
Abstract
The main goal of this work was to identify molecular signatures in envelope surface glycoprotein that may be correlated with coreceptor usage by different human immunodeficiency virus (HIV)-2 strains. From inspection of aligned HIV-2 sequences, we verified that V1/V2 region showed the highest degree of amino acid sequence heterogeneity, including polymorphisms in N-linked glycosylation sites, sequence, and length. Furthermore, we did not find any correlation between the net charge and specific amino acid positions in V3 region with any particular coreceptor usage pattern. In conclusion, we showed that for HIV-2, the genetic determinants for coreceptor usage are distinct from those of HIV-1. More specifically, we did not identify any molecular signature, based on discrete amino acid positions either in V1/V2 or in V3 regions, which could be assigned to the preferential usage of a specific coreceptor.
H
Although several GPCRs have been shown to act as coreceptors for HIV in vitro, the role of alternative coreceptors, besides CCR5 and CXCR4, in HIV-1 infection is considered as being of limited importance in vitro and probably in vivo. However, some exceptions have been reported (reviewed in Ref. 4 ), suggesting the possibility that alternative molecules could have physiological relevance as cofactors for HIV-1 entry. The same is true concerning HIV-2 infection, wherein the more promiscuous usage of chemokine receptors as coreceptors has been widely referred in vitro. 5 In addition, the existence of primary isolates with atypical coreceptor usage profiles 6 –8 may further indicate that in HIV-2, the capacity to use other coreceptors besides CCR5 and CXCR4 should be potentially more common.
In HIV-1, the third variable (V3) region of Env SU glycoprotein seems to be the major molecular determinant of coreceptor usage: higher positive net charge (above +5) in V3 region of HIV-1, together with the presence of basic amino acids in positions 11 and/or 25 (referred to V3 sequence), has been associated with CXCR4 usage (or usage of both CXCR4 and CCR5). Accordingly, analysis of amino acid composition of the V3 region is used to predict coreceptor usage by HIV-1, namely in clinical practice using CCR5 inhibitors. 9
However, in HIV-2, functional studies aiming to decipher the contribution of envelope glycoproteins' regions to cellular receptors usage are much more scarce and in some aspects contradictory. Some studies associated the presence of particular amino acids in the V3 sequence with CCR5 or CXCR4 usage, whereas others have not found any relationship between them (reviewed in Ref. 5 ). More recently we have shown that the usage of CCR8 and the switch from CCR8 to CCR5 or to CCR5/CXCR4 dual tropic phenotype are determined by amino acids located in the base and tip of variable regions 1 and 2 (V1 and V2, respectively) of HIV-2 Env SU glycoprotein. 7
To further define the molecular determinants of coreceptor usage by HIV-2, we cloned, sequenced, and analyzed the region between the first and third conserved regions (C1–C3) of the env gene of six HIV-2 primary isolates, obtained from patients at different disease stages, showing a diversified spectrum of coreceptor usage (e.g., CCR5, CXCR4, and both CCR5/CXCR4), including viruses able to enter cells using alternative coreceptors such as CCR8.
The HIV-2 primary isolates were obtained from a cohort of infected individuals, the clinical and immunological data of which are summarized in Table 1. The infectivity assays regarding coreceptors usage were performed as described. 8 This set of HIV-2 primary isolates was analyzed by the ability to productively infect GHOST cells co-expressing CD4 and different coreceptors (CCR1, CCR2b, CCR3, CCR5, CXCR4, CCR8, GPR15, and CXCR6). Viral progeny production was measured by reverse transcriptase activity in culture supernatants and scored as semiquantitative results (Table 1). The coreceptors used by each viral isolate revealed a wide range of viral biotypes: R5 (HIV-2UCFL2069/MLC), X4 (HIV-2UCFL2068/MIL), R5X4 (HIV-2UCFL2061/MS), and multiple coreceptors, including CCR5 and CXCR4 (HIV-2UCFL2066/ETP, HIV-2UCFL2067/JAU, and HIV-2UCFL2071/TER).
Asympt = asymptomatic; sympt = symptomatic; clinical stage according to CDC classification is referred in parentheses.
Viral replication was assessed by peak RT activity measured in culture supernatants during a 21-day period after virus inoculation. −, peak RT activity <10 pg/ml; +, peak RT activity between 10 and 100 pg/ml; ++, peak RT activity between 101 and 1,000 pg/ml; +++, peak RT activity >1,000 pg/ml.
R5, usage of CCR5; X4, usage of CXCR4; R5X4, usage of both CCR5 and CXCR4 coreceptors; multiple CoR, usage of a broad range of coreceptors, including CCR5 and CXCR4.
RT, reverse transcriptase.
The Env coding region spanning from C1 to C3 of each virus was amplified by polymerase chain reaction as previously described.
10
Amplified fragments were cloned into the pCR4Blunt-TOPO vector (Invitrogen) and used to transform ultracompetent Escherichia coli DH5α-T1 cells (Invitrogen). Five of each env clones were sequenced and aligned using CLUSTAL X software. Owing to the similarity between clones, a consensus sequence was constructed for each HIV-2 strain, and their nucleotide and derived amino acid sequence were aligned with other HIV-2 and simian immunodeficiency virus (SIV) references from Los Alamos HIV sequence database (available online at
Since in HIV-1, the variable region (V3) of the Env SU glycoprotein has been referred as the main determinant for CCR5 and CXCR4 usage, we first analyzed the amino acid sequence of the V3 region of the cloned viruses. The alignment of the V3 region of envelope proteins revealed that these six isolates do not show any major divergence when compared with other HIV-2 strains (Fig. 1). From this alignment, it was also evident that in contrast to HIV-1, HIV-2 sequences corresponding to V3 loop showed limited variability. This emphasizes the notion that in HIV-2, the V3 region is probably less exposed and thus under minor selective pressure than its HIV-1 counterpart.
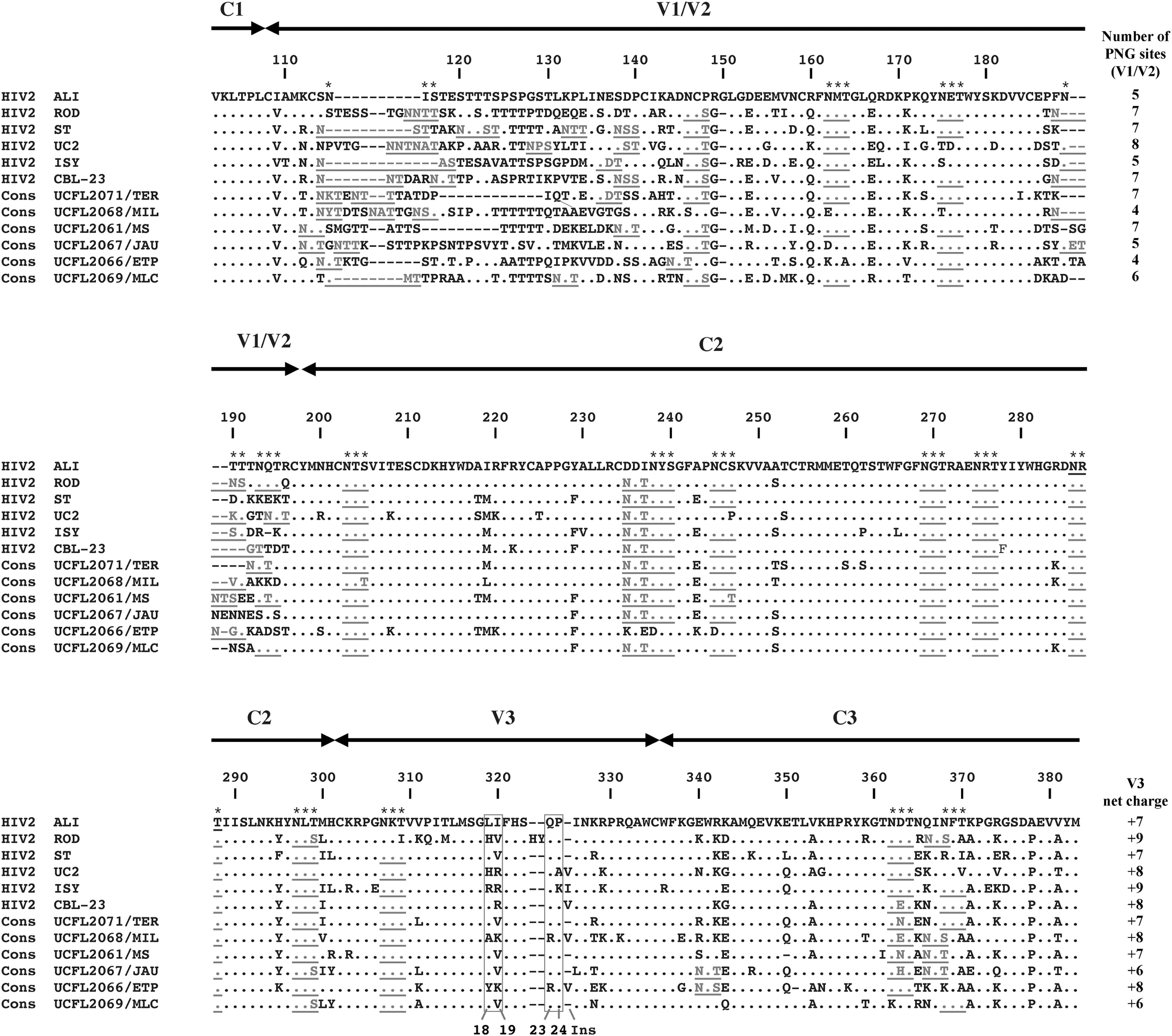
Characterization of the C1–C3 region from HIV-2 envelope glycoproteins. Amino acids are denoted by single-letter code. Amino acid numbering starts at the first amino acid after the signal peptide of HIV-2ALI strain (GenBank accession number: AF082339). The * above amino acids indicates the PNG sites of HIV-2ALI; the PNG sites of all the other HIV-2 strains are represented in gray and underlined. The gray boxes highlight the positions 18, 19, 23, and 24 of the V3 region (numbered according to the first cysteine residue); “Ins” represents the insertion after position 24 that is present in some V3 sequences. (−) represents a gap and (.) represents the same amino acid as shown in HIV-2ALI sequence. The number of PNG sites of the V1/V2 region and the global net charge of V3 are shown on the right. Accession numbers of the sequences included in this alignment are HIV-2ALI (AF082339), HIV-2ROD (M15390), HIV-2ST (M31113), HIV-2UC2 (U38293), HIV-2ISY (J04498), HIV-2CBL23 (U05352), HIV-2UCFL2066/ETP (GQ331132), HIV-2UCFL2067/JAU (GQ331133), HIV-2UCFL2068/MIL (GQ331134), HIV-2UCFL2069/MLC (GQ331135), HIV-2UCFL2061/MS (GQ331136), and HIV-2UCFL2071/TER (GQ331137). HIV, human immunodeficiency virus; PNG, putative N-linked glycosylation.
The study of putative N-linked glycosylation (PNG) sites (defined using N-Glycosite; available at
Analyzing the sequence alignment of HIV-2 viruses (Fig. 1), and considering the rules already mentioned, it seems particularly evident that no correlation/prediction of coreceptor usage could be established based on V3 sequences. The discrepancy between phenotypic data and amino acid composition of V3 alone emphasizes the notion that (1) in HIV-2, other regions of the Env glycoproteins are directly involved in coreceptor engagement; (2) cooperative interactions between different Env subunits must exist, and those are cell type and virus strain dependent.
In contrast to the V3 region, the variable regions 1 and 2 (V1/V2) of Env SU glycoprotein showed high degree of variability, in particular, in the V1 region (Fig. 1). This variability affects (1) number of PNG sites, (2) length, and (3) the net charge of V1/V2.
In general, the degree of glycosylation of HIV-2 Env is lower than that of HIV-1 and its influence on the use of coreceptors is not established. The analysis of HIV-2 sequences reveals that the number of PNG sites within the V1/V2 region varies between 4 (in HIV-2UCFL2068/MIL and HIV-2UCFL2066/ETP) and 7 (HIV-2UCFL2061/MS and HIV-2UCFL2071/TER), and did not correlate with coreceptor usage (Fig. 1). The contribution of PNG in HIV-2 tropism is scant, and as in HIV-1, the influence of discrete PNG is probably context dependent. Furthermore, the same mutations affecting PNG sites could have different effects in tropism, depending on the overall Env structure and the molecular mechanism modulating binding to cellular receptors. 13 Nevertheless, it seems that CXCR4 usage is more frequently observed in those strains with lower number of PNG sites within the V1/V2 region (HIV-2UCFL2068/MIL, HIV-2UCFL2067/JAU, and HIV-2UCFL2066/ETP, which have 4, 5, and 4 PNG sites, respectively), whereas those that do not use CXCR4 showed higher number of N-glycosylation sites (HIV-2UCFL2061/MS, HIV-2UCFL2071/TER, and HIV-2UCFL2069/MLC with 7, 7, and 6 PNG sites, respectively).
From inspection of V1/V2 sequences, it is very hard to find any particular molecular signature evocative of the coreceptor usage profile shown by HIV-2 strains included in this study. Similarly to HIV-1, the V1/V2 region of HIV-2 could be involved in mechanisms allowing viral escape from antibody-mediated neutralization. This explains the outstanding sequence heterogeneity detected in this region. However, in contrast to HIV-1, the V1/V2 region of HIV-2 seems also to be a major determinant of coreceptor usage. In fact, using a panel of isogenic mutant HIV-2 viruses, we recently demonstrated that the switch from CCR8 (R8) to CCR5 (R5) or to CCR5/CXCR4 (R5X4) dual tropic phenotype is determined by amino acids located in the base and tip of V1 and V2 loops; furthermore, it was also shown that several mutations are needed to convert an R8 isolate into an R5 or R5X4 variant. 7 This suggests that cooperating mutations must be simultaneously present within the V1/V2 regions to change the coreceptor engaged, precluding the use of simple rules based on the presence of a specific amino acid in a particular position to predict coreceptor usage.
In conclusion, and corroborating previous observations, 10 the characterization of Env SU glycoprotein of primary HIV-2 isolates revealed that (1) there is no V3 sequence “rule” (i.e., a specific amino acid in a particular position) that could be used to accurately predict coreceptor usage by HIV-2 isolates (e.g., CCR5 or CXCR4) and (2) although the V1/V2 region is involved in coreceptor engagement, there is no “molecular signature” based on discrete amino acid positions that could be assigned to the preferential usage of a specific coreceptor. Thus our results emphasize the notion that, in HIV-2, an accurate coreceptor usage predictive tool should be based on the overall sequence of V1, V2, and V3 regions, 7,14 rather than in rules aiming the identification of specific amino acids in a particular position of the V3 loop. 12
Footnotes
Acknowledgments
This work was supported by grants from Fundação para a Ciência e Tecnologia and Ministério da Saúde de Portugal (FCT grant: VIH/SAU/0006/2011).
Author Disclosure Statement
No competing financial interests exist.