
Abstract
HIV is the causative pathogen of AIDS, which has generated worldwide concern. Long noncoding RNAs (lncRNAs) are a rising star in virus–host cross-talk pathways; they are differentially expressed during many viral infections and are involved in multiple biological processes. Currently, lncRNA growth arrest-specific transcript 5 (GAS5) is known to be downregulated during HIV-1 infection. However, the functions and mechanisms of GAS5 in HIV-1 infection remain largely unknown. In this report, it was found for the first time that GAS5 could inhibit HIV-1 replication. Interestingly, using bioinformatics analyses (with Genomica and starBase.v2.0), GAS5 was found to potentially interact with miR-873. It was further verified that GAS5 could suppress miR-873. Moreover, miR-873 could promote HIV-1 replication. Together, these results not only suggest that GAS5 may inhibit HIV-1 replication through interaction with miR-873 but the results may also provide novel biomarkers for antiviral drugs or potential targets for future therapeutics for HIV/AIDS.
HIV
During HIV infection, lncRNA nuclear paraspeckle assembly transcript 1 (NEAT1) is upregulated. NEAT1 can attenuate HIV-1 Rev-dependent nuclear-cytoplasmic export. 10 LncRNA noncoding repressor of nuclear factor of activated T cells (NRON) is decreased by the early viral protein Nef, and intriguingly increased by the late viral protein Vpu. NRON can repress activity of viral long terminal repeat, 11 facilitating degradation of the viral transactivator protein Tat. 12 Furthermore, HIV-encoded antisense lncRNAs are also involved in modulating HIV gene expression by acting as epigenetic modulators. 13,14 Through these works, the functions of lncRNAs in HIV regulation are gradually coming to light.
Growth arrest-specific transcript 5 (GAS5) is a growth suppressor lncRNA that has crucial effects on tumorigenesis, 4,15 and often been taken as a novel diagnostic and prognostic biomarker for chemoradiotherapy. 16,17 Interestingly, it was reported that GAS5 can operate as a decoy for hepatitis virus C (HCV) NS3 protein to inhibit HCV replication. However, as a prelude since then, other virus infections regulated by GAS5 have not been discussed. Hence, the contributions of GAS5 in HIV-1 infection are still poorly understood, although this lncRNA was found to be downregulated in HIV-1 infection. 11
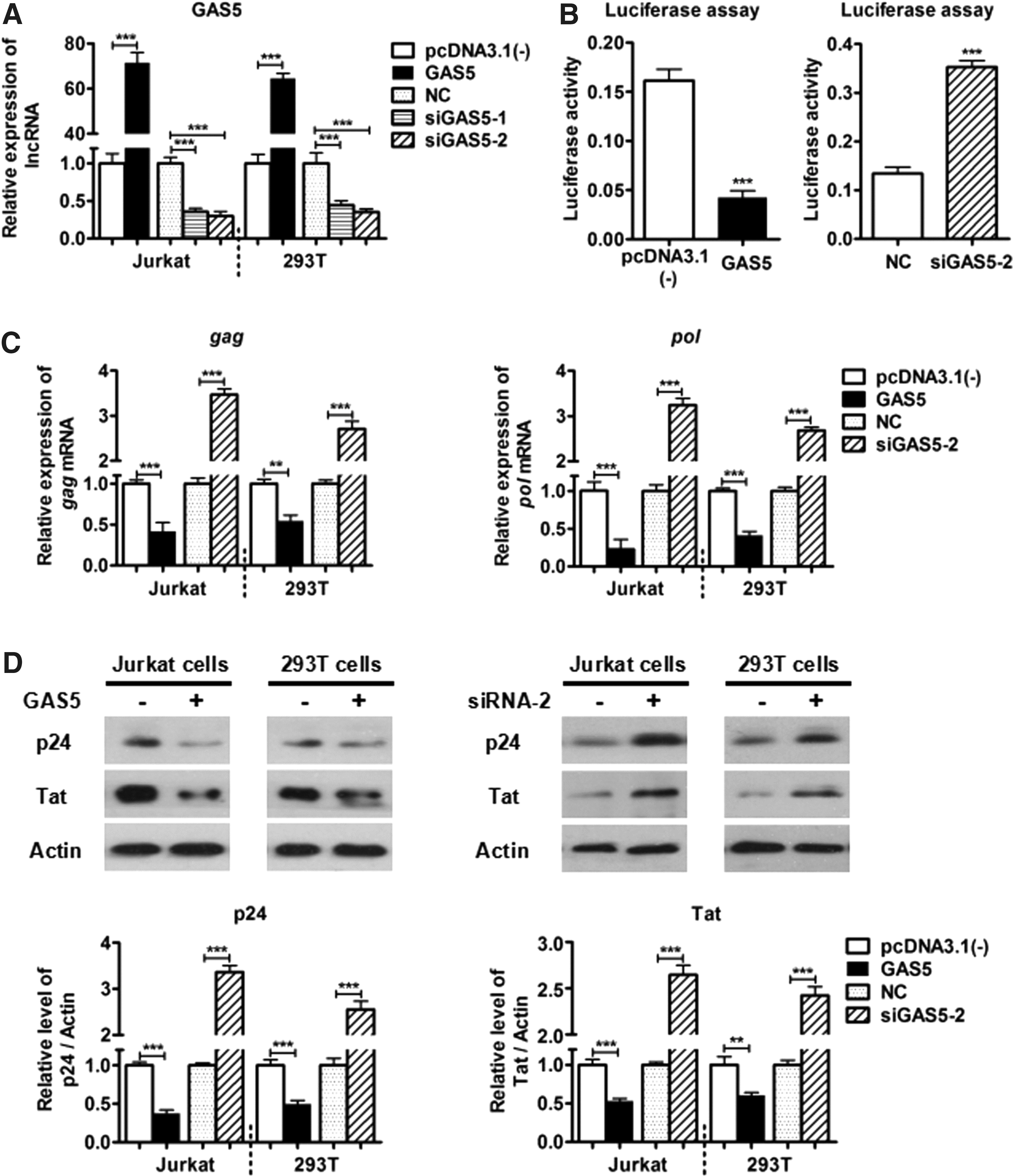
To preliminarily determine whether GAS5 is involved in HIV-1 replication, we first assessed the transfection efficiency of a GAS5 overexpression plasmid (gifts provided by Dr. Huang 4 ), in comparison with the pcDND3.1(-) vector in Jurkat and 293T cells. siGAS5-1: (5′-GGACCAGCUUAAUGGUUCUTT-3′) and siGAS5-2: (5′-GCAGACCUGUUAUCCUAAATT-3′) were also studied, in comparison with the negative control (NC) (5′-UUCUCCGAACGUGUCACGUTT-3′) (GenePharma, Shanghai, China). Lipofectamine 2000 (Invitrogen) was used for cotransfection. Jurkat and 293T cells were cultured in Roswell Park Memorial Institute-1640 (RPMI-1640)/Dulbecco's modified Eagle's medium, supplemented with 10% fetal bovine serum (HyClone), in a 37°C humidified atmosphere containing 5% CO2. After 48 h of transfection, total RNA was extracted from cells using the Trizol reagent (Invitrogen), and cDNA was obtained by the Reverse Transcription Kit (Thermo). Real-Time (RT)-quantitative polymerase chain reaction (qPCR) was performed with a SYBR green Real-Time PCR Master Mix Kit (Toyobo). These RT-qPCR results found GAS5 overexpression, by nearly 70-fold, in the GAS5-overexpressing group compared with the control group. The level of GAS5 was degraded ∼70% by GAS5-siRNA-1/2 compared with the NC group (Fig. 1A), and siGAS5-2 was selected for further experiments.
Inhibition of HIV-1 replication by GAS5.
Then, dual-luciferase reporter assays (Promega) were conducted to detect the efficacy of GAS5 on HIV-1NL4-3 by measuring the luciferase activity, normalized to Renilla activity. GAS5 or siGAS5-2, HIV-1 provirus plasmid pNL4-3.Luc.R-E-, and Renilla (pRL-TK; Promega) were cotransfected into 293T cells along with the helper plasmid pVSV-G at a weight ratio of 4:1. The cells were harvested 48 h post-transfection. The results showed that the luciferase activity of HIV-1NL4-3 was markedly decreased (by nearly 75%) after GAS5 overexpression compared with the control group. Conversely, siGAS5-2 promoted luciferase activity (by more than 2-fold) compared with the NC group (Fig. 1B).
To further confirm the function of GAS5, the mRNA levels of HIV-1NL4-3 gag and pol were determined by RT-qPCR in Jurkat and 293T cells. The primer sequences (kindly provided by Dr. Zhao) were as follows: gag forward primer, 5′-GGAAGTGACATAGCAGGAAC-3′, reverse primer, 5′-CATGCTGTCATCATTTCTTC-3′; pol forward primer, 5′-TTCCCTCAAATCACTCTTTG-3′, reverse primer, 5′-GGATGCGGTATTCCTAATT-3′; and Actin forward primer, 5′-CACGATGGAGGGGCCGGACTCATC-3′, reverse primer, 5′-TAAAGACCTCTATGCCAACACAGT-3′. We found that the overexpression of GAS5 could significantly reduce the mRNA levels of HIV-1NL4-3 gag and pol (by nearly 70%) compared with the control group. Conversely, siGAS5-2 had the opposing effect, increasing HIV-1 mRNA promotion (by nearly 3.5-fold) compared with the NC group (Fig. 1C).
In addition, HIV-1 protein levels were measured by Western blotting in Jurkat cells to verify GAS5 function with the employed transfection method. Cells were washed twice with ice-cold 1 × phosphate-buffered saline and lysed on ice with RIPA buffer (Beyotime) containing a 1 × protease inhibitor cocktail. Then, a BCA protein assay (Thermo) was used to quantify the protein lysate. The protein samples were resolved by 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Roche) by electroblotting. The following antibodies were used to perform the Western blotting analysis: anti-β-Actin (1:1,000 dilution; Abcam), anti-HIV-1 p24 (1:500 dilution; Fitzgerald), and anti-HIV-1 Tat (1:400 dilution; Abcam). The proteins were observed using an enhanced chemiluminescence system (Advansta) with a Kodak imager (Carestream Health) after incubating with 1:10,000-diluted horseradish peroxidase-conjugated secondary antibodies (Sungene Biotech) for 1.5 h at room temperature. The results revealed that the overexpression of GAS5 could inhibit the HIV-1 p24 and Tat proteins (by nearly 60%) compared with the control group. Furthermore, the siGAS5-2 had the opposite effect, promoting protein expression (by nearly 3-fold) compared with the NC group (Fig. 1D). Similar results were observed in 293T cells. All data are representative of three independent experiments with triplicate samples. All of the mentioned results demonstrate that GAS5 can inhibit HIV-1 replication.
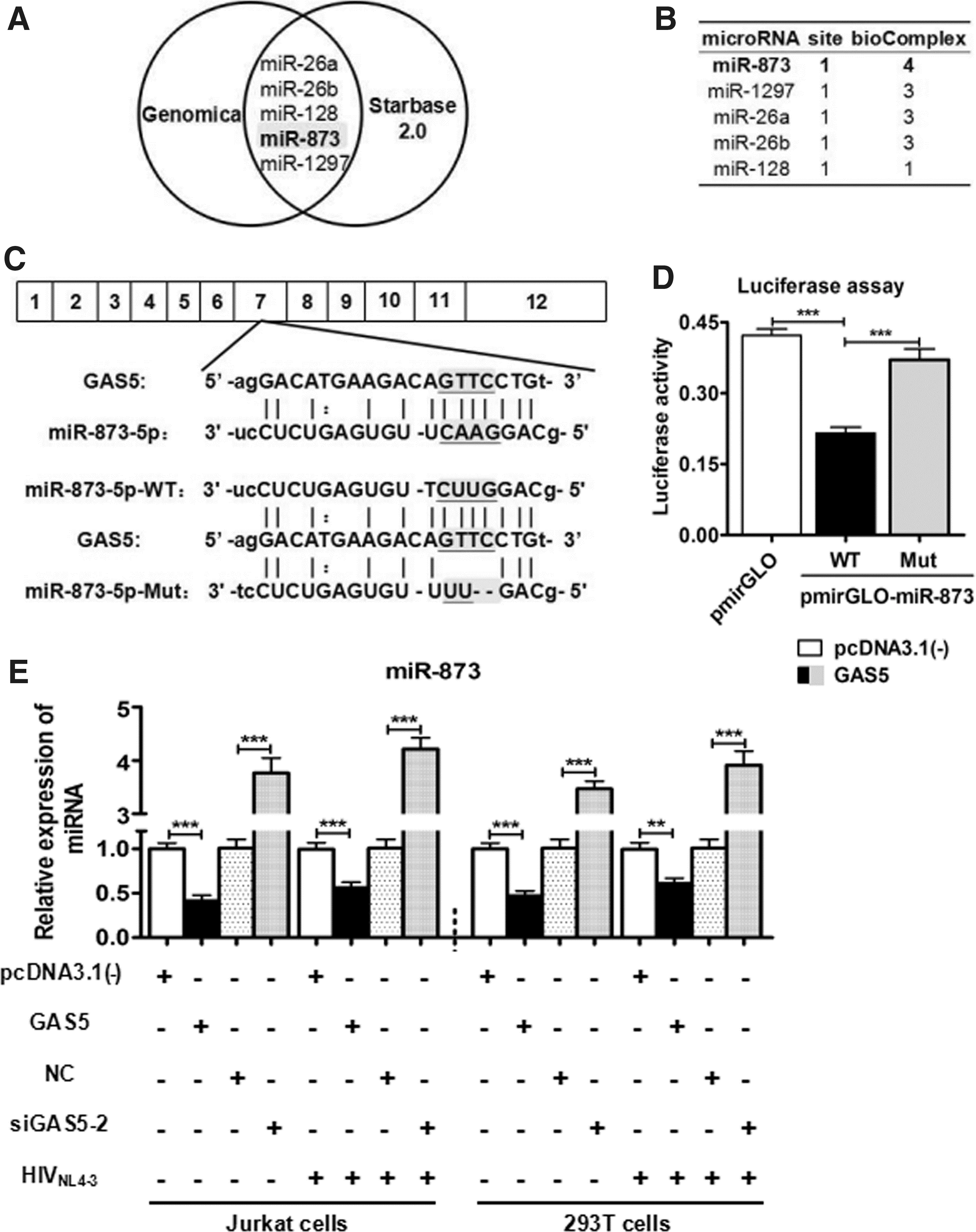
In HIV infection, many microRNAs (miRNAs) have been reported to target several HIV-1 host dependency factors that suppressed HIV-1 provirus, 18 or target antiviral proteins that stimulated HIV replication. 19 To further reveal the mechanisms of GAS5 in HIV-1 replication, bioinformatics software (Genomica and starBase.v2.0) were used to predict GAS5–miRNA interaction networks. Five miRNAs were found in both programs. MiR-873 was then chosen for further research due to its higher “bioComplex scores.” Competing endogenous RNA (ceRNA) is a vital part of the RNA–RNA interaction network, among which lncRNAs may serve as miRNA sponges by acting as ceRNAs. Previous studies have also confirmed that GAS5 could be a ceRNA, directly affecting the function of many miRNAs in tumors research. 20 –22 GAS5 (Fig. 2C) consists of 12 exons; the putative binding site of miR-873 is in exon 7# as calculated by the starBase.v2.0 program (Fig. 2C). The binding sequences were chemically synthesized and inserted into the pmirGLO-miR-873-WT or pmirGLO-miR-873-Mut vectors, and this was confirmed by sequencing. The results showed that the overexpression of GAS5 reduced pmirGLO-miR-873-WT activity (by nearly 40%) in 293T cells compared with the pmirGLO or the pmirGLO-miR-873-Mut groups (Fig. 2D). RT-qPCR assay further demonstrated that the overexpression of GAS5 suppressed miR-873 expression (by nearly 50%), compared with the control group. Knockdown of GAS5 increased miR-873 levels (by nearly 4-fold) compared with the NC group (Fig. 2E). Similar results were observed in HIVNL4-3 infection (Fig. 2E). Together, these results demonstrate that GAS5 can suppress miR-873. In addition, it has been reported that miR-873 can be repressed by lncRNA necrosis-related factor in cardiovascular diseases. 23
Effects of GAS5 on miR-873 levels.
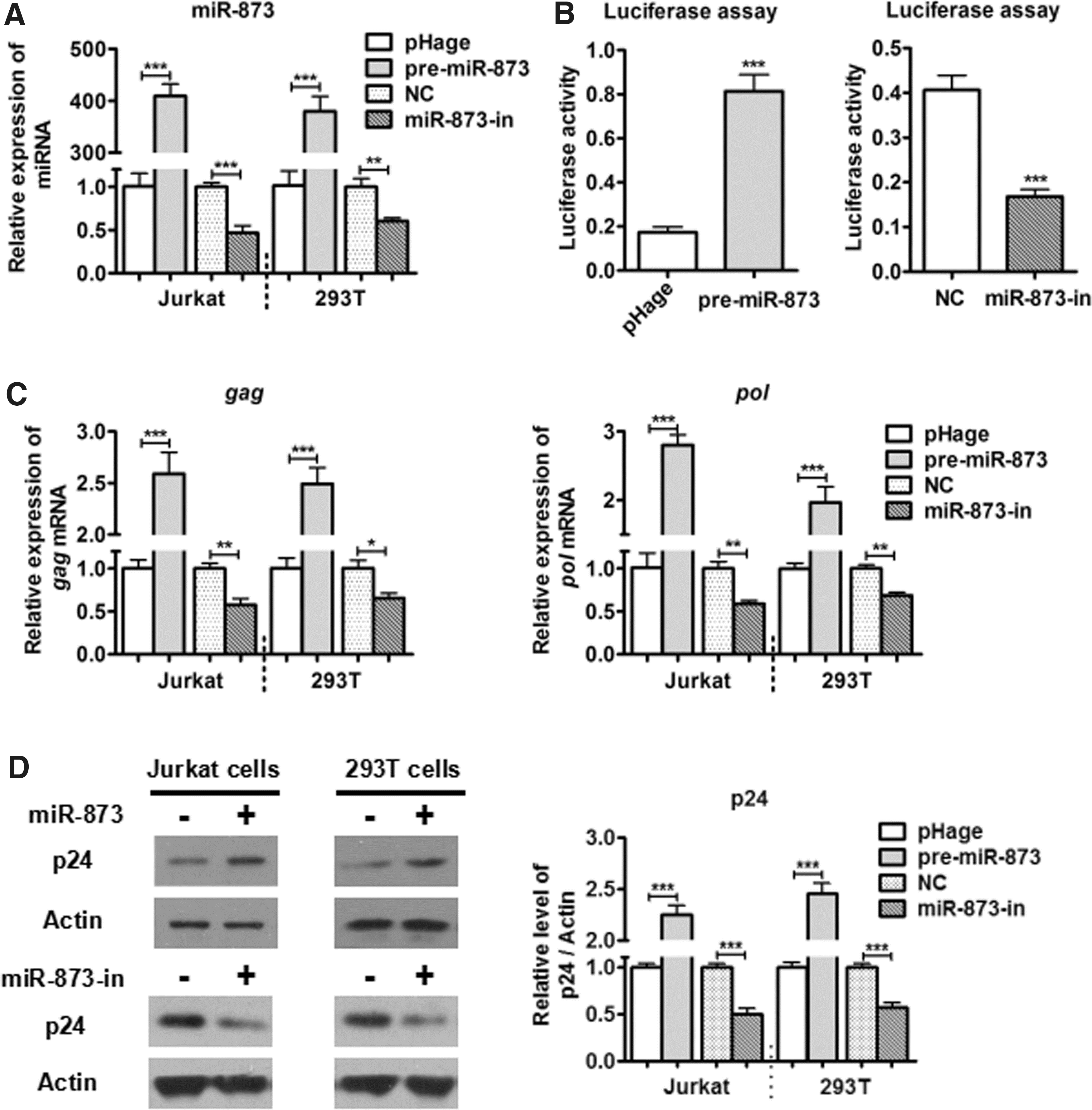
Given that GAS5 could inhibit HIV-1 replication and suppress miR-873, we speculated that miR-873 might have positive effects on HIV-1 replication. Using methods similar to what was already described, pHAGE-hsa-miR-873 or miR-873-Inhibitor (miR-873-in) (Ribobio) was cotransfected into Jurkat and 293T cells with pNL4-3.Luc.R-E- and the remainder of the reporter system. The results showed that miR-873 overexpression enhanced the miR-873 expression (by >400-fold) compared with the control group. Knockdown of miR-873 (by its inhibitor) would decrease the miR-873 expression (by nearly 50%) compared with the NC group (Fig. 3A). miR-873 could increase the luciferase activity of HIV-1NL4-3 (by almost 3-fold) in 293T cells compared with the control group (Fig. 3B). Moreover, miR-873 could facilitate the mRNA expressions of HIV-1NL4-3 gag and pol (by nearly 2.5-fold) by RT-qPCR, and promote p24 protein expression (by more than 2-fold) by Western blotting in Jurkat/293T cells, compared with the NC group (Fig. 3C, D). However, overexpression or inhibition of miR-873 could not affect the expression of Tat protein compared with the NC group (Supplementary Fig. S1; Supplementary Data are available online at
miR-873 promotes HIV-1 replication.
In summary, we identified that lncRNA GAS5 could attenuate HIV-1 replication and that it may act as a ceRNA, suppressing miR-873. These results propose a ceRNA mechanism in the immunopathogenic cross-talk of AIDS and could yield prospective biomarkers for anti-HIV treatment.
Footnotes
Acknowledgments
This work was supported by the National Natural Sciences Foundation of China (No. 81371790, 81641093, 81371422, 81571481, and 31170154), Major AIDS and Viral Hepatitis and Other Major Infectious Disease Prevention and Control project of China (No. 2014ZX10001003), the Fundamental Research Funds for the Central Universities of China, and the Translational Medical Research Fund of Wuhan University School of Medicine.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.