
Abstract
Risk of HIV acquisition varies, and some individuals are highly HIV-1-exposed, yet, persistently seronegative (HESN). The immunologic mechanisms contributing to this phenomenon are an area of intense interest. As immune activation and inflammation facilitate disease progression in HIV-1-infected persons and gastrointestinal-associated lymphoid tissue is a highly susceptible site for transmission, we hypothesized that reduced gut mucosal immune reactivity may contribute to reduced HIV-1 susceptibility in HESN men with a history of numerous rectal sexual exposures. To test this, we used ex vivo mucosal explants from freshly acquired colorectal biopsies from healthy control and HESN subjects who were stimulated with specific innate immune ligands and inactivated whole pathogens. Immune reactivity was then assessed via cytokine arrays and proteomic analysis. Mucosal immune cell compositions were quantified via immunohistochemistry. We found that explants from HESN subjects produced less proinflammatory cytokines compared with controls following innate immune stimulation; while noninflammatory cytokines were similar between groups. Proteomic analysis identified several immune response proteins to be differentially expressed between HIV-1-stimulated HESN and control explants. Immunohistochemical examination of colorectal mucosa showed similar amounts of T cells, macrophages, and dendritic cells between groups. The results of this pilot study suggest that mucosal innate immune reactivity is dampened in HESN versus control groups, despite presence of similar densities of immune cells in the colorectal mucosa. This observed modulation of the rectal mucosal immune response may contribute to lower risk of mucosal HIV-1 transmission in these individuals.
Introduction
W
The most common sites of HIV-1 transmission are mucosal. Rectal mucosa is uniquely susceptible, with per-sexual exposure transmission rates 10 to 100-fold higher than vaginal mucosa. 2 To date, most HESN mucosal studies have focused on vaginal mucosa, with less known about rectal transmission. The mucosa houses many cells of the innate immune system, which utilize pathogen recognition receptors (PRRs) against specific pathogen-associated molecular patterns, leading to cell activation and production of cytokines that can then activate the adaptive immune response.
Mucosal inflammation appears to facilitate HIV-1 transmission. Mucosal infections that trigger PRRs (e.g., Herpes Simplex Virus) have been associated with increased acquisition risk. 3,4 Clinical trials of mucosal microbicides inadvertently causing inflammation have actually increased acquisition risk. 5,6 Furthermore, direct Toll-like receptor (TLR) stimulation or blocking mucosal inflammation increases or decreases vaginal transmission of simian immunodeficiency virus (SIV) in rhesus macaques, respectively. 7,8 Presumably, inflammatory activation of CD4+ T lymphocytes increases target cell susceptibility, since HIV-1 replication depends on transcription factors induced by activation. Thus, it is reasonable to hypothesize that predisposition to inflammation and immune activation at mucosal surfaces affects susceptibility to HIV-1.
In this pilot study, we examine innate immune reactivity in rectal mucosa of HESN men who have sex with men (MSM) versus age-matched control MSM, quantifying innate immune responsiveness, secreted proteins, and immune cell composition in the rectal mucosa. We found lower proinflammatory mucosal cytokine responsiveness and differential secreted protein expression in HESN, but similar immune cell composition of the mucosa. To our knowledge, this is the first description of an immune quiescent phenotype in the rectal mucosa of HESN men.
Materials and Methods
Study participants and samples
All subjects were enrolled after providing informed consent under a UCLA Institutional Review Board approved protocol. The HESN group included MSM recruited from the original Multicenter AIDS Cohort Study (MACS) and were designated HESN based on history of unprotected receptive anal intercourse (RAI) during a time of high HIV-1 prevalence. This group was defined by the top 10th percentile of unprotected anal receptive sexual exposures in the previous 2.5 years at MACS enrollment (1984–1985) and has been the subject of numerous other HESN studies. 9 –12 All HESN have been tested and negative for CCR5 Δ32 mutation. The control group included age-matched MSM who denied history of multiple high-risk sexual exposures and were recruited from the UCLA Center for AIDS Research Mucosal Immunology Core Laboratory Registry. Before enrollment, all subjects were confirmed HIV-1 seronegative and screened for absence of active Neisseria gonorrhoeae or Chlamydia trachomatis infection via urine and rectal nucleic acid amplification test. Enrolled subjects then participated in two study visits spaced 4–6 weeks apart for collection of colorectal mucosal biopsies via sigmoidoscopy (30 biopsies collected from rectum up to 30 cm from the anal verge).
Ex vivo mucosal explant immune stimulations
Gut mucosal biopsies were immediately processed as previously described 13,14 and cultured in triplicate in standard tissue culture plates using RPMI 1640 medium with 2.5 mg/ml amphotericin B and 0.1 mg/ml piperacillin–tazobactam in a 37° humidified incubator. Whole biopsy explants were exposed in triplicate overnight to innate immune receptor ligands (InvivoGen) or inactivated pathogens at concentrations previously confirmed to induce inflammatory response in control peripheral blood mononuclear cells: lipopolysaccharide 10,000 EU/ml (TLR-4), imiquimod 5 μg/ml (TLR-7), CpG oligodeoxynucleotides 25 μg/ml (TLR-9), muramyldipeptide 10 μg/ml (NOD-2), HIV-1BaL (3 × 103 tissue culture infectious dose50 [TCID50]), C. trachomatis (5 × 108 inclusion forming units [IFU]/ml), and HSV-2 (3 × 107 plaque forming unit [PFU]). HIV-1BaL was prepared via standard transfection methods and then heat inactivated. Whole inactivated C. trachomatis was kindly provided by Dr. Kathleen Kelly (UCLA) and whole inactivated HSV-2 was kindly provided by Dr. Ren Sun (UCLA).
Cytokine quantification
Supernatants were collected from stimulated explants after 18 h and immediately stored at −80°C. Cytokines were quantified using multiplex high-sensitivity Luminex arrays (R&D Systems). All samples were batched and run together to minimize interassay variability. Data were then normalized to the untreated condition for each subject and expressed as Fold Change.
Mass spectrometry and proteomics analysis
Explant supernatants were purified of debris via ultracentrifugation, followed by trypsin digestion using the filter aided sample preparation (FASP) method
15
and requantification at the peptide level before label-free mass spectrometry (MS) analysis. Preparation of rectal biopsy tissue and rectal sponge samples is described in “Supplementary Methods” (Supplementary Data are available online at
Immunohistochemistry and quantitative analysis
Gut mucosal biopsies were collected as described above, then immediately embedded in OCT medium, and snap-frozen in liquid nitrogen. Tissue was sectioned into 4 μm cryostat sections and stained following heat-induced antigen retrieval with the following: CD3 (polyclonal; Dako), CD4 (4B12; ThermoFisher), CD8 (C8/144B; Dako), CD68 (PG-M1; Dako), and CD209 (DCN46; BD Biosciences). Signal was detected using Envision+ HRP (Dako), then slides were digitized at 20 × magnification using Aperio ScanScope AT (Leica Biosystems), and morphometric analysis performed using Tissue Studio (Definiens, Inc.).
Statistical analyses
Study population demographic characteristics were compared using unpaired t-tests for numerical data and Fisher's exact test for nominal data (Table 1). Cytokine data were analyzed with nonparametric Wilcoxon matched-pairs test for each cytokine pairing each unique stimulus and corrected for multiple comparisons using Benjamini–Hochberg procedure (FDR 0.1). Immunohistochemistry quantitative analyses were compared using unpaired t-tests. All statistical analyses were performed using GraphPad Prism 7 (GraphPad Software).
Number of sexual partners and frequency in year preceding enrollment in current study.
Number of unprotected RAI partners in the 2 years preceding enrollment in MACS. This data were used to determine HESN designation.
SD, standard deviation; NA, not available; MACS, Multicenter AIDS Cohort Study; HESN, HIV-1-exposed seronegative.
Results
Subjects
HESN MSM (n = 4) were MACS participants within the upper 10th percentile for self-reported RAI partners in the 2.5 years before MACS enrollment in 1984–1985. 9,17 Age-matched control MSM (n = 12) without reported high-risk sexual exposure history were recruited from the UCLA Mucosal Immunology Core Registry. The HESN group is distinguished based on history of frequent unprotected RAI during a time of high HIV-1 prevalence, while the control group denied similar history. At the time of this study, most HESN individuals had recent sexual exposures compared with the control individuals. Routine laboratory examinations, including prevalence of chronic viral and sexually transmitted infections, were similar between the two groups (Table 1).
Rectal mucosal explants from HESN produced minimal proinflammatory cytokines following innate immune stimulation
Cytokines are critical regulators of immune homeostasis in the mucosa, and even small changes in magnitude or timing of expression can alter the inflammatory balance. 18 To characterize colorectal mucosal innate immune reactivity, cytokine production from freshly acquired colorectal biopsy explants was quantified 18 h after ex vivo stimulation with selected innate immune ligands or whole inactivated pathogens. HESN explants produced fewer cytokines overall compared with controls, in particular, in response to stimulation with TLR-4, TLR-7, TLR-9, and NOD-2 (Fig. 1). We further subclassified cytokines as proinflammatory versus noninflammatory and noted that most observed differences between HESN and control explants were in cytokines classically involved in proinflammatory response. Among the cytokines with significant differences were interleukin (IL)-6 (light blue bars) and IL-1β (purple bars), both of which have well-known roles in mucosal inflammation. 19 All explants produced similar amounts of noninflammatory cytokines IL-10, IL-4, and IL-5. As cytokines are innate immune signaling molecules, even small changes in expression may have significant downstream functional implications.
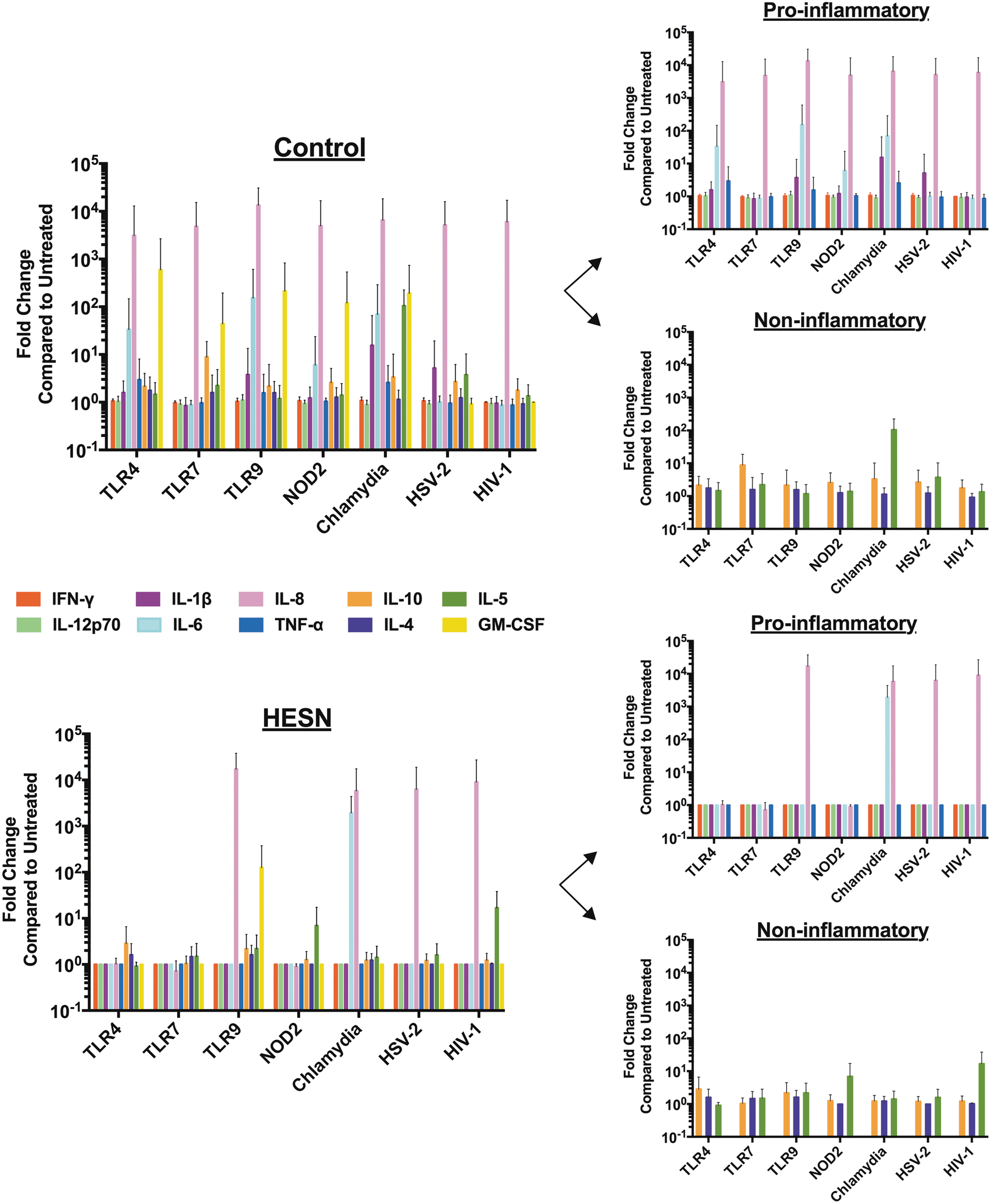
Reduced cytokine production in HESN rectal mucosa following innate immune stimulation. Freshly acquired explants from control versus HESN subjects were cultured with various innate immune stimuli (x-axis) overnight and cytokine release assessed with Luminex arrays. HESN explants produced significantly less proinflammatory cytokines IFN-γ (P = .015, adjusted .039), IL-1β (P = .02, adjusted .039), IL-12p70 (P = .02, adjusted .039), and IL-6 (P = .031, adjusted .062). Other cytokines TNF-α (P = .195) and IL-8 (P = .468) were decreased, but did not reach significance. Similar amounts of noninflammatory cytokines IL-10 (P = .0716), IL-4 (P = .078), and IL-5 (P = 0.687) were produced by all explants. Data graphed as mean (colored bars) ± standard deviation. Fold Change calculated based on untreated condition for each participant. HESN, HIV-1-exposed seronegative; IFN-γ, interferon-gamma; IL, interleukin; TNF-α, tumor necrosis factor-alpha.
Differential protein expression in colorectal mucosal explants from HESN compared with controls
Mucosal fluid contains numerous soluble innate immune factors, such as chemokines and defensins, many of which have been correlated with protection against HIV-1 infection in HESN subjects. 20 –22 While these factors have been well described in other mucosal sites such as the female genital tract, the soluble innate immune factor environment in rectal fluid has only recently been detailed. 15 We used proteomic analysis by MS of explant supernatants following ex vivo stimulation with inactivated HIV-1 as another assessment of innate immune reactivity. Of the 176 human proteins observed, 8% (n = 14) were differentially expressed between groups; five were overabundant, and nine were underabundant in HESNs (Fig. 2). While these differences were not statistically significant after correction for multiple comparisons, functional annotation of these proteins was consistent with lower immune activation in HESN mucosa, with relative reduction of several downstream inflammatory and antimicrobial proteins, including complement component C4, lactotransferrin, and intelectin (Fig. 2B). Among the overabundant proteins, some proteins were involved in inflammation, fibrinogen, and cathepsin B. These proteins were uniquely expressed in HESN and, in conjunction with the reduction in other inflammatory proteins, again suggest an altered inflammatory response in HESN. Of note, the MS methods are not optimized to detect cytokines, and such proteins do not pass data filtering.
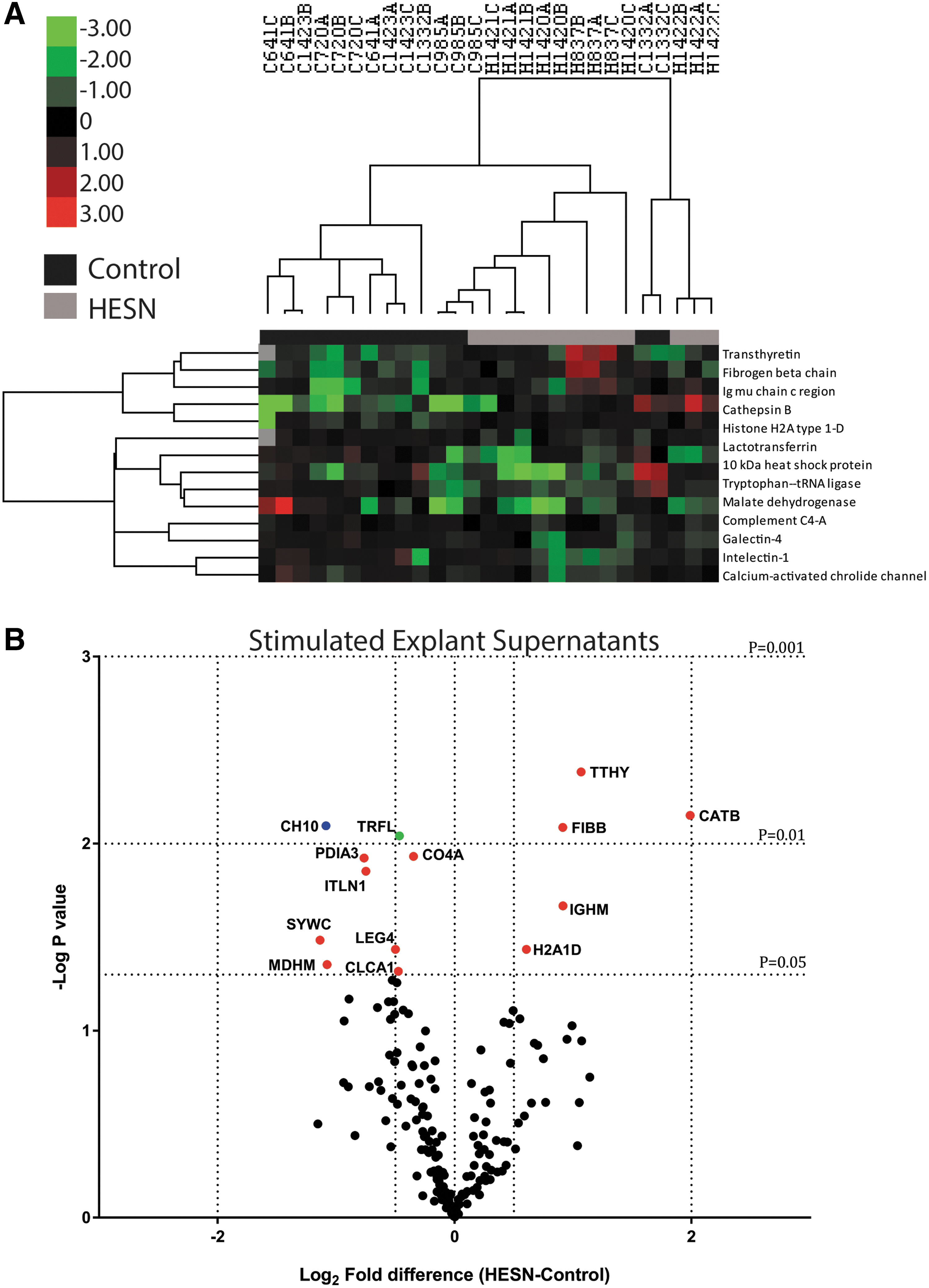
Proteomics analysis of secreted mucosal proteins in HESN and control MSM.
We also examined the baseline proteomic profile of the rectal mucosa in HESN and control MSM by performing similar proteomic analysis on rectal fluid and unstimulated whole rectal biopsy tissue. The proteomic signature of HESN rectal tissue and secretions is distinct from that of control rectal tissue, as shown by hierarchical clustering (Supplementary Figs. S1A and S2A). Functional annotation did not reveal any significant differences in immune/inflammatory pathways, although notably one of the most significant decreased proteins was lactotransferrin (Supplementary Figs. S1B and S2B), which was also decreased in HESN following HIV-1 stimulation (Fig. 2B). The implication of these observed differences is the focus of a separate study.
Densities of immune cells are similar between HESN and control colorectal mucosa
To determine if the observed differences between HESN and controls were due to differing mucosal immune cell populations, quantitative immunohistochemistry was performed on colorectal mucosal samples. Both total T cells (CD3+) and CD4+ and CD8+ subsets were present at comparable densities between groups (Fig. 3 and data not shown). To examine the antigen presenting cell (APC), we selected a general macrophage marker (CD68) and used CD209 as a dendritic cell marker because of its known role in HIV-1 infection. Overall populations of macrophages (CD68+) and dendritic cells (CD209+) were also comparable (Fig. 3), suggesting relative normal densities of innate immune cells and adaptive immune targets of HIV-1 infection in HESN.
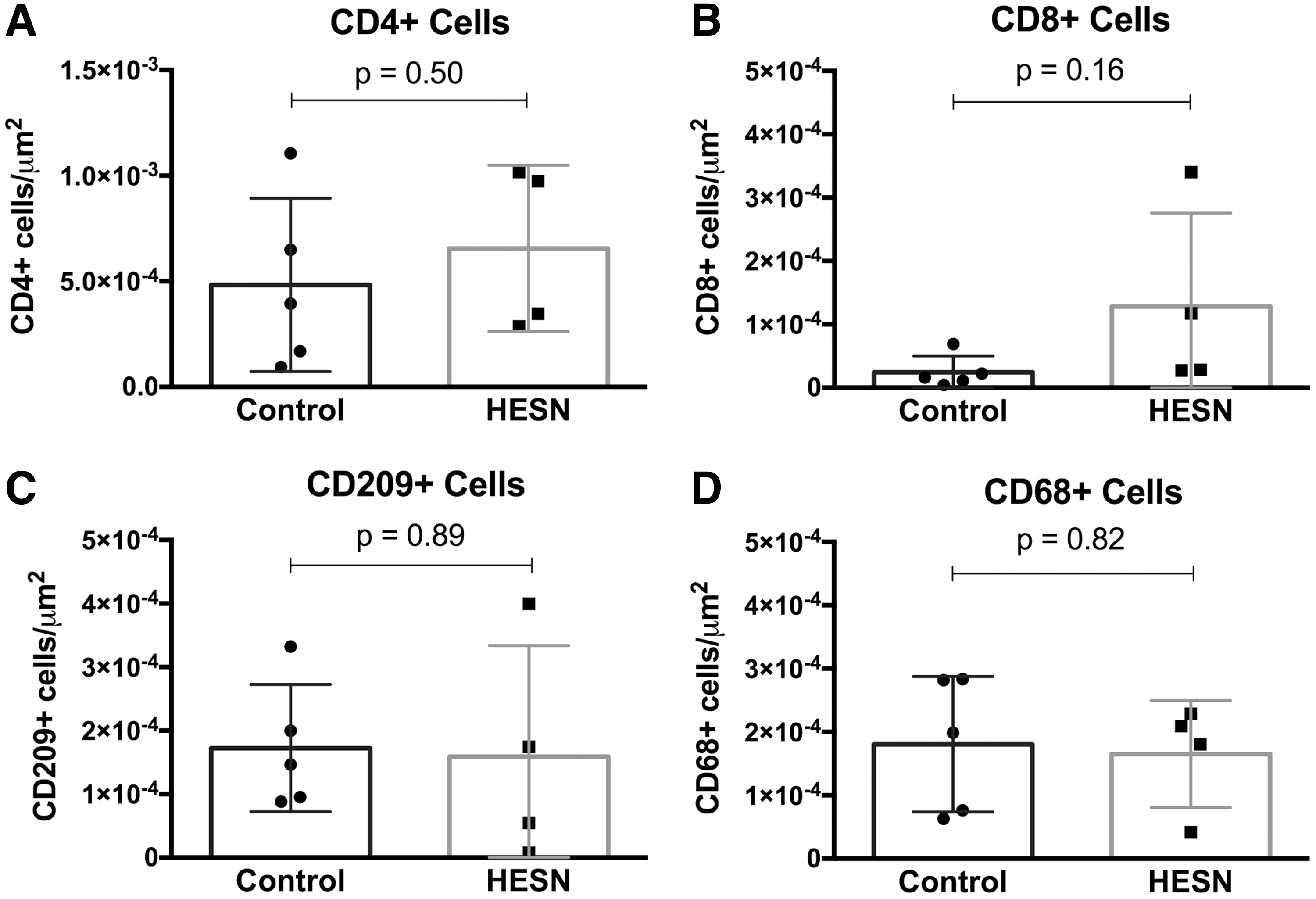
Immune cell densities in rectal mucosa from HESN and control MSM. Quantitative immunohistochemistry shows similar densities of immune cell populations in rectal tissue between HESN and selected age-matched control subjects. Freshly acquired rectal mucosal biopsies were snap frozen, then stained for
Discussion
Several HESN studies have suggested the seemingly paradoxical “immune quiescence” hypothesis, in which low immune reactivity (rather than effective antiviral immunity) mediates protection. Most have shown lower T cell activation and cytokine production in blood, 23 –27 while data on mucosal immunity are fewer and limited to vaginal mucosa. 28,29 Suggested mediators include increased regulatory T cells, 25 antiproteases, 20 –22 and alterations in interferon regulatory factor-1 expression. 30 The totality of these findings suggest that HESN exhibit lowered immune reactivity, reducing the pool of activated target cells susceptible for HIV-1 infection and spread, 31,32 although this hypothesis remains controversial, and others have suggested that ongoing exposure can mediate protection through maintaining cellular immunity against HIV-1, therefore, with increased immune activation. 33 –35
In this pilot study, we show dampening of rectal mucosal innate immune responsiveness in HESN MSM, as evidenced by low proinflammatory cytokine production and a unique proteomic profile, which is unrelated to gross differences in mucosal densities of T lymphocytes or antigen-presenting cells. While our MS methods were not optimized to detect cytokine proteins, we were able to quantify unique changes in other immunomodulatory proteins, including those previously identified in other HESN and transmission studies. 36,37 Collectively these data suggest an overall reduced mucosal inflammatory response following ex vivo challenge. The reduced proinflammatory cytokine production agrees with prior studies, 27,29 although this is the first description in the rectal mucosa and the first in HESN persons without ongoing high levels of sexual exposure. These points suggest that cytokines may modulate risk of HIV-1 transmission, be it via direct influence on viral permissiveness of resident target cells or chemoattraction of suitable target cells. 38,39 As our cohort did not report unusual or repeated infections or ongoing frequent risky exposures to HIV-1, this likely represents intrinsic mucosal immune quiescence without global immunodeficiency.
The mechanism underlying the dampened immune responsiveness in HESN remains unclear. Prior studies have identified alterations in T cell signaling and activation, chemokine/cytokine signaling, including interferon, and PRR expression as the drivers of this phenotype; however, the causal factors triggering these changes in HESN are not known. 27,29,40,41 Ongoing sexual exposure may mediate immune activation independent of infections. Camara et al. suggested that a cohort of HESN were protected by higher condom use, and lower immune activation was simply a marker of reduced unprotected sexual exposure. 24 This would not explain our findings in HESN men with a distant history of high levels of unprotected exposure and minimal ongoing exposure. More recent data show that sexual exposures alter the microbiome, in turn, influencing immune reactivity. 42 Without ongoing high-risk exposures, the data suggest that our HESN (1) have an intrinsic difference in innate immunity or (2) exhibit a persisting difference resulting from cumulative exposures. An analysis of the effects of repeated RAI on rectal mucosal immune response is the subject of ongoing investigation.
Our study is subject to caveats. The number of subjects in our pilot study was limited due to stringency of HESN criteria and loss to follow-up of MACS participants since 1985. Similarly, the ethnic diversity in our HESN group was limited largely by the homogenous makeup of the MACS cohort, and thus, we cannot exclude the potential confounding influence of ethnicity differences between the two groups. Although general differences in cytokine production and proteomics were statistically significant, the small number of subjects precluded statistical power to identify specific biomarkers or direct pathways. The analysis of mucosal immune cells was limited to broad categories and did not break down specific subtypes (e.g., dendritic cell subtypes) or examine the activation status of cells. Further definition of mucosal immune cell composition is ongoing. Despite these caveats, this study provides some of the first data on immune activation and proteomics in the rectal compartment of HESN MSM.
Footnotes
Acknowledgments
We are grateful to all the participants for their time and willingness to participate in this study. We also acknowledge the support of Dr. Roger Detels, Dr. Otoniel Martinez-Maza, and John Oishi with the Los Angeles MACS. We also thank Dr. Garrett Westmacott and Stuart McCorrister from the National Microbiology Laboratory Mass Spectrometry Core Facility for technical support. This work was supported, in part, by National Institutes of Health/National Institute of Allergy and Infectious Diseases (NIH/NIAID) grant AI100638 to O.O.Y. J.A.F. was supported, in part, by NIH training grants T32AI089398 and T32MH080634. Additional support from UCLA AIDS Institute/Center for AIDS Research Seed Grant (supported by NIH program grant AI028697) to J.A.F., a grant from AIDS Healthcare Foundation to O.O.Y., and a Team Grant from Canadian Institutes of Health Research to A.D.B. (TMI 317268). The UCLA Mucosal Immunology Core provided developmental funds and technical support (NIH/NIAID AI028697 CFAR).
Author Contributions
J.A.F., L.R., J.C.H., and J.E. performed experiments and analyzed the data. T.S. contributed to clinical study design and managed acquisition of clinical specimens. J.A.F., J.C.H., P.A.A., O.O.Y., conceived and designed the study. J.A.F., L.R., A.D.B., P.A.A., O.O.Y., wrote and edited the article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.