
Abstract
We have constructed a single chain fragment variable (scFv) phage display library from a simian immunodeficiency virus (SIV)-infected rhesus macaque that developed unusually high-titer neutralizing antibody responses against tier-3, neutralization-resistant SIVmac239. The library was screened using trimeric (gp140) and monomeric (gp120) forms of the SIVmac239 envelope (Env) glycoprotein. We also cloned variable-heavy and variable-light (VH-VL) antibody fragments from seven previously described rhesus macaque B-cell lines (BLCLs) that produce SIV gp120-specific monoclonal antibodies (mAbs). Thirty-two gp140-specific mAbs were selected along with 20 gp120-specific ones. gp120-specific mAbs were only from the VH4 family, while gp41-specific mAbs were primarily from VH1, followed by VH4 and VH3. Rhesus macaque BLCL-derived mAbs belonged primarily to the VH4 family of antibodies followed by VH3 and a smaller number of VH1s. A preferential VH combination with Vλ light chain was observed with phage display-selected SIV Env-specific mAbs (gp120 and gp140), but not with BLCL-derived antibodies or the unpanned library. None of the tested antibodies had detectable neutralizing activity against tier-3 SIVmac239. The majority of gp120-specifc mAbs potently neutralized tier-1 SIVmac316 with 50% inhibitory concentration (IC50) values below 1 μg/ml. For gp140-specific antibodies, which were all specific for the gp41-subunit, 2 out of 11 tested neutralized SIVmac316 (IC50 of 7 and 5 μg/ml, respectively). These data suggest an order of preferential VH segment usage for SIV-specific antibodies in rhesus macaques. These antibodies will be useful in assessing the contribution of non-neutralizing antibodies to inhibition of SIV infection in vitro and in vivo.
Introduction
T
Induction of broadly neutralizing antibodies (bnAbs) is probably the most desirable feature for development of an efficacious HIV vaccine. However, bnAbs are difficult to generate by vaccination despite the design of complex vaccine immunogens. The difficulties are partly due to the failure of immunogens to engage B cells that express the adequate germline receptors. 13 –16 The RV144 trial, the most recent phase III clinical trial, found no correlation of partial protection with neutralizing antibodies (nAbs), but instead found a correlation with non-neutralizing Env-binding antibodies that target the V1-V2 loop. 17 –21 Similarly, live-attenuated strains of SIVmac239 provide sterilizing protection in rhesus macaques, but analysis of antibody responses fails to detect potent nAbs in the protected animals. 22
Thus, the rare nature of bnAbs raises questions as to the role of these antibodies relative to the total Env-specific population of antibodies and whether efforts to induce responses that produce bnAbs is possible using current vaccine strategies. Understanding variable-heavy (VH) segment usage could also be important for the induction of protective non-neutralizing antibodies (nnAbs) such as those observed in the RV144 vaccine trial. 17 –21
The vast majority of HIV vaccine candidates are being tested in the SIV-macaque model of AIDS pathogenesis. Despite a very high degree of homology between human and rhesus macaque Ig genes, it is unclear whether similarities exist in VH segment usage following SIV infection or vaccination. Single cell-based methodologies using flow cytometry have allowed for comprehensive analyses of SIV/HIV-1 Env specific Ig gene repertoires. However, these techniques require antigen-specific cell sorting, which could result in unperceived biases due to the paucity of HIV/SIV Env proteins.
Construction of phage display libraries is not influenced by antigen specificity. Thereafter, the libraries can be screened indefinitely with whole antigen, antigen fragments, or targeted forms of antigens. One of the major disadvantages of phage display technology is the random combination of VH and variable-light (VL) chains, which do not represent the natural association of a VH and its in vivo associated VL. 23
In this study, we report the construction of a single chain fragment variable (scFv) phage display library from an SIV-infected macaque with potent neutralizing activity against the tier-3, neutralization resistant SIVmac239 isolate. 24 Ig gene families were analyzed following library panning onto oligomeric (gp140) and monomeric (gp120) forms of SIV Env glycoprotein. Analysis of the unpanned library and the rhesus macaques BLCLs revealed that the VH4 family is the most prominent among rhesus antibodies. The same observation was made for gp120-specifc antibodies, which were all VH4s. However, the exclusivity of VH4 usage for gp120-specific antibodies could be the result of suboptimal selection due to the paucity of SIV Env antigen.
In contrast, gp140-specific antibodies were highly diverse with antibodies belonging to VH1, VH3, and VH4 families. Unlike the unpanned library or rhesus macaques BLCLs, VH1 was the most abundant family among gp140-specific antibodies, followed by VH4 and VH3. Major differences in VH segment usage and neutralization potency were found between antibodies targeting the gp120 surface subunit or the gp41-ectodomain. Phage display is undoubtedly a useful method that can improve our knowledge of VH segment usage in experimental SIV infection of rhesus macaques.
Materials and Methods
Animal samples
Archived spleen biopsies were obtained from Mm333-95, a rhesus macaque infected with SIVmac239. 24 The AE637 plasma pool from SIV-infected macaques and plasma from SIVmac239Δnef immunized macaque Mm376-04 were previously described. 24,25
Construction of scFv library
Protocols for library construction and monoclonal antibody (mAb) selection were previously described. 26 Total RNA was prepared using QIAGEN's RNeasy Kit and following manufacturer's protocol (QIAGEN, Valencia, CA). Complementary DNA (cDNA) was synthesized using the SuperScript III First-Strand Synthesis Kit (Invitrogen, Carlsbad, CA) and used as template for polymerase chain reaction (PCR). Briefly, a scFv library with a long or short linker between variable fragment heavy chains (VH) and light chains (Vκ and Vλ) was constructed using two-step PCR amplifications. Table 1 presents PCR primers used for amplification of antibody variable regions. In the first round, VH genes were amplified by performing six reactions with VH primers for short linkers and six reactions with VH primers for long linkers. For both sets (short or long linkers), one reverse primer corresponding to human IgG isotypes was used. Sixteen Vκ and 27 Vλ (forward and reverse) primer combinations were used for amplification of VL fragments. Vκ and Vλ gene products were mixed together. In the second step, equimolar VH short or long linkers and Variable light chains (Vκ plus Vλ) were used to create overlap-extension PCR products for short or long linker scFvs. The resulting VH-VL products and the phagemid vector pComb3xSS were digested with Sfi1 (New England Biolabs, Ipswich, MA) and gel-purified before vector-insert ligation using T4 DNA ligase (New England Biolabs). Ligation DNA was ethanol-precipitated and purified using standard molecular biology techniques. Ligation DNA was then electroporated into XL1 Blue Escherichia coli using a Gene Pulser Xcell (Bio-Rad, Hercules, CA), and a phage library preparation was obtained after amplification in presence of VCSM13 helper phage. Library diversity was assessed by randomly picking 88 clones for DNA mini-preparation and DNA sequencing. These 88 clones picked from the unpanned library represent at least 1/50,000th of phage input for screening and selection of SIV+ clones. Briefly, 1 μl of diluted phage preparations (10−3 to 10−6) is used to infect 50 μl of bacterial cultures and plated onto Luria-Bertani broth (LB broth; AmericanBio, Natick, MA) agar plates supplemented with 50 μg/ml of carbenicillin (AmericanBio). Single colonies were picked and cultured for DNA mini-preparation and sequencing. Library screening (panning) for selection of SIV+ clones started with 3 × 50 μl of phage preparation.
In the first round, VH genes were amplified by performing six reactions with VH primers for short linkers and six reactions with VH primers for long linkers. For both sets (short or long linkers), one reverse primer corresponding to human IgG isotypes was used. Sixteen Vκ and 27 Vλ (forward and reverse) primer combinations were used for amplification of VL fragments. Vκ and Vλ gene products were mixed together. In the second step, equimolar VH short or long linkers and VL chains (Vκ plus Vλ) were used to create overlap extension PCR products for short or long linker scFvs.
PCR, polymerase chain reaction; scFv, single chain fragment variable; VH, variable-heavy; VL, variable-light.
Library panning and screening
Purification of the secreted monomeric (gp120) and oligomeric (gp140) forms of SIV Env was previously described. 25 Enzyme-linked immunosorbent assay (ELISA) plates were coated with 100 ng of antigen (gp140 or gp120) per well. Four rounds of panning were initially performed onto gp140. Two rounds of panning onto gp120 were later performed because no gp120-specific antibodies were detected after gp140 panning. The number of pannings was limited to two, to maintain diversity and avoid clone redundancy (as observed with gp140). ELISA plates were coated with 100 ng of gp140 or gp120 for identification of Env-specific scFvs present in the supernatant of Isopropyl β-D-1 thiogalactopyranoside (IPTG) induced bacterial cultures. Each supernatant was also tested on bovine serum albumin (BSA) as a negative control (100 ng/well).
Antibody fragment cloning from rhesus macaque B cell lines
Immortalized rhesus macaque B cell lines (BLCLs) producing mAbs targeting SIV Env glycoprotein were previously described. 8,27,28 Seven of these BLCLs, expressing the SIV Env-specific rhesus monocolonal antibodies, were used in this study: 1.10A, C26, 1.11A, 5.5B, 3.11H, 3.4E, and 3.5F. 8,27,28
Total RNA was prepared using QIAGEN's RNeasy Kit (Qiagen). cDNA was synthesized using the SuperScript III First-Strand Synthesis Kit (Invitrogen) and used as template for the PCR reaction. Antibody VH-VL fragments were amplified using the same primers as for the phage display library (Table 1). To obtain VH fragments, a pool of all forward VH primers (short linker) was used in combination with the corresponding reverse primer. To obtain Vκ or Vλ fragments, a pool of all forward Vκ or Vλ forward primers was used in combination with a pool of the corresponding reverse primers. All BLCLs were tested with both sets of Vκ and Vλ primers. In the second step of PCR the VH and VL obtained from each BLCL were linked together and cloned into pComb3XSS, as described for the phage library.
scFv sequence analysis
DNA mini-preparation was performed for library or antigen-specific colonies following protocol from the manufacturer (QIAGEN, Valencia, CA). scFv sequences were determined by Sanger sequencing using primers Ompseq (5′-aagacagctatcgcgattgcag-3′) and HRML-F (5′-ggtggttcctctagatcttcc-3′). VH and VL fragments were trimmed using the DNA analysis software Geneious (Auckland, New Zealand) and analyzed using IMGT/V-QUEST in the International Immunogenetics Database (
Somatic hypermutation frequencies were determined using nucleotide identity with the closest germline obtained with IgBlast tool from the National Center for Biotechnology Information (NCBI). IgBlast tool was used for both heavy (VH) and light (VL) chains except for VL with V gene IGKV3-8*-01 P for which closest germline was found using IMGT/V-QUEST. Clonality was determined with the criteria that antibodies from a clone had the same V and J allele assignments, the same CDRH3 length, and ≥80% CDRH3 identity.
Conversion of scFv into scFv-Fc format
Two gp120-specific scFvs and 1 gp140-specific scFv were converted into scFv-Fc format corresponding to human IgG1 isotype as previously described.
29
scFv nucleotide sequences were codon-optimized, and synthetic genes (
ELISA and immunoblotting
ELISA was performed as previously described. 25 ELISA plates were coated at 100 ng of antigen per well. An horseradish peroxidase (HRP)-conjugated anti-HA mAb was used for detection of scFvs (Thermo Fisher Scientific, Waltham, MA). An HRP-conjugated anti-human IgG (Santa Cruz Biotechnology, Santa Cruz, CA) was used for detection of plasma antibodies or scFv-Fc fusion proteins. TMB substrate and TMB stop solution (Bio-Rad) were used to detect bound HRP-conjugated antibodies. Primary antibody (scFvs, scFv-Fcs, or plasma antibodies) binding was determined by acquisition of absorption on a VICTOR plate reader (PerkinElmer, Waltham, MA) after HRP reaction at 450 nm.
Immunoblotting was performed as previously described 25 using the same primary and secondary antibodies as for the ELISA.
SIV neutralization assay
Virus neutralization was performed by SEAP assay as previously described. 3,4,7,25 In this study, we used 5,000 C8166-SEAP cells per well. Cells were infected with either 1 ng of tier-3 resistant strain SIVmac239, 1 ng of tier-2 moderately resistant strain SIV251-MT4, 3 or 5 ng of tier-1 sensitive strain SIVmac316. 9
Results
We previously described a rhesus macaque (Mm333-95) with unusually high-titer nAbs against SIVmac239. To examine the antibody specificities generated in this animal, an scFv phage display library was constructed using the primers presented in Table 1 and following previously described protocols. 26 Eighty-eight clones were randomly picked from the library and sent out for DNA sequencing (Eton Biosciences, Boston, MA). Only sequences with intact reading frames (i.e., no stop codon) encoding VH or VL chain were included in the analysis (Figs. 1 and 2). As expected we found usage of several VH families, including VH1-VH4 and VH7. Twenty-nine of the 57 VH clones obtained were from VH4. Of the remaining clones, 18 were VH3, 7 were VH1, 2 were VH2, and 1 was VH7 (Fig. 1a). Among the 46 VL clones, 22 were Vλ and 24 were Vκ (Fig. 1b).
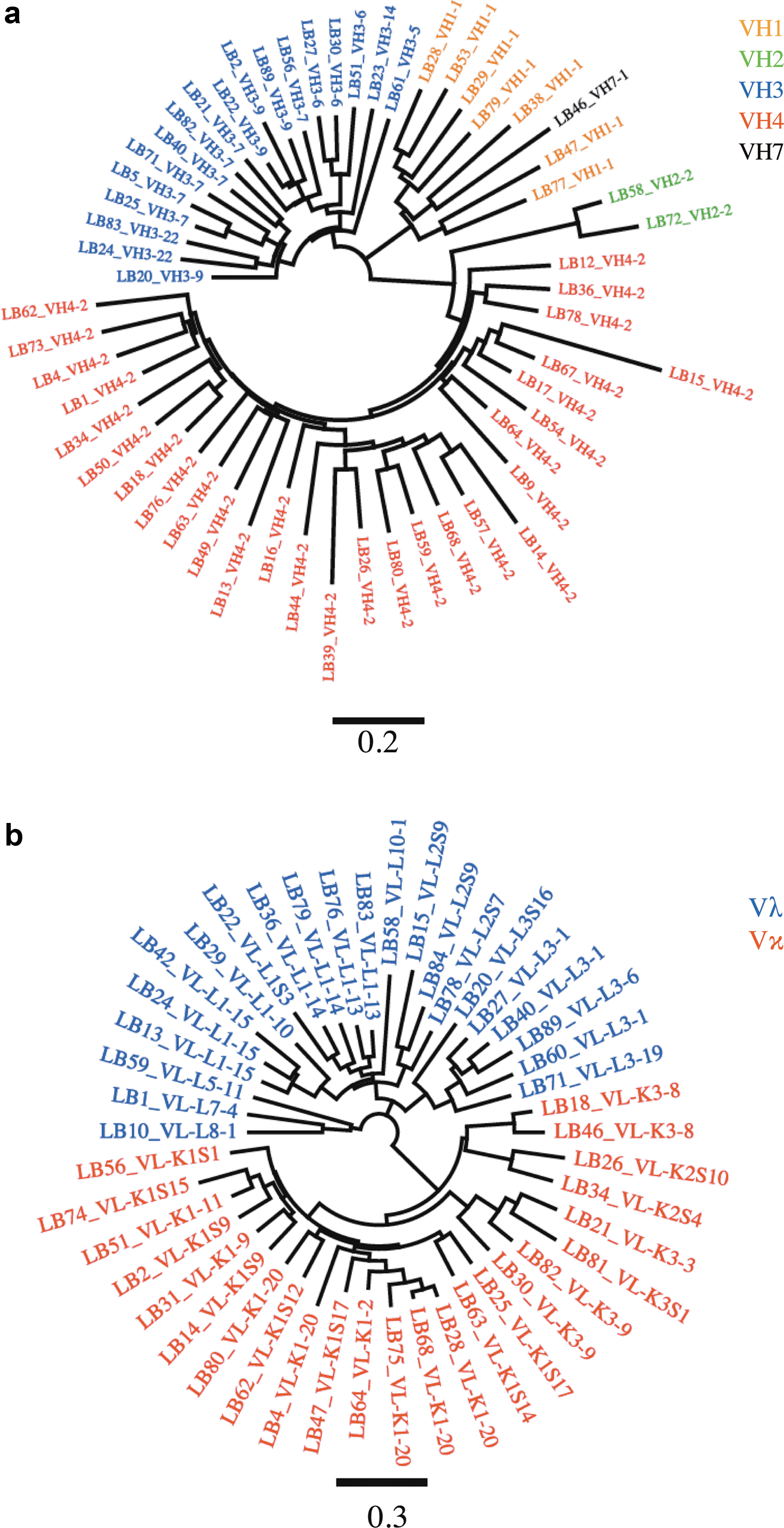
Diversity observed with antibody obtained from the unpanned library. Antibody nucleotide sequences corresponding to variable region from
Random pairing of VH and VL antibody fragments from the unpanned library 32 scFvs was analyzed. All antibody VH fragments were randomly combined with VL chain fragments (Vκ or Vλ). Sequences were analyzed using Prism. Ig germline family was determined using IMGT/V-Quest. VH, variable-heavy; VL, variable-light.
In addition, we produced 32 scFv clones (VH+VL combination) of which 16 were VH4, 11 were VH3, 2 were VH1, 1 was VH2, and 1 was VH7 (Fig. 2). As expected for these scFvs, VHs randomly combined with Vλ or Vκ. Out of the 32 VLs (17 Vλ and 15 Vκ), 2 out of 3 were Vλ in combination with VH1 segments, 1 Vκ combined with the only antibodies with VH2 segments, 7 out of 13 Vλ combined with VH3 segments, 7 out of 16 Vλ combined with VH4, and 1 Vκ combined with the only VH7 (Fig. 2). Sequence alignment revealed that the library was highly diverse, and no redundancy was observed for any of the identified functional VH or VL.
The library was subsequently panned using SIVmac239 gp140 as a selection antigen for four rounds. Forty-six SIV Env-specific clones were selected of which 32 were unique. Eighteen clones were selected for further analysis based on VH family relationship and CDR-H3 similarities. All clones tested by ELISA bound to gp140. We then distinguished specificity to gp120 or gp41 by immunoblot and found all tested clones were specific to the ectodomain of gp41. Because no gp120-specific clones were identified after panning onto gp140, we next panned the original library on monomeric gp120. We limited panning to two rounds to avoid clone redundancy (as was observed with gp140 panning round No. 4) and we selected 20 anti-gp120 scFvs, which bound to gp120 and gp140 but not BSA.
Despite limiting panning on gp120 to two rounds, comparing the antibody gene sequences revealed more diversity among gp41-specific mAbs than gp120-specific antibodies (Fig. 3). All 20 gp120-specific scFvs were derived from the VH4 family alleles and were associated exclusively with Vλ light chains. In contrast, the 32 gp41-specific mAbs had a broader distribution, including 19 from VH1, 10 from VH4, and 3 from VH3. gp41-specific scFvs also combined with 10 Vκ chains and 22 Vλ chains. These data reflect a higher presence of Vλ light chain among selected SIV Env-specific antibodies, while a more balanced VH-Vκ versus VH-Vλ association was observed with the unpanned library. A noticeable difference with the unpanned library and rhesus BLCLs is that VH1 was the most abundant family among the gp41-specific antibodies. Table 2 presents a summary of the characteristics of phage display selected antibodies. Antibodies specific to gp41 and gp120 could be assigned to 12 and 7 clonal groups, respectively.
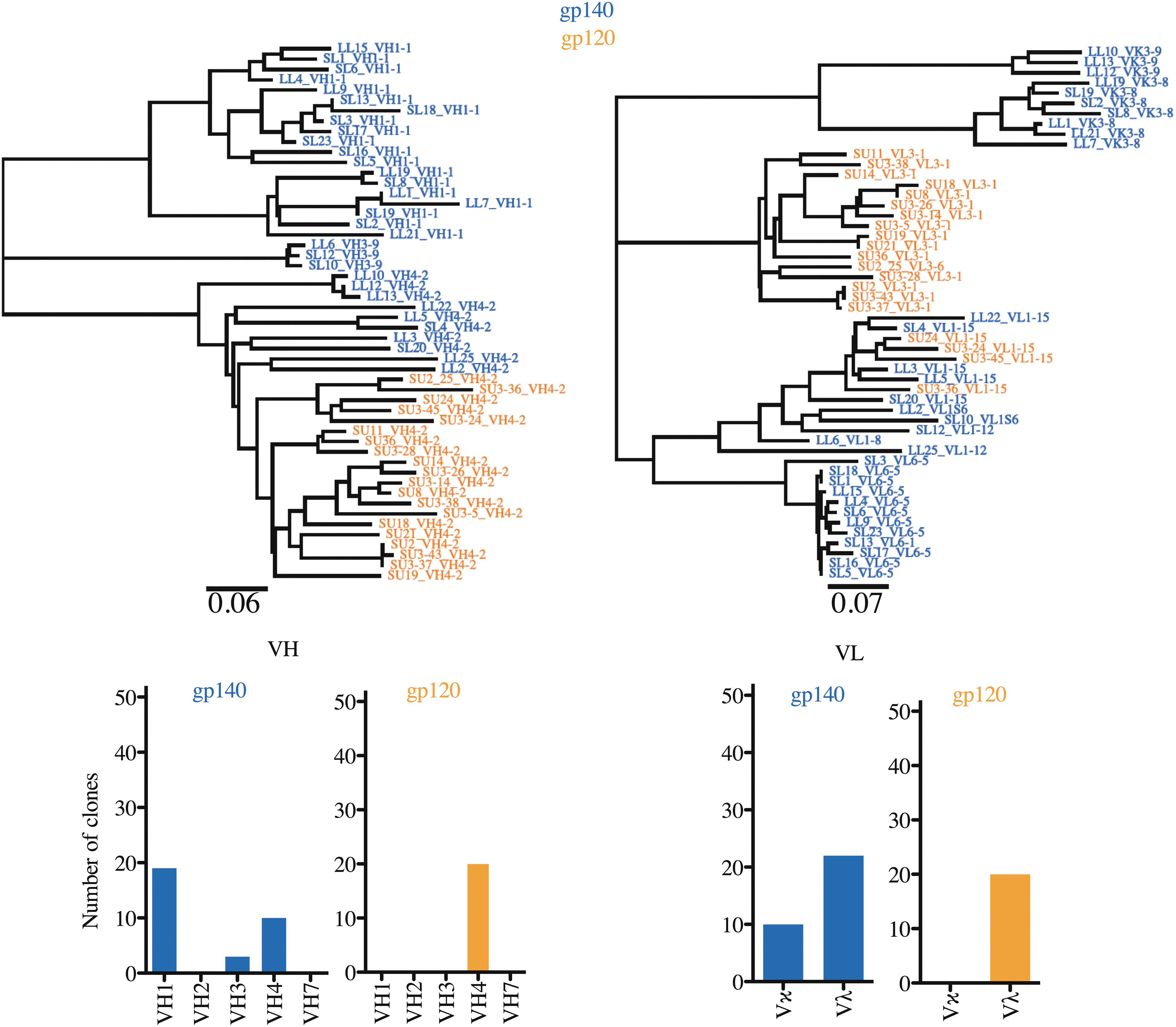
Phylogenetic relationship of SIV-specific antibodies. SIV-specific antibodies were selected after panning onto gp140 trimeric or gp120 monomeric forms of the SIV Env glycoprotein. Antibody nucleotide sequences were analyzed using IMGT/V-Quest. Numbers of clones were summarized for each major VH family and each type of VL (Vκ or Vλ). Env, envelope; SIV, simian immunodeficiency virus.
Heavy and light chain V genes and complementary determining region (CDRH3 and CDRL3) were determined using IMGT/V-QUEST. Somatic hypermutation frequencies [SHM (%)] represent the nucleotide sequence divergence with closest sequence obtained using IgBlast tool from the National Center for Biotechnology Information (NCBI). scFv binding specificity was determined by ELISA using soluble SIVmac239 gp120 and gp140. IgBlast tool was used for both heavy (VH) and light (VL) chains except for VL with V gene IGKV3-8*-01 P (shaded box) for which closest germline was found using IMGT/V-QUEST. Clonality was determined with the criteria that antibodies from a clone had the same V and J allele assignments, the same CDRH3 length, and ≥80% CDRH3 identity.
ELISA, enzyme-linked immunosorbent assay; SHM, somatic hypermutation.
No gp120-specific antibodies were selected by panning onto gp140. There are at least two possible explanations: first, the gp41 ectodomain may be highly exposed in the context of the gp140 antigen, or second, gp41-specific antibodies were very abundantly produced in animal Mm333-95. Therefore, Mm333-95 plasma was tested for comparison of gp120- and gp41-directed antibody levels. Mm333-95 plasma was compared to control plasma consisting of pooled plasma from SIV-infected macaques and Mm376-04, an SIVmac239Δnef immunized macaque. 25 These plasmas were tested for binding to SIVmac239 gp120 and gp140 (Fig. 4). ELISA revealed that Mm333-95 plasma and pooled plasma contained more gp140-binding antibodies than gp120-binding antibodies (Fig. 4a). gp140 Binding antibodies appeared to be more abundant in Mm333-95 plasma in comparison with the pooled SIV+ plasma. On the contrary, the SIVmac239Δnef infected macaque Mm376-04 contained an equal ratio of gp120- and gp140-binding antibodies. Similar results were obtained by immunoblotting, where Mm333-95 plasma produced a band corresponding to gp41-ectodomain binding that was more intense than for plasma from SIVmac239Δnef-immunized animal Mm376-04 or pooled plasma from SIV+ macaques (Fig. 4b).
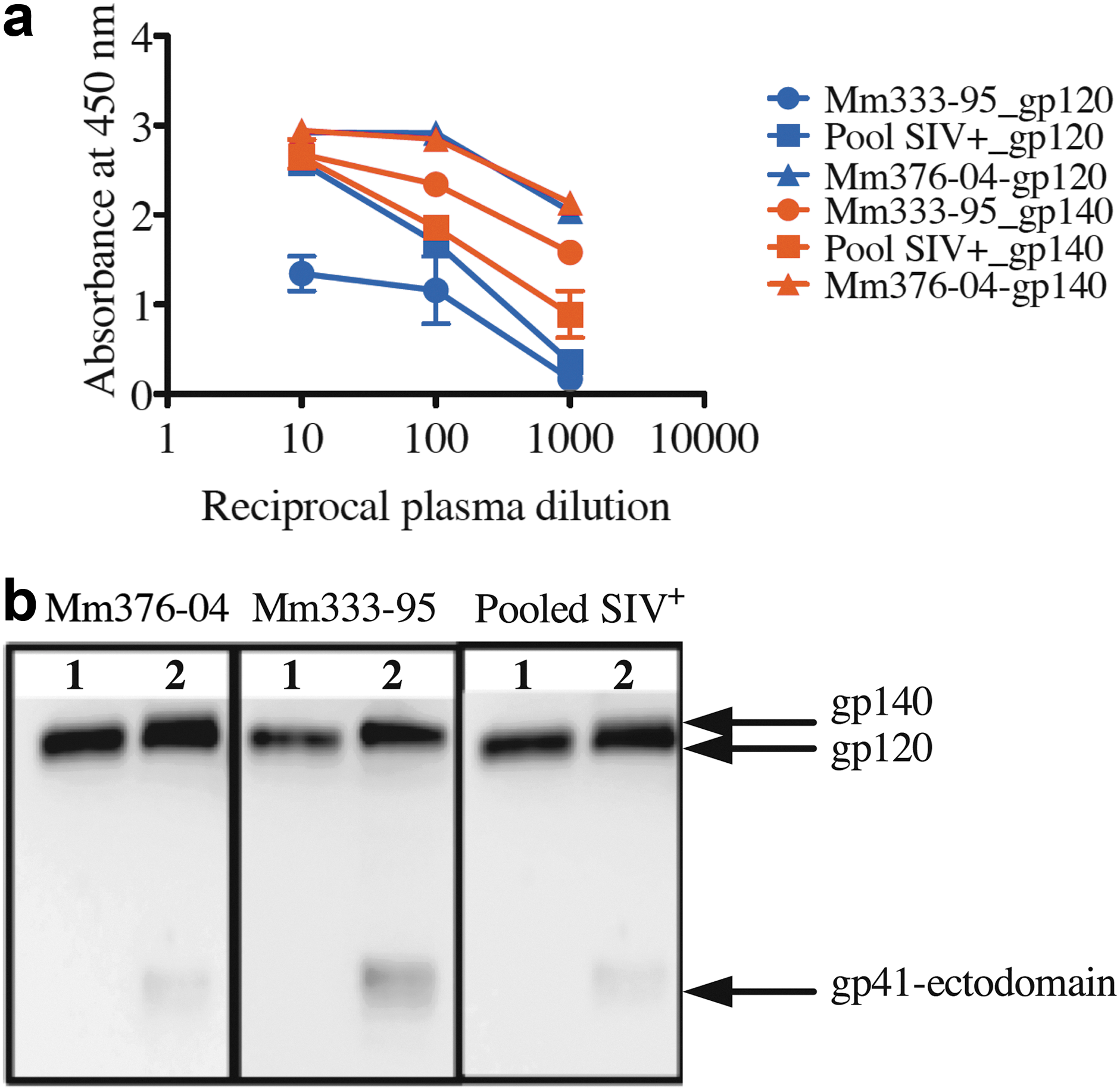
Characterization of anti-SIV mAbs and Mm333-95 plasma. Mm333-95 plasma was assayed for levels of anti-gp120, gp140 binding along with control plasma from SIV-infected macaques (Pooled SIV+), and from a SIVmac239Δnef immunized macaque (Mm376-04). ELISA plates were coated with 100 ng of SIVmac239 gp120 or gp140. For immunoblotting, gp120 and gp140 were separated on 12% SDS gels.
Purified scFv proteins were evaluated for neutralizing activity against SIVmac239, a neutralization resistant tier-3 strain, and SIVmac316, a neutralization sensitive tier-1 strain. No detectable neutralization of SIVmac239 was observed with gp41 or gp120-specific antibodies (data not presented). Two of the gp41-specific scFv neutralized SIVmac316 with a 50% inhibition concentration (IC50) of 5 and 7 μg/ml, respectively (Fig. 5a). SL5 and SL13 were closely related and both from the VH1 family. The CDR-H3 sequence of SL5 was PGDYSGSFDF, and the CDR-H3 sequence of SL13 was PGDYTGAFDL. All nine tested gp120-specific scFvs potently neutralized SIVmac316 with an IC50 below 1 μg/ml. Only SU2 and SU8 had an IC50 of 2.5 μg/ml (Fig. 5a).

SIVmac neutralization by selected mAbs. Representative mAbs were tested for neutralization of the tier-3 resistant strain SIVmac239, tier-2 moderately resistant SIVmac251, and the sensitive strain SIVmac316.
Because virus neutralization by scFvs may lack additional avidity effects found in dimeric forms of antibody, we selected two gp120-specific (SU24 and SU3.43) and one gp41-specific (SL5) scFvs for conversion into the scFv-Fc format. Gene sequences corresponding to SU24, SU3.43, and SL5 were codon-optimized for expression in mammalian cells. The specificity of the three scFv-Fc proteins was validated by immunoblotting (Fig. 5b). We then tested the three scFv-Fc proteins for neutralizing activity against SIV (Fig. 5c). None of the antibodies had detectable neutralization activity against SIVmac239, a neutralization resistant tier-3 virus. gp120-specific antibodies SU24-Fc and SU3.43-Fc potently neutralized tier-2 (SIVmac251-MT4) and tier-1 (SIVmac316) viruses with IC50 values below 1 μg/ml (Fig. 5c). SL5-Fc had no detectable activity against SIVmac239, SIVmac251, or SIVmac316 (Fig. 5c).
These results are similar to those obtained with scFv forms of the gp120-specific antibodies, suggesting that there was little to no gain in potency due to addition of the Fc domain. In the case of SL5-Fc, it is unclear what may have caused the slight loss of neutralization potency, which did not decrease below the 50% infectivity with SIVmac316. It is possible that the larger size of the scFv-Fc format is limiting access to the gp41-ectodomain. More analysis will be required to determine whether gp41-specific antibody binding could be improved by combination with gp120 binding.
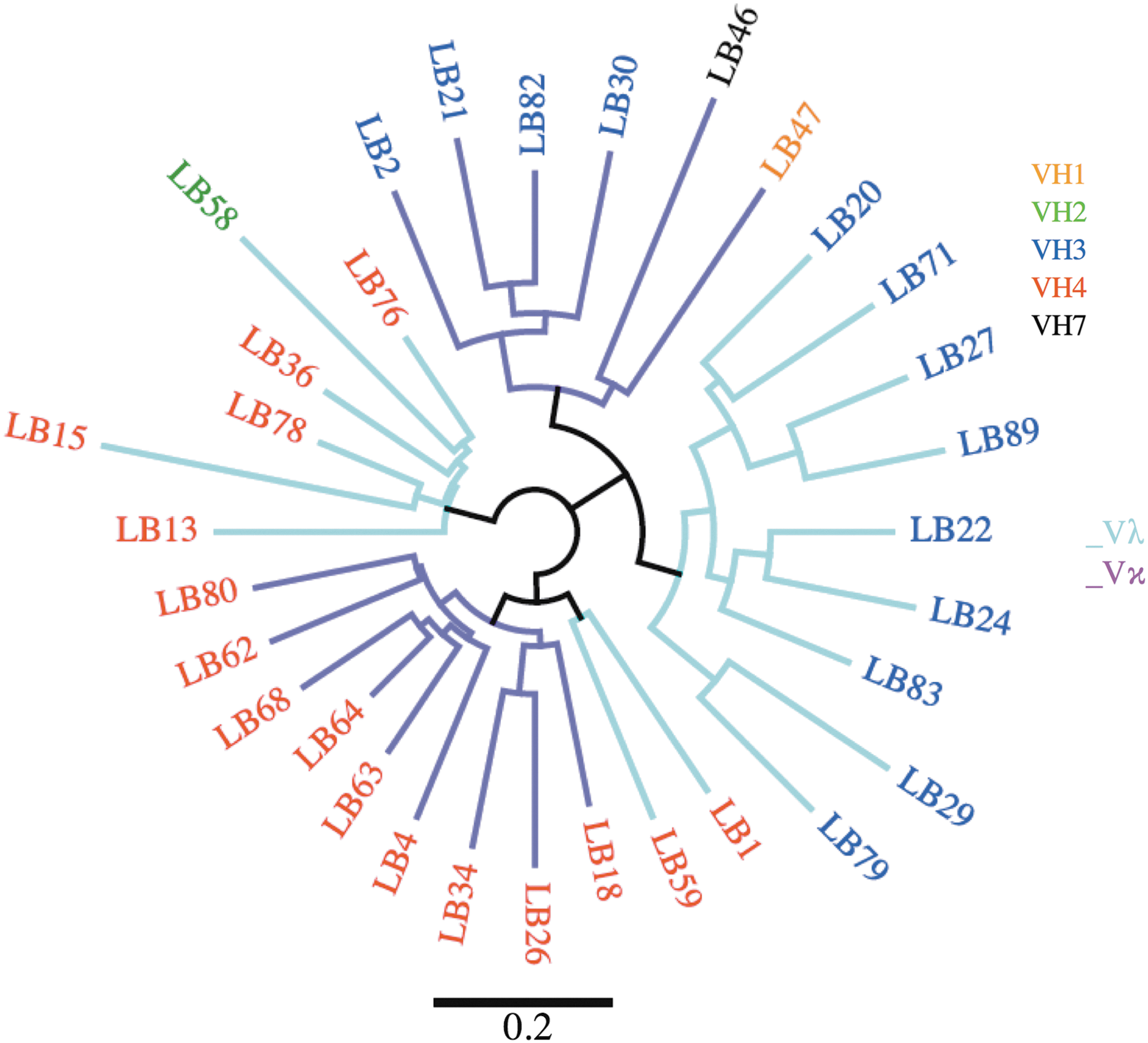
To assess the validity of phage display based analysis of Ig gene repertoire, the antibody sequences of seven SIV gp120-specific mAbs were cloned from rhesus macaque BLCLs (Fig. 6).
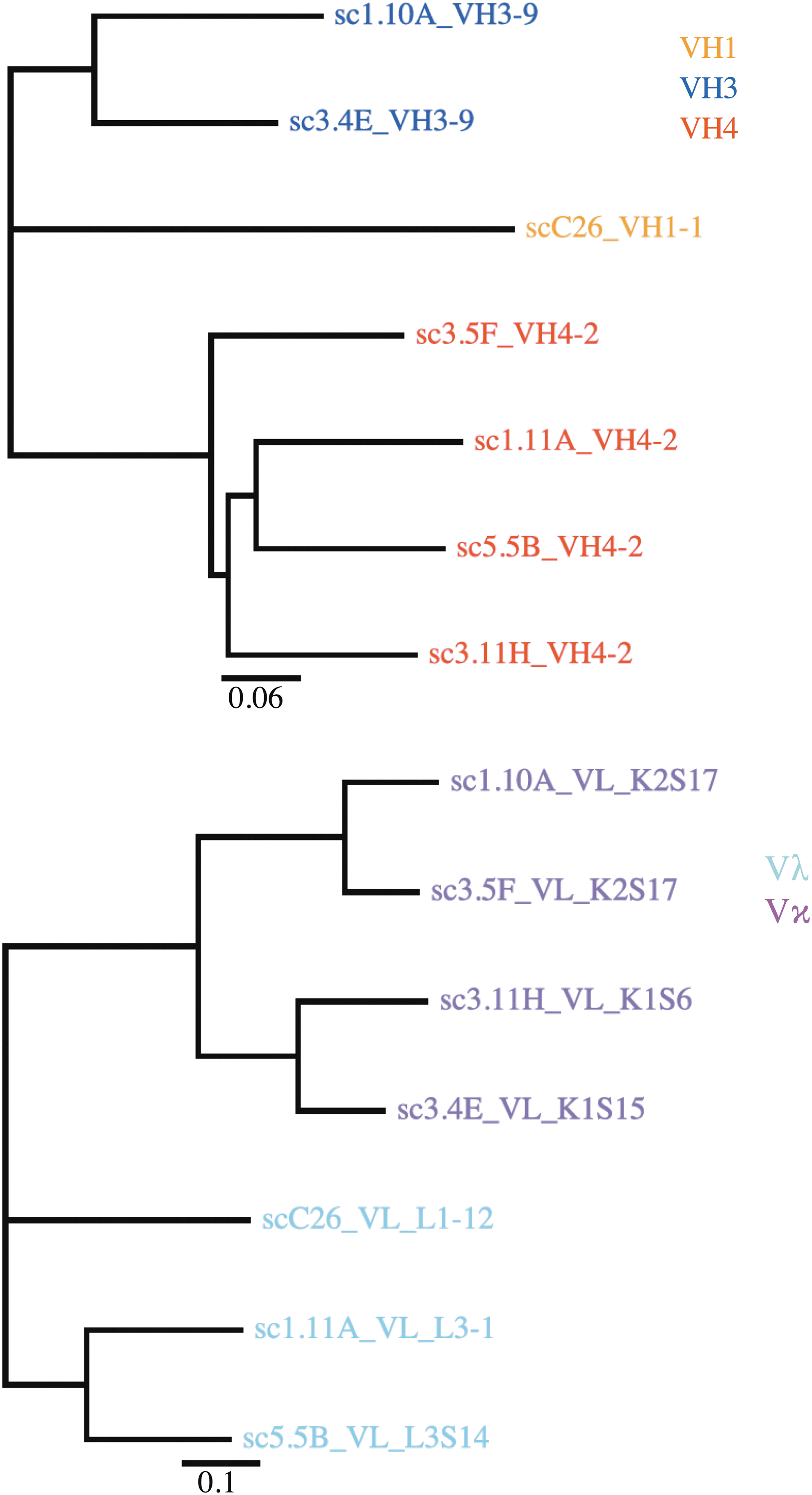
Phylogenetic relationship of mAbs from immortalized rhesus macaque B cells. Antibody variable fragments were obtained by PCR using VH and VL specific primers. Nucleotide sequences were analyzed using IMGT/V-Quest. Numbers of clones were summarized for each major VH family and each type of VL (Vκ or Vλ). PCR, polymerase chain reaction.
Four of the antibodies were derived from VH4, two from VH3 and one from VH1. This VH family representation was similar to that of the unpanned library. We found that random VH-VL combinations were observed, with four Vκ and 3 Vλ VL chains in contrast to the phage display library. The clonal antibody gene sequences of the BLCL belonged to the VH4, VH3, and VH1 families similar to the panel of antibodies produced by panning on gp140. We also observed a balanced use of light chains [four Vκ (4) and three Vλ] for BLCLs, similar to the unpanned library, but in contrast to the phage display-selected mAbs, altogether suggesting the existence of a preference of VL during phage display selection on SIV antigen.
Discussion
Using phage display selection, we have identified an order of VH family abundance (VH4 > VH3 > VH1) among SIV-Env specific mAbs. A similar VH family representation was observed with anti-SIV mAbs from immortalized rhesus macaque BLCLs. In contrast to BLCLs or the unpanned library, VH1 was the largest group among the SIV Env gp41-specific mAbs. In a previous study using phage display, all gp120-specific mAbs isolated from an SIVsmH635FC-infected rhesus macaque were reported to be VH3 and all gp41-specific mAbs were VH4s. 30
Furthermore, the selected gp120-specific mAbs potently neutralized multiple SIV strains, while the gp41-specific mAbs had no detectable activity. These results suggest the existence of Ig gene usage differences between gp120 and gp41 mAbs within a given rhesus macaque. In general, anti-gp120 mAbs appear to have a greater potential for SIV neutralization; however, it is difficult to determine whether these differences are the result of a suboptimal antibody selection due to differences in exposure of antibody binding sites across the SIV Env glycoprotein.
The SIVmac239-infected macaque Mm333-95 used in this study also displayed a higher concentration of gp41-specific antibodies in the plasma at week 42 postinfection when high-titer neutralization was detectable. 24 The high content of gp41-specific antibodies in Mm333-95 is intriguing because selected gp120-specific mAbs displayed more potential for tier-2 and tier-3 SIVmac neutralization. None of the tested mAbs had neutralizing activity against SIVmac239 that recapitulated the potent neutralizing activity of the plasma isolated from this animal. This raises the possibility that several mAbs may be required for potent neutralization. However, we have so far been unable to prove that possibility by testing different combinations of selected mAbs (data not presented). The lack of antibody capable of neutralizing SIVmac239 could also be due to biases in antibody VH and VL fragment combination. In this study, we showed that Mm376-04, an SIVmac239Δnef immunized animal, showed similar levels of SIVgp120- and gp41-specific antibodies in the plasma. We previously detected higher levels of SIV Env gp120-specific peripheral blood B cells from Mm376-04. 25
Identification of antigen-specific memory B cells from peripheral blood may be influenced by the choice of antigen. Design of multiple forms of antigens should be considered for B cell sorting by flow cytometry–based methods to avoid the loss of valuable clones. For example, the use of competitive probe binding and scaffolded probes has allowed the targeted isolation of SIV-Env specific mAbs by flow sorting. 31 The latter study successfully cloned mAbs against major site of SIV vulnerability, including nAbs, but also some nnAbs. This flow sorting methodology also revealed that the majority of SIV gp120-specific antibodies were from the VH4 family, with few from the VH3 family and far less from VH7 or VH1 families. However, it is unlikely that such a targeted approach would allow the cataloging of nnAbs for the maximum coverage of SIV Env glycoprotein.
Interestingly, phage display technology offers the possibility of screening the same library with an unlimited number of antigen scaffolds. Nonetheless, VH-VL pairing biases observed with phage display-selected mAbs should be given special attention. In summary, we believe that understanding of Ig fragment usage will shed light on the induction of protective anti-HIV/SIV antibody responses regardless of whether they should be neutralizing or not.
Sequence Data
All nucleotide sequences were submitted to the GenBank and are under accession numbers KU641040-KU641091 and KX785386-KX785463.
Footnotes
Acknowledgments
The authors thank Laura Hall for reagent ordering. The authors thank Dr. Christoph Rader for kindly providing the scFv fusion vector (Scripps Research Institute, Jupiter, FL). The authors also thank Dr. Shuji Sato and Dr. Andrea Kirmaier for useful comments about the article. This study was supported, in part, by a Boston College fund.
Author Disclosure Statement
No competing financial interests exist.