
Abstract
Rectal transmission accounts for the majority of HIV transmissions among men who have sex with men (MSM). We previously demonstrated a distinct rectal mucosal immune environment in MSM engaging in condomless receptive anal intercourse (CRAI) compared with men who do not engage in anal intercourse, including enrichment of the microbiota for the Prevotellaceae family as compared with Bacteroidaceae. Here, we expand upon these previous findings to determine differences by anatomic site of collection (anal vs. rectal mucosa) and examine associations of the predominant taxa with other genera. We analyzed 16SrRNA gene sequences of the V1–V2 region generated on an Illumina MiSeq® from 35 MSM engaging in CRAI and 20 male controls. Observation by principal coordinates analysis and analysis of similarities test showed differing composition of the microbiota by anatomic site of collection. When analyzing the top 10 abundant genera from each anatomic site by generalized linear models, the predominant genera (Prevotella enrichment among MSM engaging in CRAI vs. Bacteroides among controls) were consistent; however, the two sites shared only four common genera. In addition, associations between the relative abundance of Prevotella and Bacteroides with other prevalent genera, by Spearman's rank correlations, were inconsistent when stratifying by study group. Prevotella versus Bacteroides predominant microbiota may not define a consistent underlying microbial community, and our data underline the importance of anatomic sampling site. Understanding the rectal mucosal immune environment, of which the microbiota is a critical component, will enable a better understanding of rectal HIV transmission.
M
Detailed methods on the clinical cohort were previously published. 3 In brief, 41 HIV negative MSM engaging in CRAI at least four times monthly and 21 controls underwent rectal mucosal sampling conducted at two study visits separated by a median of 9 weeks. Clinicians collected mucosal swabs for microbiota sequencing at two anatomic locations: the anus and through a rigid sigmoidoscope ∼8–10 cm from the anal verge (rectal mucosa). Swabs were stored in lysis buffer at −80°C; DNA was extracted and amplified using previously described methods. 3 Microbiota data generated by sequencing the V1–V2 region of the 16SrRNA gene on an Illumina MiSeq® instrument (rarefied to 2,215 sequences) were available from 54 participants for these analyses due to missing specimens and adequate sequence generation. The Greengenes database (version 13_8) was used to classify operational taxonomic units (OTUs) to the genus level. Sequences were deposited to the GEO database (accession GSE83245).
Alpha diversity was measured using Chao1 and Shannon indices and statistical significance was assessed with generalized linear models (GLMs) using RStudio® adjusting for age using the generalized estimated equation approach to account for repeated measurements. Beta diversity was measured using Bray–Curtis dissimilarity analysis and observed by principal coordinate analysis (PCoA). Differences in microbial composition between anatomic collection sites were evaluated using the analysis of similarities (ANOSIM) test. 6 The 10 most abundant genera in both anatomic sites were identified and compared with GLMs as already described. No differences were seen between visit 1 and visit 2, thus overall model-based means were reported. Spearman's rank correlation coefficients, adjusted for multiple correlations by the Holm–Bonferroni method, were calculated to evaluate the relationships between the OTUs of interest among MSM engaging in CRAI and controls for the rectal mucosa.
Microbiota sequences from the anus and the rectal mucosa were analyzed from 34 MSM engaging in CRAI and 20 controls. MSM engaging in CRAI were slightly older than controls (30 years vs. 27 years; p = .02); however, there were no differences in self-reported race between MSM engaging in CRAI and controls (78% white, 13% black, 9% other; p = .16). Alpha diversity indices (Shannon and Chao1) did not differ between anatomic sites of collection (p = .078; p = .38, respectively). Beta diversity differed between anatomic sites; the rectal mucosa and anus clustered separately in PCoA (Fig. 1A) and had statistically dissimilar microbial compositions (ANOSIM R = 0.1703, p = .001).
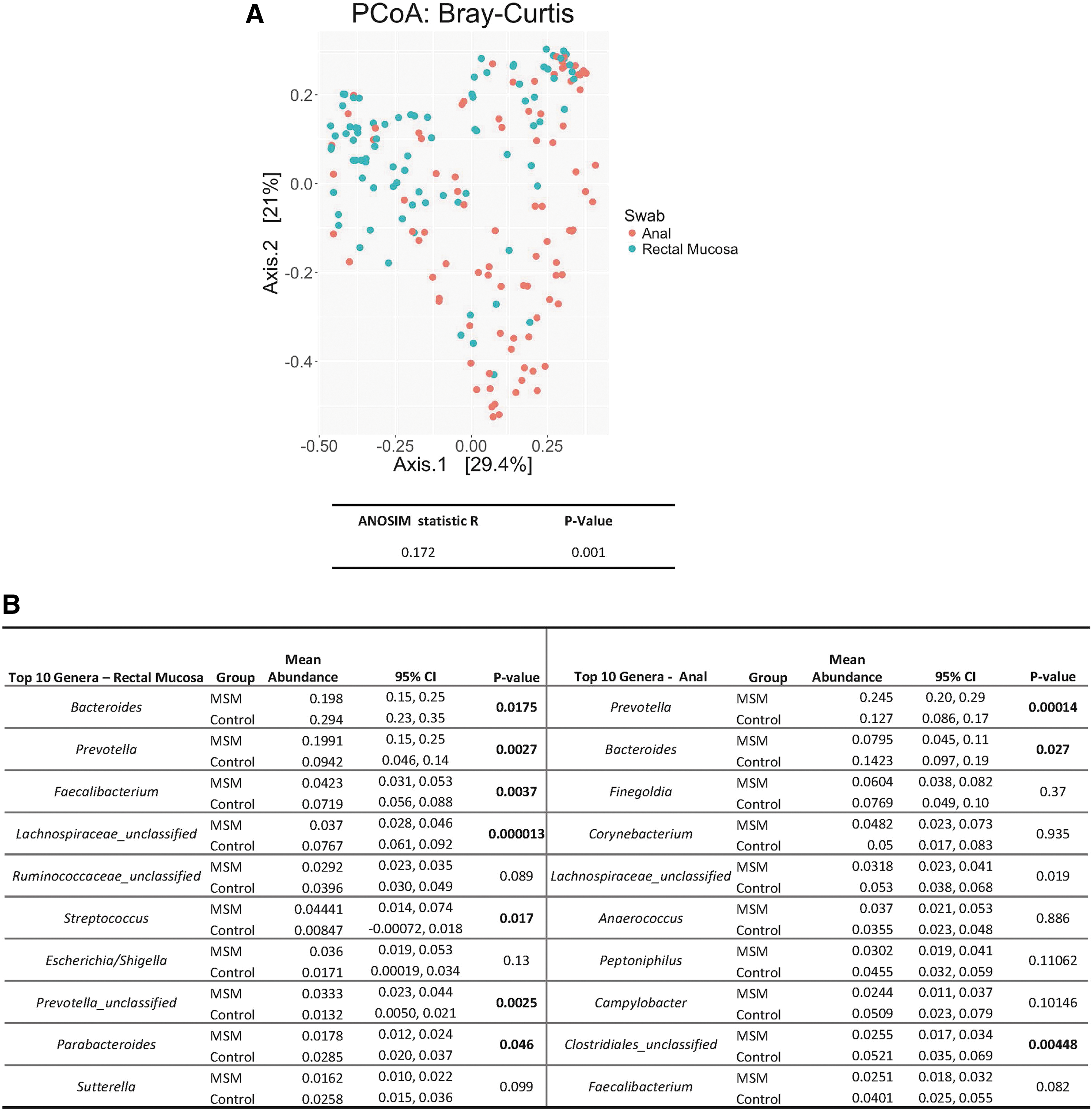
Bray–Curtis dissimilarity displayed by principal coordinate analysis shows anal and rectal mucosal swabs clustering independently, and ANOSIM results confirm statistical significance

The top 10 genera and relative abundances from each anatomic site are listed in Figure 1B. Although there were prominent similarities between the most abundant genera from both anatomic sites (Prevotella and Bacteroides), only the top four genera from the rectal mucosa were present in the anus. Not surprisingly, Corynebacterium, a component of normal skin flora, was the fourth most prevalent genus from the anus but was absent in the rectal mucosa. 7 In addition, taxa with pathogenic potential such as Streptococcus and Escherichia/Shigella were present only in the rectal mucosa. 7,8
The microbiota of MSM engaging in CRAI was enriched for taxa from the Prevotella genus in both the rectal mucosa and anus, whereas controls were enriched for the Bacteroides genus at both sites (Fig. 1B). Of interest, MSM engaging in CRAI exhibited a statistically significant increase in an unclassified genus also belonging to the Prevotellaceae family and the pathogenic genera Streptococcus, compared with controls in the rectal mucosa.
We examined associations between the relative abundance of Prevotella and Bacteroides and other genera stratified by study group (Fig. 1C, D) in the rectal mucosa. Both study groups showed a statistically significant negative association between Prevotella and Bacteroides. In rectal mucosa, Prevotella was positively correlated with an unclassified genus from the Prevotellaceae family in both study groups. Interestingly, a significant negative association between Prevotella and both Lachnospiraceae and Faecalibacterium was seen among controls, whereas a positive association was seen among MSM engaging in CRAI. Finally, Prevotella was negatively associated with genera containing pathogenic bacteria such as Escherichia/Shigella and Streptococcus among MSM but not among controls.
In this analysis, we have expanded upon our previously published data that showed rectal mucosal enrichment for Prevotellaceae among MSM engaging in CRAI as contrasted to Bacteroidaceae among controls. 3 Here, we compared the microbiota composition at the genus level between specimens collected at the anus and rectal mucosa and demonstrated that although the predominant genera (Prevotella vs. Bacteroides) were consistent, significant differences were apparent in the top OTUs between the anatomic sites. Previous studies have determined that rectal swabs, fecal samples, and rectal biopsies offer different perspectives of the gut microbiota, and our data support these conclusions. 9,10 In our study, taxa commonly associated with skin flora 7,11 were more prevalent in the anus, and enrichment for a second genus of Prevotella and pathogenic taxa among MSM engaging in CRAI was only detected in the rectal mucosa. The finding of two distinct Prevotella genera also suggests that a single species does not account for the enrichment seen among MSM engaging in CRAI.
Enrichment for Prevotella or Bacteroides in the rectal mucosa was inconsistently associated with other genera when stratifying by study group and was not consistent with previously reported associations from stool specimens. 12 In addition, although MSM engaging in CRAI were enriched for the pathogenic genera Streptococcus and Escherichia/Shigella, this was negatively associated with the relative abundance of Prevotella. This suggests that enrichment for either Prevotella or Bacteroides may not represent an underlying microbial community as previously suggested 13 and/or that CRAI may operate to influence other taxa independent of its effect on the relative abundance of Prevotella.
This study is limited by its modest sample size and potential confounders such as diet and medications. We were unable to fully characterize the specific species of Prevotella contributing to the enrichment seen among MSM, which will be the goal of future research. Additional research including ex vivo and in vitro experiments will be needed to fully understand the interaction of microbial taxa in the rectal mucosa and implications for mucosal immunity.
Understanding the rectal mucosal immune environment, of which the microbiota is a critical component, will enable a better understanding of rectal HIV transmission and the development of effective biomedical HIV prevention interventions. Based on our data, researchers must carefully ensure that the most relevant anatomic sites are sampled in microbiome investigations and interpret results beyond the predominant genera with caution.
Footnotes
Acknowledgments
We would like to thank the Emory Center for AIDS Research Clinical Virology Core laboratory for its contributions in generating the 16SrRNA sequencing data. This work was supported by the National Institutes of Health: K23AI108335, and a supplement grant to the Emory Center for AIDS Research P30 AI050409.
Author Disclosure Statement
No competing financial interests exist.