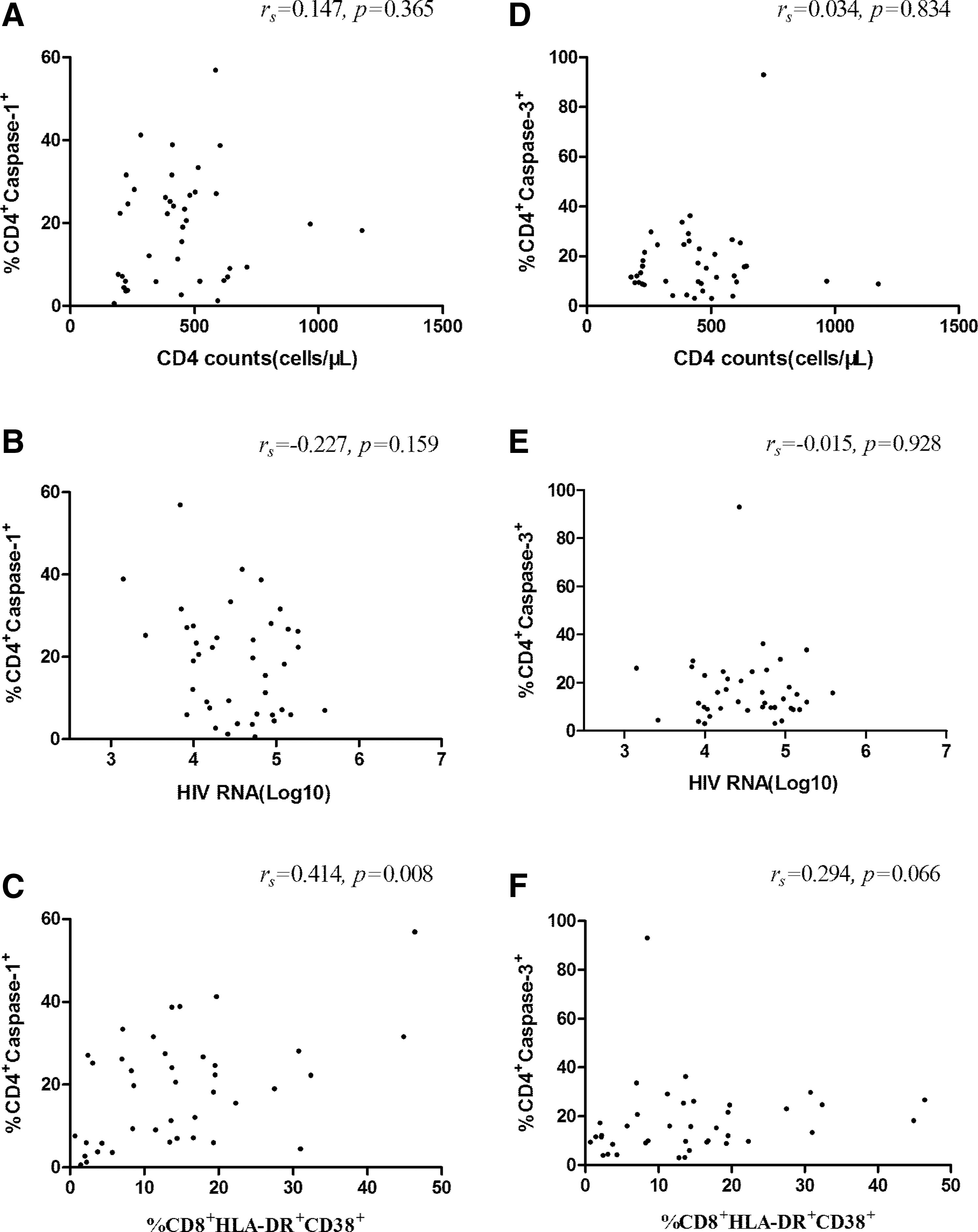
Correlations between CD4+ T cell counts, HIV-1 RNA levels, and percentages of HLA-DR+CD38+ CD8+ T cells, Caspase 1+CD4+ T cells, and Caspase 3+CD4+ T cells among the untreated HIV-1-infected patients. Correlations between percentages of Caspase 1+CD4+ T cells and CD4+ T cell counts
Meanwhile, among the untreated HIV-1-infected patients, neither CD4+ T cell counts nor HIV-1 RNA levels were correlated with the proportions of CD4+ T cells expressing Caspase 3 too (Fig. 3D, E). Additionally, percentages of HLA-DR+CD38+CD8+ T cells did not correlate with CD4+ T cells expressing Caspase 3 (Fig. 3F).
Caspase 1 expression is downregulated in CD4+ T cells in HIV-1-infected patients following HAART
The percentage of CD4+ T cells expressing Caspase 1 significantly decreased in the HAART group compared with untreated HIV-1-infected patients [6.05% (3.43%–9.78%) vs. 19.40% (6.36%–27.00%); p < .001]. Furthermore, the percentage of Caspase 1+ CD4+ T cells in the HAART group was comparable with that observed in healthy controls [6.05% (3.43%–9.78%) vs. 4.65% (3.85%–6.88%); p = .514] (Fig. 1A, B). In contrast to the percentage of CD4+ T cells expressing Caspase 1, the percentage of CD4+ T cells expressing Caspase 3 in the HAART group was comparable with that observed in untreated HIV-1-infected patients [9.67% (7.09%–19.80%) vs. 12.75% (9.11%–24.20%); p = .132] and was still higher than that observed in healthy controls [9.67% (7.09%–19.80%) vs. 4.18% (3.42%–5.52%); p < .001] (Fig. 2A, B).
The percentage of CD8+ T cells expressing Caspase 1 did not change significantly in the HAART group compared with untreated patients (Fig. 1A). While the percentage of CD8+ T cells expressing Caspase 3 declined significantly in the HAART group, it was still higher than the percentage observed in the healthy controls (Fig. 2A).
Furthermore, the percentages of memory and naive CD4+ T cells expressing Caspase 1 were significantly lower in the HAART group compared with untreated HIV-1-infected patients and were comparable with the percentages observed in healthy controls (Fig. 1C). However, the percentages of memory and naive CD4+ T cells expressing Caspase 3 in the HAART group were comparable with those observed in untreated HIV-1-infected patients and were still higher than those observed in healthy controls (Fig. 2C).
Moreover, the percentages of memory and naïve CD8+ T cells expressing both Caspase 1 and Caspase 3 were similar as those observed in all CD8+ T cells. The percentages of the two subgroups of CD8+ T cells expressing Caspase 1 did not change significantly in the HAART group compared with untreated HIV-1-infected patients(Fig. 1D), while the expression levels of Caspase 3 in each of the cell lines declined significantly in the HAART group, but still remained higher than levels in cells from healthy controls (Fig. 2D).
Increased expression of apoptosis-related markers in both CD4+ and CD8+ T cells despite HAART
Markers signifying early stages of the extrinsic and intrinsic apoptotic pathways—Caspase 8 and Caspase 9, respectively—in T cells changed inconsistently following HAART; however, they remained higher than those measured in healthy controls. The percentages of CD4+ T cells expressing Caspase 9, but not Caspase 8, decreased significantly in the HAART group, but were still higher than those observed in healthy controls. Similarly, the percentages of Fas+ and FasL+ CD4+ T cells remained elevated in the HAART group. The percentages of HLA-DR+CD38+ CD4+ T cells decreased in the HAART group, but remained higher than those of healthy controls. However, the percentage of CD8+HLA-DR+CD38+ T cells in the HAART group decreased to a level comparable with that of healthy controls. The percentages of Annexin V+7-AAD−CD4+ T cells in untreated HIV-1-infected patients were significantly higher than that observed in healthy controls; however, even though the percentage decreased in the HAART group, it still remained higher than that of healthy controls (Fig. 4.).
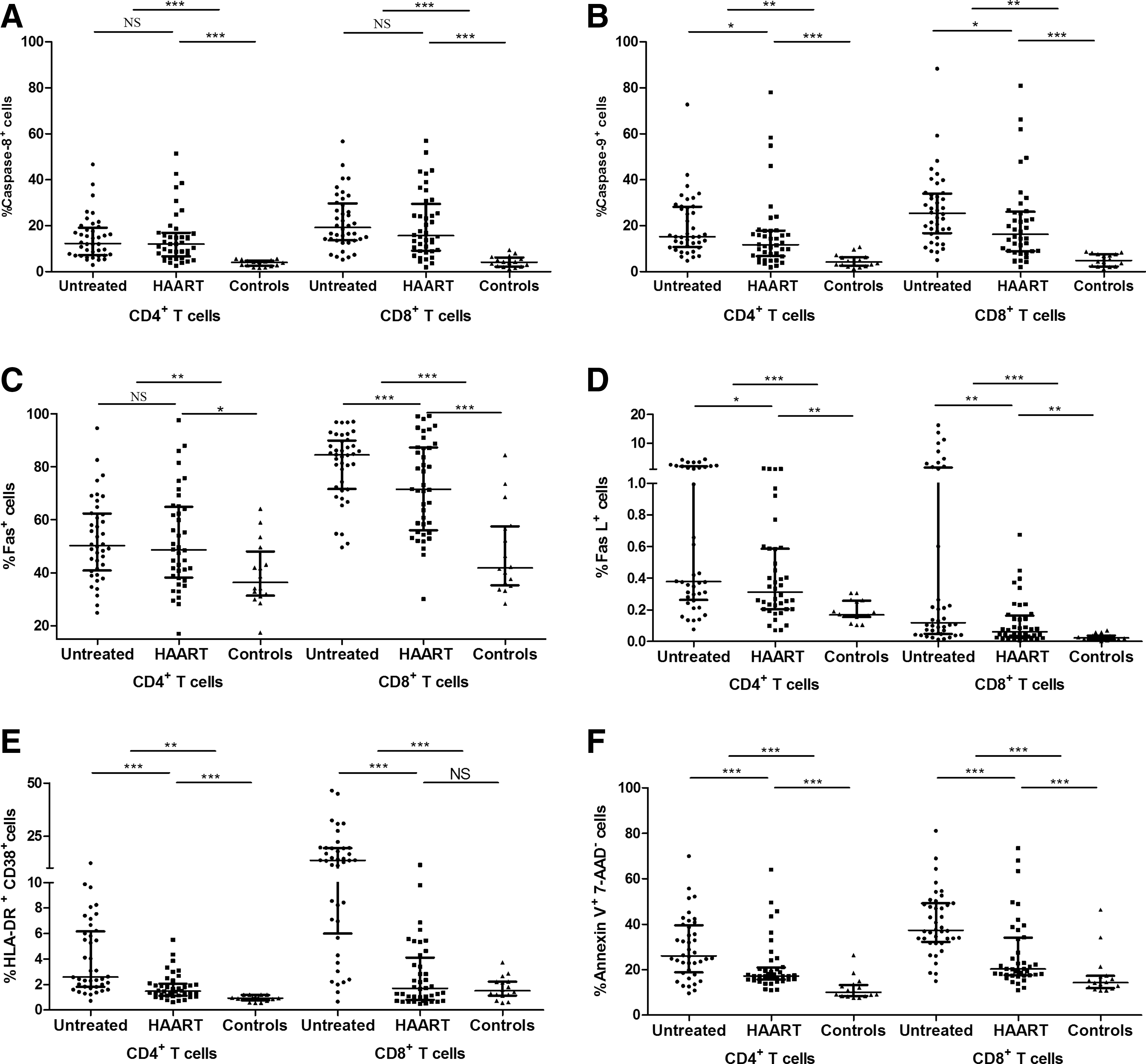
Increased expression of apoptosis-related markers, Caspase 8+
Discussion
Both pyroptosis and apoptosis were reported to be the main causes of CD4+ T cell depletion during HIV-1 infection. 1 –8 In this study, we reported the impact of HAART on the percentages of T cells undergoing pyroptosis and apoptosis in HIV-1-infected patients.
Pyroptosis is triggered by Caspase 1 following the initiation of specific cytosolic signals. 10,19 –22 In agreement with this, Doitsh et al. 8 reported that pyroptosis accounted for >95% of resting lymphoid CD4+ T cell loss. We confirmed that a high proportion of peripheral CD4+ T cells were undergoing pyroptosis, indicating that pyroptosis may be one of the main causes of CD4+ T cell depletion. Meanwhile, the percentages of CD8+ T cells expressing Caspase 1 did not increase in HIV-1-infected patients. One explanation might involve bystander CD4+ T cells being abortively infected by HIV-1, resulting in cytosolic accumulation of short HIV-1 DNA transcripts. 10 However, CD8+ T cells cannot be infected abortively or completely. This may further explain why CD4+ T cells were depleted, while the number of CD8+ T cells remained unchanged in HIV-1-infected patients. Interestingly, the CD4+ T cell counts were not correlated with the percentages of CD4+ T cells that express Caspase-1 in the current studies. The most likely explanation of this observation is that pyroptosis is only one of the mechanisms of CD4+ T cell destruction in HIV infection. Direct cytopathic effect of HIV-1 and apoptosis also cause CD4+ T cell death. Decreased thymic output and enhanced migration of CD4+ T cells to lymph nodes further account for the depletion of CD4+ T cell counts. 23 –25
In a previous study, Caspase 1 serum levels were negatively correlated with HIV-1 RNA levels. 26 A recent study showed that cell-to-cell transmission of HIV-1 was required to trigger pyroptotic death of lymphoid tissue-derived CD4+ T cells, while free HIV-1 particles failed to induce pyroptosis in vitro. 22 This may explain our result that the percentage of CD4+ T cells expressing Caspase 1 was not correlated with HIV-1 RNA levels. Munoz-Arias et al. reported that the majority of blood-derived CD4+ T cells from HIV-1-infected subjects, regardless of HAART, did not exhibit Caspase 1 activation. 27 In contrast, we observed that HAART inhibited pyroptosis based on the percentage of CD4+ T cells expressing Caspase 1 decreasing to levels comparable with that of healthy controls. Similar to our result, previous studies also showed elevated expression of Caspase 1 in CD4+ T cells both in vivo and in vitro. 28 –30 Therefore, HAART may not directly prevent pyroptotic death of CD4+ T cells by inhibiting HIV-1 replication, but rather by blocking upstream events involved in productive HIV-1 infection in other CD4+ T cells.
It was previously reported that pyroptosis links the two signature events in HIV-1 infection: CD4+ T cell depletion and chronic inflammation. 8,9 In our study, the percentages of both CD4+ T cells and CD8+ T cells that express Caspase 1 were positively correlated with levels of the activation markers, HLA-DR and CD38. Following HAART, Caspase 1 expression was inhibited, while immune activation remained high in CD4 T cells. This phenomenon may be due to immune activation being driven by other factors (e.g., microbial translocation) besides persistent HIV-1 infection. 31 –33
Apoptosis was linked to CD4+ T cell loss in HIV-1 infection both in vitro and in vivo. 1 –7,34 –36 Our study showed that both the intrinsic and extrinsic apoptotic pathways were activated in HIV-1-infected patients, which were similar to previous reports. 34,35,37 Therefore, both apoptosis and pyroptosis contributed to CD4+ T cell depletion in HIV-1-infected patients. Unlike pyroptosis, apoptotic activity, and specifically that associated with the extrinsic pathway, remained elevated after HAART according to our results. Furthermore, both Fas and FasL, which are triggers of the extrinsic apoptotic pathway, were also highly expressed in CD4+ T cells. This is consistent with previous findings reported from other laboratories. 35,38 –40 This result indicated that apoptosis, but not pyroptosis, may have impeded immune restoration after HAART. Therefore, methods to decrease apoptosis to restore CD4+ T cell counts in patients with impaired immune reconstruction require further investigation.
The main limitation of this study was that we did not investigate CD4+ T cells from different tissues, such as lymph nodes or spleens. Cell-to-cell transmission of HIV-1 was shown to be essential to initiate pyroptotic cell death. 22 Given that the cells in this study were obtained from peripheral blood where cell contact occurs less frequently compared with that observed in lymphoid tissue, the results from our study could not be expanded to T cells associated with lymph nodes or spleens. Additionally, the cross-sectional nature of this study also impedes causal inferences. Thus, further investigations are still needed.
In conclusion, our study found that the percentages of CD4+ T cells expressing both Caspase 1 and Caspase 3 increased in HIV-1-infected patients, while only the percentage of CD8+ T cells expressing Caspase 3 increased in these patients. Our results suggested that inhibition of Caspase 1, but not Caspase 3, induction of cell death pathways in CD4+ T cells could be mediated by effective antiretroviral therapy, which warrants further investigation.
Footnotes
Acknowledgments
The authors are thankful to all of the HIV-1-infected patients enrolled for this study. This work received grants from the Chinese National Natural Science Foundation (grant no. 81301420), the 863 project (SS2014AA021403), and the 12th five-year infectious disease research project (2012ZX10001-003 and 2012ZX09303013).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.