
Abstract
This study aimed to explore the contribution of high-mannose glycans in the masking of conserved V3 crown (GPG) and V2i epitopes on the hypervariable loops of most exposed distal surface of HIV-1 Env. Using lectins specific to Manα1-2Man residue containing Man6-9GlcNAc2 glycans extensively decorating HIV-1 Env, we found that Manα1-2Man-binding lectins enhance the exposure of these partially and transiently exposed epitopes and consequentially increase the neutralization strength of antibodies against these epitopes.
T
Despite being hypervariable, the V3 and V1/V2 loops present certain conserved features to perform certain well-defined functions in the virus, such as CD4 engagement, coreceptor interaction, viral tropism determination, and infectivity. 4 A well-defined neutralizing domain characterized within the tip of V3 and V1/V2 loops is the target of antibodies that are made by essentially all HIV-infected individuals and are easily induced by most candidate HIV vaccines. 5 These epitopes display conformational heterogeneity and are known to be dependent upon the native conformation of Env and remain occluded within the unliganded trimeric Env spike. 6
Recent structural data suggest that these epitopes remain extensively covered with N-linked glycans on and adjacent to V1/V2 and V3 loops on the native Env, and contribute to the conformational heterogeneity of these epitopes. The predominant type, ∼98% of glycans on native Env, is immature unprocessed Man5-9GlcNAc2 type N-glycans with fully processed mature GlcNAc2Man3GlcNAc2 and the GlcNAcMan5GlcNAc2-derived N-glycans representing only minor population. It has also been shown that Manα1-2Man terminating glycans (Man6-9GlcNAc2) are threefold more abundant on native Env than the monomeric gp120. 1,6 Taking these facts into consideration, we designed a study to elucidate the biological significance of the masking of V3 crown and V2i epitopes by high-mannose glycans using Manα1-2Man-binding lectins. 7,8
We generated pseudoviruses (PsVs) by cotransfecting the HEK 293T cells with a viral plasmid containing env gene and Δenv backbone plasmids pNL4-3Δenv or pSG3Δenv. The PsVs were used to infect TZM-bl reporter cells at 100 to 200 tissue culture infectious dose (TCID50) in the presence of di-ethyl amino-ethyl (DEAE)-dextran (10 μg/ml) for 48 h. The lectins were added to the viruses at a fixed concentration (IC50) and incubated for 30 min at 37°C. For neutralization experiments, serial dilutions of mAbs were added to the virus–lectin mixture and incubated for 1 h at 37°C followed by adding the TZM.bl cells along with DEAE-dextran. After 48 h, the virus neutralization was measured using a β-galactosidase-based assay.
We tested three anti-HIV lectins: griffithsin (GRFT), cyanovirin-N (CV-N), and scytovirin (SV-N), which recognize the Manα1-2Man-containing residues present on the terminals of D1, D2, and D3 arms of the Man6-9GlcNAc2 (Table 1). All the lectins inhibited the viruses, Figure 1a, GRFT being most potent followed by CV-N and SV-N, with median IC50 values 0.003, 0.039, and 0.356 μg/ml, respectively. The inhibitory potential of lectins correlated with their avidity, that is, number of glycan motifs recognized on the high-mannose structure. Subsequently, we estimated the neutralization sensitivity of the viruses to two representative antibodies against V2i (LDI/V) and V3(GPG) epitopes. We found that all viruses were more sensitive to V3 mAb2557 with median IC50 of 1.25 μg/ml, whereas V2i mAb1393Awas relatively less efficient in neutralizing the viruses, median IC50 24.86 μg/ml. These findings are in agreement with the published literature that V3 antibodies display more potency and breadth than V2i antibodies, Figure 1b. 9,10
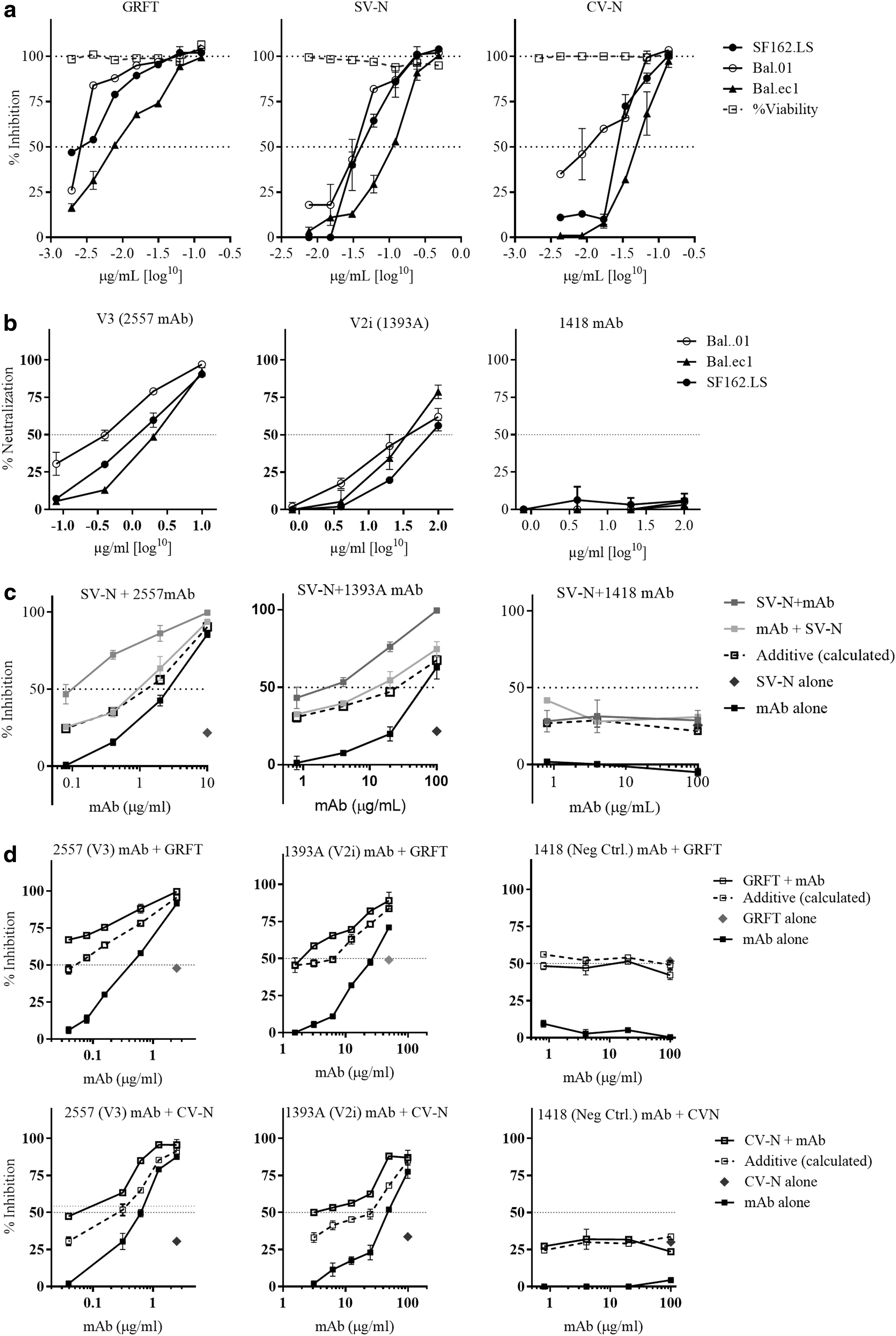
Inhibitory properties of Manα1-2Man-binding lectins and exposure of glycan-shielded loops.
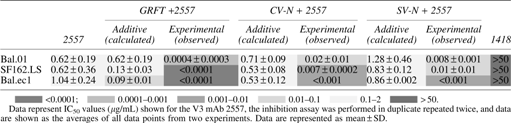
IC50 values (in μg/ml) are shown for the three viruses tested, the inhibition assay was performed in duplicate, and data are shown as the averages of all data points from two experiments. Data are represented as mean + SD.
While determining the effect of Manα1-2Man-binding lectins on the neutralization sensitivity of viruses to antibodies, 11 it was observed that in the presence of lectins, the neutralization capacity of all the antibodies displayed a highly significant increase, which was many fold higher than the predicted additive values (Fig. 1c, d). Such results were obtained only when viruses were incubated with lectins before the addition of mAbs. No enhancement in the neutralization potency was observed when viruses were incubated with mAbs followed by addition of lectins (Fig. 1c).
The fold change difference in the neutralization potency of 2557 against

Since it is well established that V3 and V1/V2 epitopes displaying conformational heterogeneity with Man6-9GlcNAc2 types of glycans abundantly present on the HIV-1 Env with terminal D1, D2, and D3 branches ending with Manα1-2Man remain occluded and display partial and transient exposure, 12 we attempted to explore whether Manα1-2Man-binding lectins, which recognize Man6-9GlcNAc2, affect the neutralization sensitivity of viruses to V3 crown and V2i antibodies. Only tier 1 viruses were chosen for this study as V2i antibodies efficiently neutralize tier 1 viruses exclusively. All the viruses were strongly inhibited by GRFT followed by CV-N and SV-N, suggesting the relative contribution of Man6-9GlcNAc2 glycans on the virus Env.
The higher IC50 values for GRFT followed by CV-N and SV-N can be explained by avidity of these lectins for the Man6-9GlcNAc2 glycan with GRFT recognizing all the three arms D1–D3, followed by CV-N with two binding sites and SV-N recognizing only D3 arm. The strongly enhanced neutralization sensitivity of the viruses to 2557 and 1393A mAb in the presence of Manα1-2Man-binding lectins can be explained because of increased exposure of V3 crown epitope, which lies beneath V1/V2 loop with its epitope-bearing crown pointing toward the inner trimer axis, away from the trimer surface. Furthermore, N-glycans, although not part of V2i epitope, have been shown to point away from V2i in the crystal structure of scaffolded V1/V2, influencing the antibody recognition and function mainly to induce and maintain proper folding of the V1/V2 domain. 13,14
Recent structural data have shown that N-linked glycans cover the entire surface of Env and are important players in the control of antigenicity and immunogenicity of V3 epitopes. Furthermore, the removal of glycans at positions N187 and N197 in the V2 loop and at position N301 on the V3 base has been shown to dramatically increase the exposure of V3 crown epitopes, which are wedged between N301 to N157 glycans. With an increase in the number of V2 glycosylation sites demonstrated in V2i antibody resistance viruses compared with sensitive sites, the additional glycosylation sites being included were mainly in the hypervariable fragments of V2 domain. 9
Lectins in contrast do not interact with the protein component of Env but are exclusively specific to the glycan residues, which, in turn, affect the packing and stabilization of highly flexible crown and stem of V3 and V2 loops, thus affecting their exposure by limiting the mobility of these highly flexible regions. 10 However, we found this effect only in tier 1 viruses but not in tier 2 and tier 3 viruses, possibly other conformational constrainments restrict the accessibility of antibodies to these epitopes.
This study reveals the masking of V3 crown and V2i epitopes by glycan clusters as has been reported previously. Using Manα1-2Man-binding lectins, which stabilize the highly flexible loops on the most proximal surface of HIV-1 Env and allow access of antibodies to these epitopes, resulted in the enhancement of their neutralization potency.
Footnotes
Acknowledgments
This work was supported by grants from Indian Council of Medical Research (ICMR) no. 59/58/2012/Online/BMS/TRM, India, and NIH, United States (PO1AI100151, R24OD018340, and R01AI102740 and R21 A114520). The authors would like to thank Dr. Susan Zola Pazner, Dr. Suman Laal, Dr. Rajnish Kumar, Michael Tuen, and Flavia Camacho for excellent technical assistance. The authors would also like to thank Dr. Barry O'Keefe of Molecular Targets Development Program, Center for Cancer Research, NCI-Frederick for providing CV-N and SV-N. We also thank Constance Williams and Vincenza Itri for providing mAbs.
Author Disclosure Statement
No competing financial interests exist.