
Abstract
A known HIV-1-positive intravenous drug user was found to be human T cell lymphoma/leukemia virus-II (HTLV-II) DNA positive by polymerase chain reaction but seronegative in a screening ELISA. He was consistently DNA positive but took 2 years to fully seroconvert. Sequencing of the HTLV-II strain in his cultured T lymphocytes indicated that it is a prototypical type A strain with no major differences in the long terminal repeat DNA sequence, nor major amino acid differences in the Gag, Env, Tax, and Rex proteins. However, a mutation in its
T
Serial peripheral blood samples were obtained from a known HIV-1-positive intravenous drug user over a 3-year period. He was initially HTLV-I/II seronegative in a screening ELISA but positive for HLTV-II in DNA polymerase chain reaction (PCR) assays performed on peripheral blood mononuclear cells (PBMCs). Over the time studied, he was consistently repeatedly positive by PCR for HTLV-II DNA but took 2 years to fully seroconvert. The HTLV-II copy number was low in his PBMCs (10–100/μg DNA). Hence, we cultured his PBMCs in the presence of IL-2, as previously described, 7 and completely sequenced the HTLV-II strain in his cultured PBMCs to ascertain as to whether any mutations similar to those already described could be detected.
DNA was amplified, with overlapping HTLV-II primer pairs, cloned and sequenced as previously described. 8
The complete DNA sequence of HTLV-II 34510 (GenBank AF412314) was obtained. It is a type A HTLV-II strain. There were no major differences in its long terminal repeat sequences and that of the protypical type A strain, MoT, and the few that occurred were not in major functional sequences (Fig. 1). There were no major amino acid differences between HTLV-II 34510 and HTLV-II MoT in the Gag, Env, Tax, and Rex proteins (Fig. 2). However, a mutation in the HTLV-II 34510 created a stop codon at amino acid 543 of the Pol protein. This region encodes for the RNase function.
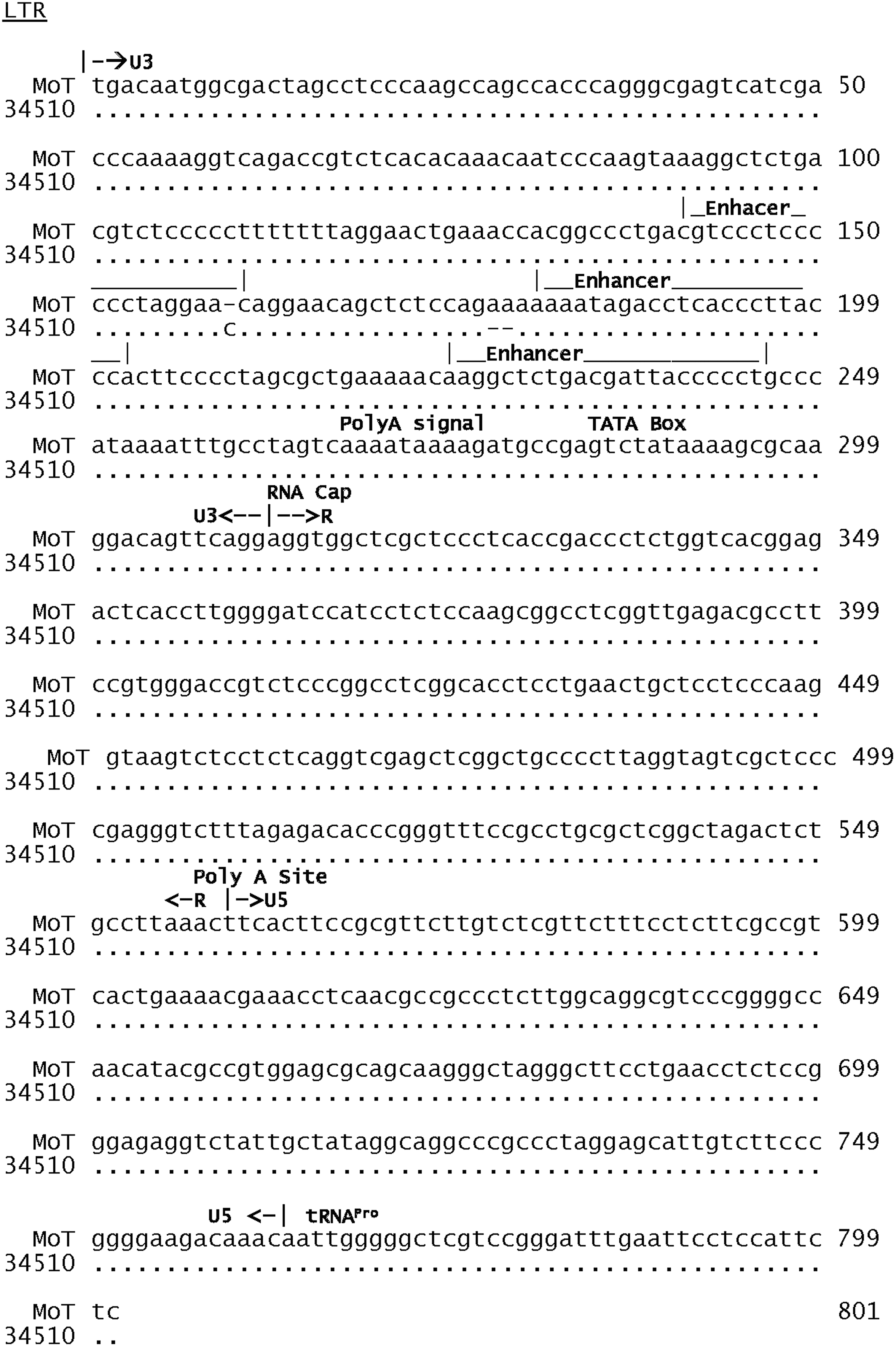
Alignment of the nucleotide sequences of the long terminal repeat regions of HTLV-II MoT and HTLV-II 34510 strains. Conserved bases are shown by the symbol (

Alignment of the amino acid sequences of HTLV-II MoT and HTLV-II 34510 strains. The Tax, Rex, Gag, Pol, and Env proteins are shown. Conserved amino acids are shown by the symbol (·). Amino acid changes are as indicated. The stop codon in the RNase sequence is shown by this symbol (*).


Previously we and others have shown in animal models for STLV-I and HTLV-I infection that mutations in Pol, p27 Rex, and Env proteins can effect replication rates of the virus and seroconversion rates of the recipient animals. 5,6 In the case of the STLV-I experiments mentioned, although a stop-codon mutation in pol was presumably responsible for slow seroconversion, it was not lethal. Both target tantalus monkeys became infected with the mutated virus. In the data presented herein, we have not demonstrated that the mutant HTLV-II 34510 isolate is infectious. However, the fact that it was the only sequence detected in the patient's cultured cells suggests that it is. For both the previously published SLTV-I Tan 90 and the HTLV-II 34510 strains to be replication competent, it would presumably require one or more molecular events, such as read through termination suppression or using a downstream AUG translational start site, for functional Pol proteins to be produced. 9 Such a downstream start site is present not too far from the RNase–RT junction of HTLV-II 34510 Pol RNA (Fig. 3).

Nucleotide and amino acid sequences of HTLV-II 34510 showing a stop codon (*) in the RNase region, and an alternative downstream translational start site (M), which would produce an almost full-length functional reverse transcriptase.
Others have reported on the observation of nonsense mutations, deletions, and insertions in the viral genes of HTLV-I carriers and adult T cell leukemia patients. 10 Similar to our mentioned finding, a G to A mutation was the most common mutation observed. Some of these resulted in stop codons in critical proteins. However, these mutations occurred in minority of proviruses detected in the subjects, and there was no correlation made with seroconversion status of the subject, leading one to assume that they were all seropositive. It is possible that these mutant viral strains could be transmitted and become the dominant strain in some people and result in slow seroconversion because of a low replication rate. Further studies comparing the viral DNA sequences in slow versus rapid seroconverters would be required to determine whether mutant viral strains are a common cause of slow seroconversion in PTLV-infected humans and simians.
Footnotes
Author Disclosure Statement
No competing financial interests exist.