
Abstract
The induction of both neutralizing antibodies and non-neutralizing antibodies with effector functions, for example, antibody-dependent cellular cytotoxicity (ADCC), is desired in the search for effective vaccines against HIV-1. In the pursuit of novel immunogens capable of inducing an efficient antibody response, rabbits were immunized with selected antigens using different prime–boost strategies. We immunized 35 different groups of rabbits with Env antigens from clinical HIV-1 subtypes A and B, including immunization with DNA alone, protein alone, and DNA prime with protein boost. The rabbit sera were screened for ADCC activity using a GranToxiLux-based assay with human peripheral blood mononuclear cells as effector cells and CEM.NKRCCR5 cells coated with HIV-1 envelope as target cells. The groups with the highest ADCC activity were further characterized for cross-reactivity between HIV-1 subtypes. The immunogen inducing the most potent and broadest ADCC response was a trimeric gp140. The ADCC activity was highest against the HIV-1 subtype corresponding to the immunogen. The ADCC activity did not necessarily reflect neutralizing activity in the pseudovirus-TZMbl assay, but there was an overall correlation between the two antiviral activities. We present a rabbit vaccination model and an assay suitable for screening HIV-1 vaccine candidates for the induction of ADCC-mediating antibodies in addition to neutralizing antibodies. The antigens and/or immunization strategies capable of inducing antibodies with ADCC activity did not necessarily induce neutralizing activity and vice versa. Nevertheless, we identified vaccine candidates that were able to concurrently induce both types of responses and that had ADCC activity that was cross-reactive between different subtypes. When searching for an effective vaccine candidate, it is important to evaluate the antibody response using a model and an assay measuring the desired function.
Introduction
T
The ability of an HIV-1 vaccine to generate a broad functional anti-HIV-1 response is crucial. HIV-1 is extremely variable and has a high evolution rate. HIV-1 is divided into four groups: M, O, N, and P. The M group, which accounts for the vast majority of HIV-1 infections, contains nine subtypes (A through J), ∼79 circulating recombinant forms (CRFs), and multiple unique recombinant forms. 22 Moreover, HIV-1 infection is characterized by a high degree of genetic variability within the infected individual. Immune responses able to meet this high diversity of HIV-1 are crucial for any efficient HIV-1 vaccine.
The growing knowledge of conserved ADCC and neutralizing epitope structures has not yet translated into improved immunogens, emphasizing the importance of continuing empiric approaches in the search for relevant HIV-1 vaccine immunogens. There is an overlap between epitopes recognized by ADCC-mediating and neutralizing antibodies, with many ADCC-mediating antibodies also mediating neutralization, but there are also many ADCC-specific epitopes and probably many more yet to be defined. 23,24
We focused on identifying and characterizing immunogens from clinical HIV-1 isolates that elicited broadly neutralizing antibodies in HIV-1-infected individuals. In this study, we screened their capacity to induce anti-HIV-1 antibodies in rabbits with ADCC function and compared the results with their IgG binding titers and neutralizing ability (part of the data previously published in Heyndrickx et al., 25 Visciano et al., 26 and Borggren et al. 27 ). The groups with the highest ADCC activity were further characterized for cross-reactivity between HIV-1 subtypes.
Materials and Methods
Rabbit immunizations
All rabbit experiments were performed in accordance with the Animal Experimentation Act of Denmark (permit No. 2007/561-1362) and European Convention ETS 123 (Protection of Vertebrate Animals used for experimental and other scientific purposes). Ten-week-old New Zealand White nulliparous female rabbits (Charles River Laboratories) were housed at the animal facility at Statens Serum Institut (SSI), Copenhagen, Denmark with an acclimatization period of at least 10 days. Groups of three to four rabbits were each immunized with DNA alone, protein alone, or combinations of DNA prime followed by protein boost. One group was immunized with virosomes. All groups of animals and immunogens are listed in Table 1. Neutralization and IgG titers have previously been published for 13 groups out of the 35 groups. These 13 groups include the 9 groups immunized with gp140 trimer protein, 1 group immunized with gp120 monomer protein 94UG018, and 3 groups immunized with DNA, gp150SOSIP-BX08 and SSImix140 (Table 1). ADCC data are new for all 35 immunization groups.
Patient from where the immunogen was derived.
ADCC for each animal, defined as the highest percentage of granzyme B activity after background subtraction (i.e., preimmunization sera tested at the same dilution). The immunogens were derived from individuals infected with HIV-1 subtypes A or subtype B, and the screening of ADCC activity was performed with a matching recombinant subtype A (UG37) and subtype B (BaL) envelope protein. Two groups were immunized with a combination of subtype B and subtype A derived immunogens, there ADCC against subtype B (BaL, left column), as well as subtype A (UG37, right column), is shown. Values below the threshold for positivity, that is, above 6.2% granzyme B, are not shown.
Neutralization against SF162. IC50 IgG, μg/ml.
Anti-rgp120IIIb IgG titer.
ADCC, antibody-dependent cellular cytotoxicity; CAF01, cationic adjuvant formulation 1; CAF02, cationic adjuvant formulation 2; e.p., electroporation; i.d., intradermally; int., intensive prime; ND, not done; s.c., subcutaneously; SSI, Statens Serum Institut.
DNA immunizations were performed intradermally (200 μg/dose) with subsequent electroporation using the OncoVet™ system (CytoPulse Sciences/Cellectis, Romainville, France) over the injected area. Protein immunizations were performed subcutaneously (s.c.) with monomeric or trimeric Env protein (20 or 100 μg/dose) in the presence (or absence) of cationic adjuvant formulation number 1 (CAF01) 28 or cationic adjuvant formulation number 2 (CAF02) containing 100 μg/dose distearoyl-sn-glycero-3-phosphocholine (DSPC) 29 in a total volume of 400 μl. Virosomes 30 were injected s.c. (200 μg/dose). Unless otherwise stated, immunizations were performed on weeks 0, 4, 8, and 12. DNA priming was performed thrice during the first week, a mode of “intensive” priming. 27 The protein boost was performed on week 14. Ear bleeding was performed before immunization (week 0) and 2 weeks after the last immunization (week 14 or 16), and all sera were stored at −20°C.
ADCC assay
The ADCC-GranToxiLux assay (OncoImmunin, Gaithersburg, MD) was performed as described by Pollara et al. 31 In brief, the target cells, CEM.NKRCCR5 cells, were coated with HIV-1 envelope protein and labeled with TFL4 and NFL1. Recombinant HIV-1 representing the envelopes of subtype A, gp140 UG37 (Polymun Scientific, Klosterneuburg, Austria); subtype B, gp120 BaL (Immune Technology, New York, NY); subtype C, gp120 ZA1197MB (Immune Technology); and subtype CRF01_AE, gp120 CM243 (Protein Sciences, Meriden, CT) were used. The protein concentration used for coating (0.01 mg/ml) was decided by competing the binding of FITC-conjugated anti-CD4 antibodies (clone SK3; BD Biosciences), as described. 31
Human peripheral blood mononuclear cells (PBMCs) were used as the source of NK cells, that is, effector cells. The PBMCs were obtained by density-gradient centrifugation of buffy coats from the Danish Blood Bank and cryopreserved until use. Before use, the PBMCs were thawed and rested overnight in R10 (RPMI; Gibco, Life Technologies, Naerum, Denmark) supplied with 10% fetal bovine serum (Gibco) and 1% penicillin–streptomycin (Gibco) at 37°C and 5% CO2. The effector (E) and target (T) cells were tested at an E:T ratio of 30:1. Rabbit sera were tested in fivefold dilutions starting at 1:10. The positive control HIV IgG immunoglobulin (NIH AIDS Research and Reagent Program) and negative control (in-house pool of HIV-negative sera) were tested in fivefold dilutions starting at 1:250. A control without antibody was also included to confirm that there was no antibody-independent activity of NK cells against the target cells. All sera were heat inactivated (1 h at 56°C).
The cells were acquired using a BD LSRII and analyzed using FlowJo (Tree Star version 8.8.7). The threshold for a positive GzB response was >6.2% after background subtraction. These criteria were established using preimmunization rabbit samples (n = 348) and defined as the above mean plus three standard deviations. ADCC for each animal was defined as the highest percentage of granzyme B activity of the effector cells after background subtraction (i.e., preimmunization sera tested at the same dilution). For comparison, the granzyme B activity was also calculated as the area under the curve (AUC) and as the highest dilution factor above the threshold.
Anti-HIV gp120 enzyme-linked immunosorbent assay
Anti-gp120-specific IgG titers were determined by enzyme-linked immunosorbent assay (ELISA), as previously described. 27
Neutralization assay
Neutralization of purified IgG was assayed in pseudovirus neutralization assays using TZMbl cells conducted as described previously. 25
Statistical analysis
The data analysis was performed using GraphPad Prism 6 software version 6.0c (GraphPad Software, Inc., La Jolla, CA). Spearman's two-tailed test was used to test for correlations.
Results
ADCC activity in rabbits immunized with HIV-1 envelope immunogens
Thirty-five groups of immunized rabbits were screened for serum ADCC activity. The HIV-1 immunogens are listed in Table 1 and include envelope DNA, protein (gp120 monomer, gp140 trimer, or gp140 expressed by virus-like particles), or a combination of DNA and protein (DNA prime followed by protein boost). The immunogens were derived from individuals infected with HIV-1 subtypes A or subtype B, and the screening of ADCC activity was performed with a matching recombinant subtype A (UG37) and subtype B (BaL) envelope protein. ADCC activity was determined 2 weeks after the last immunization, that is, week 14 or 16, and a preimmunization serum from the same animal was used as background.
We assessed the ADCC-mediating activity in rabbit sera in three different ways: the highest percentage of Granzyme B activity, the magnitude of ADCC activity (by calculating the AUC), and by calculating the last dilution with ADCC activity above the threshold of 6.2%. All three analytic methods correlated well with each other (p < .0001, Supplementary Fig. S1; Supplementary Data are available online at
In 19 out of the 35 groups, ADCC activity was detected in at least 1 animal (Table 1). In eight groups, ADCC activity was detected in all animals; in three groups, ADCC was observed in three out of four animals; in four groups, ADCC was observed in two out of four animals; and in four groups, ADCC was observed in only one animal. Although the experiments were not designed to compare the same immunogen in different forms and/or immunization regimens, gp140 trimer and DNA prime plus protein boost appeared to be the most potent inducers of ADCC-mediating antibodies, with 53% and 75% of the rabbits, respectively, having induced ADCC-mediating antibodies (Fig. 1).
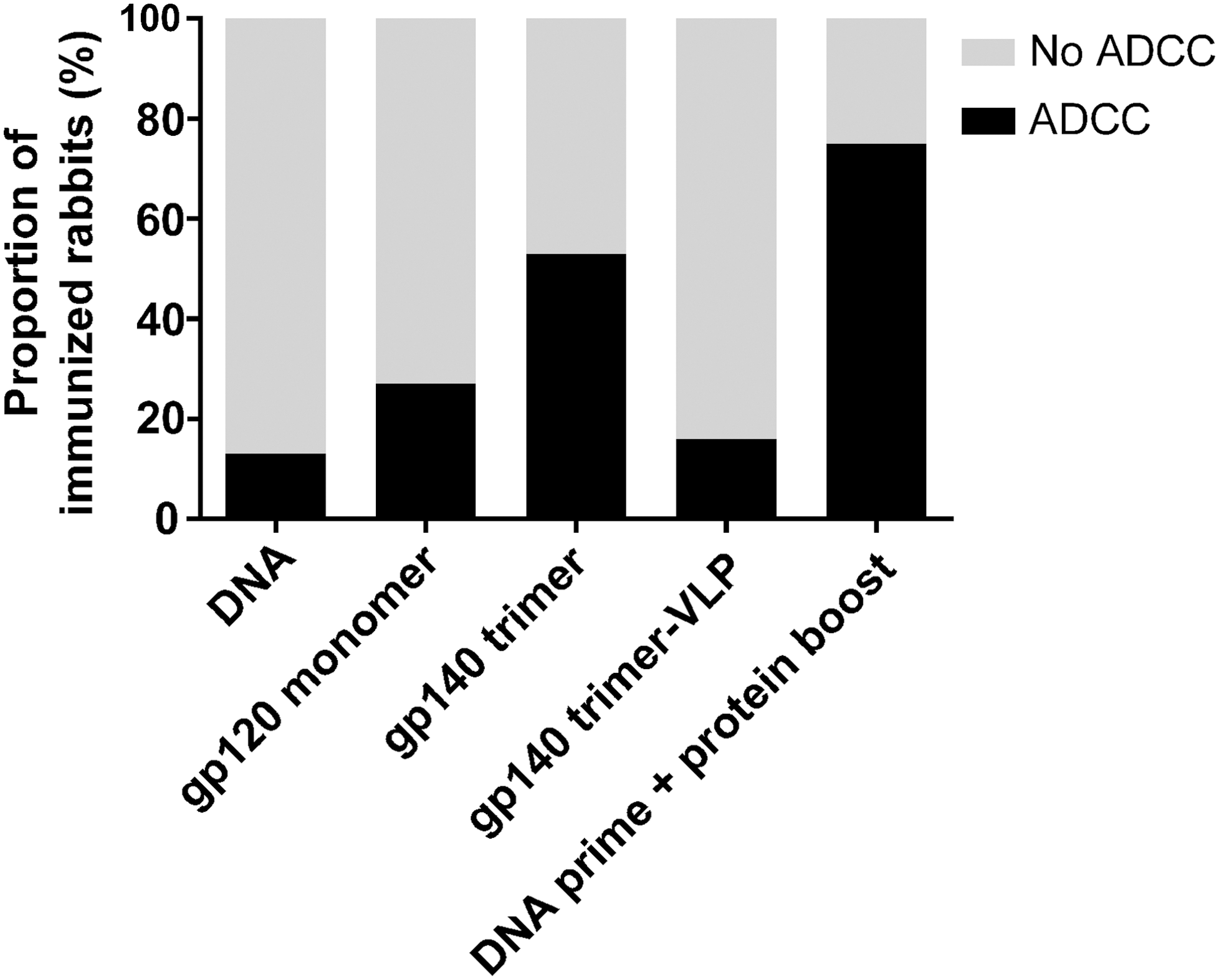
Trimeric proteins with or without DNA prime were the most potent inducers of ADCC-mediating antibodies. Thirty-five groups of immunized rabbits were screened for serum ADCC activity. In this study the data are grouped according to the type of immunogen: envelope DNA, gp120 monomer, gp140 trimer, gp140 expressed by virus-like particles, or DNA prime followed by protein boost (details described in Table 1). The immunogens were derived from individuals infected with HIV-1 subtype A or subtype B, and the screening of ADCC activity was performed with a matching recombinant subtype A (UG37) and subtype B (BaL) envelope protein. ADCC activity was determined 2 weeks after the last immunization, that is, week 14 or 16, and a preimmunization serum from the same animal was used as background. The proportion of animals in which an ADCC response was detected (black bar) or was not detected (gray bar). ADCC, antibody-dependent cellular cytotoxicity.
Correlation among ADCC activity, neutralization capacity, and IgG binding titers
To determine if the ADCC activity in the sera from the immunized rabbits correlated with the neutralizing activity, we compared the ADCC results with the neutralization capacity against the HIV-1 subtype B strain SF162. Neutralization was measured in 22 out of the 35 groups of rabbits using purified IgG in a pseudovirus-TZMbl assay (part of the data previously published in Heyndrickx et al., 25 Visciano et al., 26 and Borggren et al. 27 ). As the neutralization was performed using a subtype B strain, we correlated neutralization with ADCC in animals immunized with subtype B-derived immunogens. High ADCC activity did not necessarily result in high neutralization capacity, and vice versa. Nevertheless, when including all available data, there was a correlation between ADCC and neutralization capacity against SF162 (p = .0332, r = −0.3113) (Fig. 2A and Table 1).
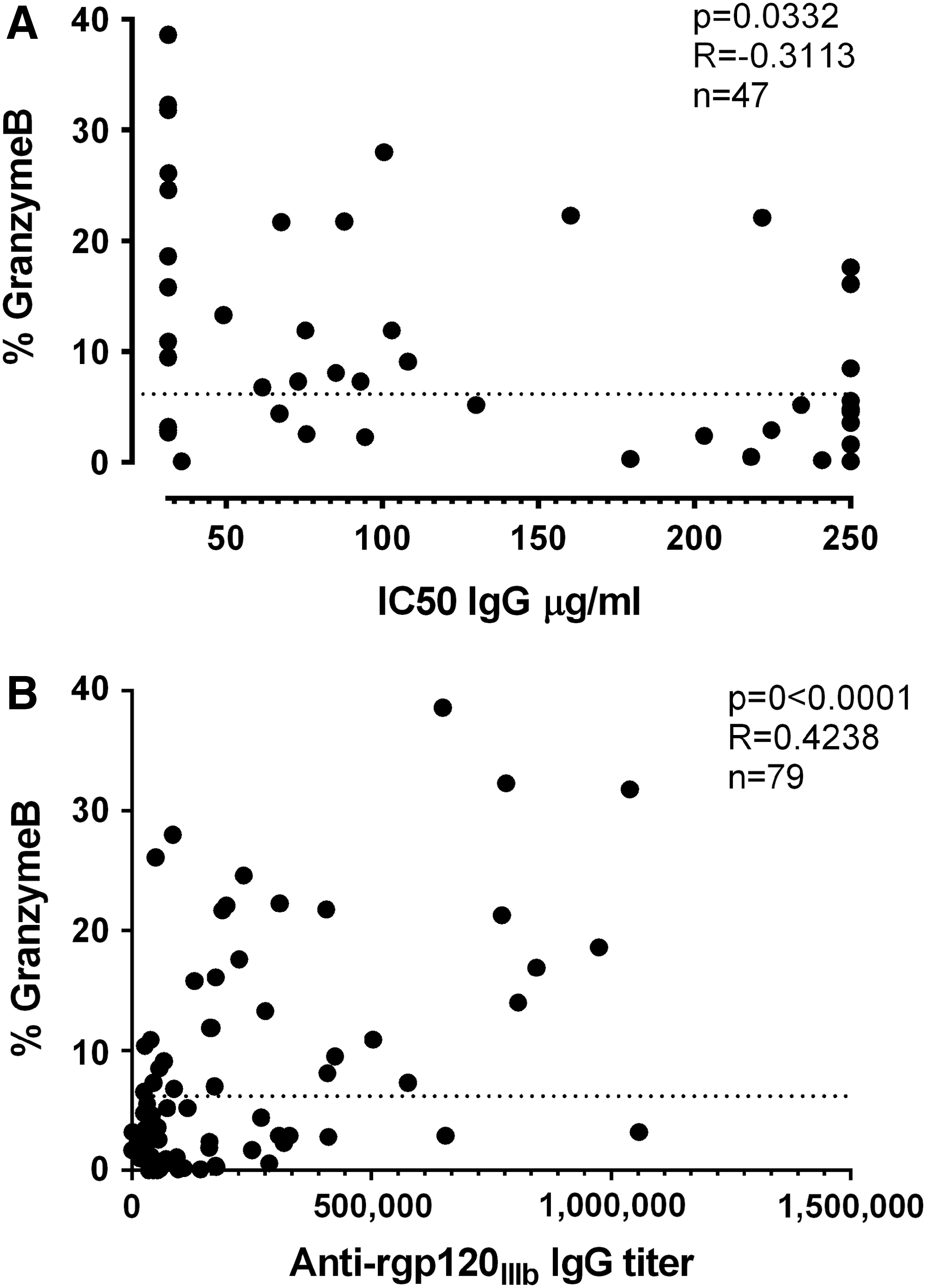
ADCC activity versus neutralization and binding titers.
Similarly, we correlated ADCC activity with subtype B HIV-1 envelope (IIIB)-specific IgG titers, as measured by ELISA (Fig. 2B and Table 1) (part of the data previously published in Heyndrickx et al., 25 Visciano et al., 26 and Borggren et al. 27 ). We correlated ADCC activity with the ELISA IgG titers in the animals immunized with subtype B-derived immunogens. There was a significant correlation between ADCC activity and IgG titers (p < .0001, r = 0.4238). This correlation was partly driven by the fact that if there was little or no HIV-1 envelope-specific IgG present in sera, there was also no ADCC activity. Nevertheless, even when selecting only the animals with ADCC activity above the threshold for positivity (above 6.2% granzyme B), a correlation between ADCC activity and IgG titer was still observed (p = .0136, r = 0.4319).
The ADCC responses in immunized rabbits were cross-reactive between different subtypes of HIV-1
A desired feature of an effective HIV-1 vaccine is breadth, that is, the induction of an immune response that is cross-reactive between different strains and subtypes. We selected the seven groups of rabbits with the highest of ADCC response against subtype matched HIV-1 envelope and tested cross-reactivity to other HIV-1 subtypes. Four of these groups were immunized with protein, including the following: (I) trimeric 136-3 gp140 in CAF01 adjuvant, (II) monomeric IIIB gp120 in CAF02, (III) trimeric ACS19554 gp140 in CAF01, and (IV) trimeric ITM1_4 gp140 in CAF01 adjuvant. The remaining three groups were immunized with DNA followed by a trimeric protein boost in CAF01 adjuvant: (V) Bx08 gp140 DNA followed by Bx08 gp140 trimer, (VI) Bx08 gp150 DNA followed by Bx08 gp140 trimer, and (VII) NIBSC 40-9 gp150 DNA followed by the corresponding protein (Table 1). These groups were tested for ADCC activity against HIV-1 envelopes from subtype A (UG37), B (BaL), C (ZA1197), and CRF01_AE (CM243). Two groups, monomer IIIB and trimer 136-3, displayed the broadest ADCC response, with cross-reactivity between the envelopes of the four different clades tested (Fig. 3), that is, clades A, B, C, and CRF01_AE. Three groups, trimer ACS19554, trimer ITM1_4, and Bx08 gp140 DNA plus protein, induced an ADCC response that was cross-reactive among subtypes A, B, and C. The last two groups, Bx08 gp150 DNA plus protein and NIBSC 40-9 DNA plus protein, induced a narrower response against the matched subtype plus one more (Fig. 3). In all cases, the response was the most robust against the subtype corresponding to the immunogen and the lowest against subtype E.
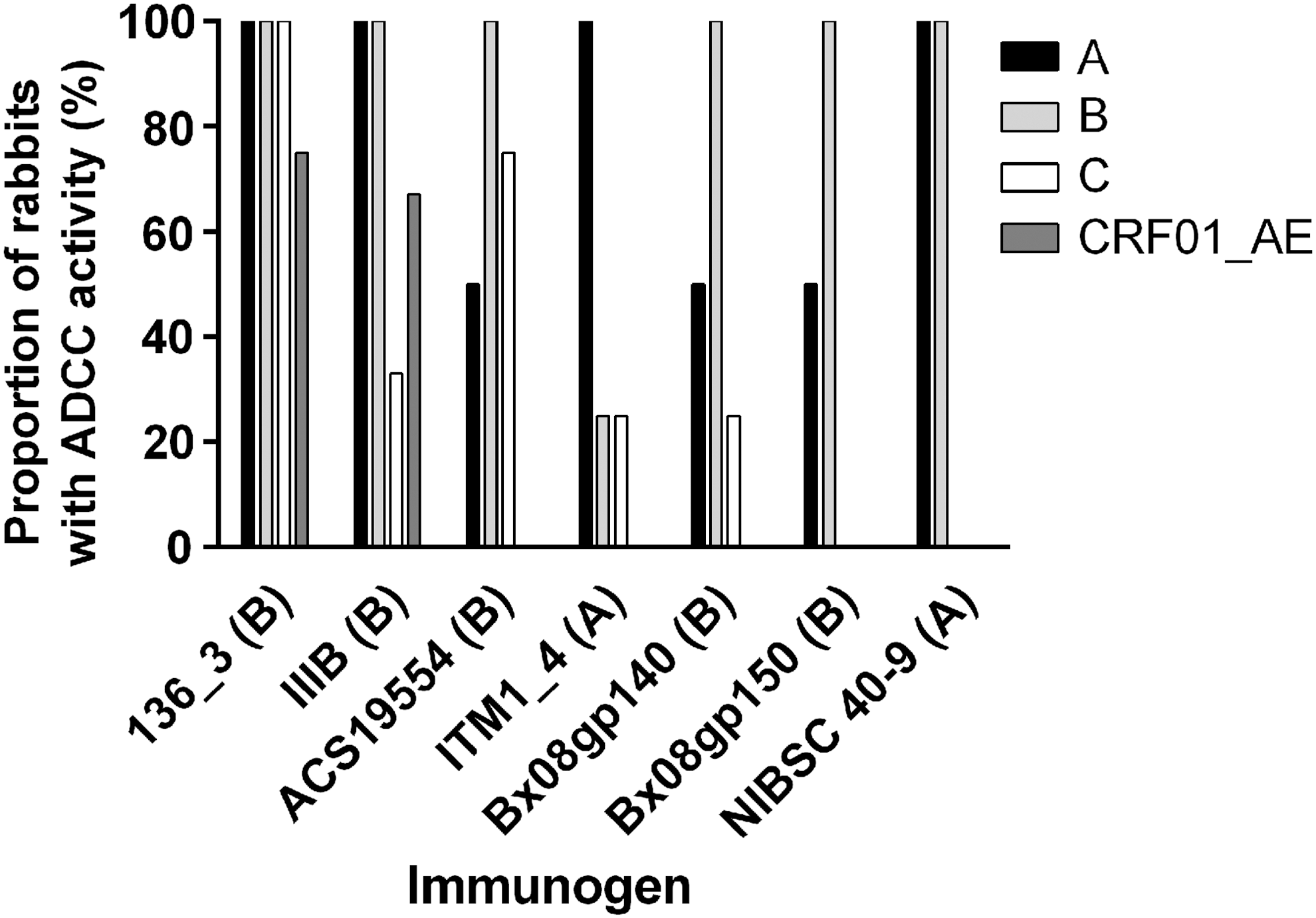
ADCC activity against different subtypes. The proportion of animals in which an ADCC response was detected. ADCC activity was determined 2 weeks after the last immunization, that is, week 14 or 16, and a preimmunization serum from the same animal was used as background. Seven groups of rabbits were immunized with trimeric 136_3 gp140, monomeric IIIB gp120, trimeric ACS19554 gp140, trimeric ITM1_4 gp140, trimeric Bx08 gp140 preceded by Bx08 gp150 DNA, trimeric Bx08 gp140 preceded by Bx08 gp140 DNA, and trimeric NIBSC 40-9 gp140 preceded by NIBSC 40-9 gp150 DNA. The sera were tested against envelopes of subtype A (UG37), subtype B (BaL), subtype C (ZA1197MB), and subtype CRF01_AE (CM243). The subtype of the immunogen is written in parentheses. CRF, circulating recombinant form.
Discussion
In this study, we identified immunogens that induced anti-HIV-1 antibodies with ADCC activity across different subtypes in a rabbit vaccination model. This is a unique study, not only identifying promising vaccine candidates but also demonstrating that it is possible to use rabbits as a small animal model when screening HIV-1 vaccine candidates for the induction of ADCC-mediating antibodies. The ADCC response was measured in a functional assay measuring the proteolytic activity of granzyme B after its delivery into target cells initiated by antibody recognition. Rabbit sera were tested, using human PBMCs as effector cells and HIV-1 envelope-coated CEM.NKR cells as target cells. It has previously been demonstrated that the results using coated target cells do not differ from when using infected target cells. 31 We confirmed that this was true also when using rabbit serum (data not shown). In this assay, the optimal dilution of rabbit sera for granzyme B activity was lower compared to what is normally observed for human sera in the same assay. 31,32 We cannot exclude the possibility that this is due to suboptimal recognition of the rabbit IgG by the human Fc receptors. However, we compared the use of human PBMCs with rabbit PBMCs, and this did not improve the outcome but we cannot exclude that this was influenced by the mismatch of combining rabbit effector cells with human target cells (Supplementary Fig. S2).
Most of the immunogens tested in this study originated from clinical HIV-1 isolates that had elicited broadly neutralizing antibodies in HIV-1 infected individuals. However, these individuals have not been screened for ADCC-mediating antibodies.
The seven immunogens inducing the most potent ADCC antibody responses were trimeric 136-3 gp140, monomeric IIIB gp120, trimeric ACS19554 gp140, trimeric ITM1_4 gp140, Bx08 gp140 DNA prime plus protein boost, Bx08 gp150 DNA prime plus protein boost, and NIBSC 40-9 gp150 DNA prime plus protein boost. 136-3 originates from a 3-month-old child infected with subtype B HIV-1 at birth. 33 IIIB is a commonly used laboratory subtype B strain of HIV-1. 34 ACS19554 originates from a progressing HIV-1 subtype B-infected adult 47 months postinfection. 35 ITM1_4 originates from a long-term survivor, an 11-year-old child infected with subtype A HIV-1 at birth. 36 Bx08 is a clade B primary isolate from an HIV-1-infected French individual 8 months post-seroconversion. 25,27,37 NIBSC 40_9 gp150 clade A was constructed from a Ugandan patient serum scoring high in neutralization titers and breadth. 38
These protein immunogens induced ADCC-mediating antibodies in rabbits when immunized with the adjuvant CAF01 or, in the case of IIIB, CAF02. CAF01 is a synthetic two-component liposomic adjuvant comprising the quaternary dimethyldioctadecylammonium ion (DDA) and trehalose 6,6′-dibehenate (TDB). 28 DDA facilitates antigen adsorption and presentation, while TDB potentiates the immune response through the Mincle receptor. 39 In CAF02, part of DDA was replaced with the neutral lipid of DSPC, which induces a more Th2-prone response. 29 Trimeric ITM1_4 gp140 was also tested without adjuvant, resulting in no measurable ADCC response (Table 1), demonstrating the importance of using a potent adjuvant. Moreover, ACS19554, 136-3, and ITM1_4 were all used as gp140 trimers. In fact, trimeric gp140 proteins in CAF01, with or without a preceding DNA prime, were overall the most compelling inducers of ADCC-mediating antibodies in this model.
However, the experiments were not set up in a way that enabled us to compare the same immunogen as a monomeric protein, a trimeric protein, DNA, virus-like particles, or a DNA prime followed by protein boost regimen. IIIB was immunized as monomeric gp120; however, here, it was used in combination with the adjuvant CAF02. We cannot exclude that the use of a different adjuvant enabled the effective ADCC response. Unfortunately, there was no comparison with the same immunogen using CAF01. The fact that trimeric gp140 is superior to monomeric gp120 with regard to the elicitation of binding titers, neutralizing antibodies, and ADCC-mediating antibodies has also been shown in other studies. 25,40
Three of the most potent inducers of ADCC-mediating antibodies, Bx08 gp140, ITM1_4 gp140, and NIBSC 40-9 gp150, also induced a high level of neutralizing antibodies against the HIV-1 subtype B strain SF162, as measured in a TZMbl assay using purified IgG. 25 Moreover, we observed an overall correlation between ADCC and neutralizing capacity when including all available data points from subtype B-derived immunogens. Nevertheless, this correlation was imperfect, and there were several instances in which the neutralization and ADCC response did not correspond with each other. The V1 V2 region of gp120 have previously been shown to be important for neutralization of SF162 41 ; the overall correlation we see when comparing naturalization and ADCC may therefore imply that V1 V2 specific antibodies may be important also for ADCC. There was also a general correlation between ADCC and HIV-1 envelope-specific IgG. A correlation between ADCC and neutralization has been observed in recent studies, 24,42 whereas earlier studies have shown the opposite, that is, a lack of correlation among neutralization, binding titers, and ADCC. 43 –46 This could suggest an incomplete overlap in the specificities of antibodies that mediate ADCC and neutralization.
Certain IgG isotypes favor ADCC (e.g., IgG1 and IgG3). The relative distribution of IgG isotypes in sera with same IgG antibody titer may be manipulated by different adjuvants and modes of immunization, therefore influencing the ADCC measured. Because rabbits only possess one IgG isotype, this is not an issue in the rabbit model.
Antibody responses cross-reactive between different HIV-1 subtypes and strains are essential for an effective vaccine. With this in mind, we tested the seven groups displaying the highest ADCC intra-subtype responses for cross-reactivity to the three other subtypes. The responses were highest against the subtype corresponding to the immunogen. Similarly, in a recent study of a primarily subtype B-infected cohort, ADCC responses were highest against subtype B. 14,47 The monomeric IIIB gp120 and trimeric 136-3 gp140 immunogens displayed the broadest ADCC response, with cross-reactivity to all of the four different clades tested. Three others, ACS19554, ITM-4, and Bx08 gp140, induced ADCC-mediating antibodies that were cross-reactive between three different clades, as well as neutralizing antibodies.
In conclusion, we present a rabbit vaccination model suitable for screening HIV-1 vaccine candidates for the induction of ADCC-mediating antibodies, as well as neutralizing antibodies. The immunogen inducing the most potent and broadest ADCC response was trimeric 136-3 gp140. Although we were able to identify vaccine candidates concurrently inducing both types of responses, ADCC-mediating antibodies did not necessarily correlate with neutralizing antibodies, highlighting the importance of using an assay measuring the desired feature.
Footnotes
Acknowledgments
The authors thank Birgit Knudsen for expert technical assistance. The project was sponsored by the European Community under EC-FP7-grant “next Generation HIV-1 Immunogens inducing broadly reactive neutralizing antibodies” NGIN_201433. Other members of the NGIN Consortium are Mauro Malnati and Lucia Loalco, Fondazione Centro San Raffaele del Monte Tabor, Milan, Italy; Hanneke Schuitemaker, AMC at the University of Amsterdam, Amsterdam, the Netherlands; Luigi Buonaguro, Istituto Nazionale Tumori “Fond. G. Pascale,” Naples, Italy; Philippe Saudan, Cytos Biotechnology AG, Zurich-Schlieren, Switzerland; Mario Clerici, Milano University Medical School, Milan, Italy; Anna-Lena Spetz and Francesca Chiodi, Karolinska Institut, Stockholm, Sweden; Jan Albert and Britta Wahren, SMI/MTC, Stockholm, Sweden; Guido Vanham, Institute of Tropical Medicine, Antwerp, Belgium; Meghna Ramaswamy, National Institute for Biological Standards & Control, Hertfordshire, United Kingdom; Sylvan Fleury and Anick Chalifour, Mymetics Corporation, Epalinges, Switzerland; Eva-Maria Fenyo and Marianne Jansson, Lund University, Lund, Sweden; and Morgane Bomsel, Institut Cochin, Paris, France.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.