
Abstract
The analysis of patient derived HIV neutralizing antibodies (nAbs) and their target epitopes in the viral envelope (Env) protein provides important basic information for vaccine design. In this study we optimized an epitope, EC26-2A4, that is targeted by neutralizing antibodies from an elite controller (EC26) and localizes in the membrane-proximal external region from the gp41 transmembrane protein. Due to its overlap with the epitope of the first generation broadly neutralizing monoclonal Ab (mAb) 2F5 associated with autoreactivity, we first defined the minimal core epitope reacting with antibodies from EC26 plasma, but not with mAb 2F5. The optimized minimal epitope, EC26-2A4ΔM, was able to induce neutralizing antibodies in vaccinated mice. We further analyzed the frequency of antibodies against the EC26-2A4ΔM peptide in HIV-positive patient sera from a treated cohort and an untreated long-term nonprogressor (LTNP) cohort. Interestingly, 27% of the LTNP sera reacted with the peptide, whereas only 9% showed reactivity in the treated cohort. Although there was no association between the presence of antibodies against the EC26-2A4ΔM epitope and viral load or CD4 count in these patients, the CD4 nadir in the treated cohort was higher in patients positive for EC26-2A4ΔM antibodies, in particular in patients having such antibodies at an early and a late timepoint after infection.
Introduction
N
Based on these studies, it was recognized that bnAbs arising in patients essentially target epitopes localizing to six major regions of the native trimeric Env spike. These are in the outer envelope glycoprotein gp120 (1) the CD4 binding site, (2) glycan peptides in the V1V2 trimer apex, (3) high mannose patches in the V3 region, in the transmembrane protein gp41, (4) the membrane-proximal external region (MPER), (5) the fusion peptide and in the gp120-gp41 interface, (6) several conformation-dependent epitopes (reviewed in Ref. 25 ). These studies helped to design soluble mimics of trimeric Env spikes, which were extensively analyzed in recent years in vaccination studies. 26
However, despite continuous improvements, such gp140 immunogens were not yet able to induce bnAbs against circulating heterologous primary HIV-1 strains, but at best against autologous strains. 26 –28 Furthermore newer versions of these immunogens (SOSIP.664) do not contain the complete highly conserved hydrophobic gp41 MPER, which is targeted by a number of Abs, both with broad and less broad neutralization potential. 29 For this reason, the MPER is often considered separately from the native Env spike in structural and vaccination studies. 30
The MPER is the most C-terminal region of the exterior part of gp41 (Fig. 1) and only partially accessible in the native Env spike. 31 However, this flexible region is accessible for bnAbs during membrane fusion, that is, during the conformational transitions induced in the native Env spike upon binding to CD4 and coreceptors. 32 –34 Vice versa, amino acids in the MPER affect the overall conformation of the envelope trimers, 35 and antibodies targeting the MPER can influence directly or indirectly—through the induction of escape mutations—the neutralization sensitivity of the virus. 36

Functional domains in gp41, including epitopes for neutralizing antibodies in the MPER. Functional domains include the FP, the NHR, and the CHR forming the six-helix bundle hairpin structure, the MPER containing most gp41 epitopes for neutralizing antibodies, the TM domain, and the ICT. Numbering including the MPER sequence refers to HIV-1HXB2 Env amino acids. MPER epitopes associated with neutralizing antibodies are indicated by horizontal lines. 40 –46,76,77 CHR, C-heptad repeat; FP, fusion peptide; ICT, intracellular tail; MPER, membrane-proximal external region; NHR, N-heptad repeat; TM, transmembrane.
Studies mapping broad neutralizing activity in patient sera showed an association of MPER-specific antibodies with neutralizing activity of the sera. 37 –40 Furthermore, several linear and partially overlapping epitopes for bnAbs are localized in the MPER (Fig. 1), including two of the first generation bnAbs, 2F5 and 4E10. 41 –43 The most potent bnAb targeting this region is 10E8, which recognizes an epitope very close to the transmembrane region and partially buried by the lipid membrane of the virus. 31,44 –46 The frequent association of MPER epitopes with lipid components in the membrane 47 often results in autoreactivity of the corresponding mAbs. 48 This autoreactivity may also cause elimination of some of these mAbs by tolerance mechanisms during B cell ontology 49,50 or in the periphery. 51
We recently identified an epitope in the gp41 MPER, EC26-2A4, which is targeted by neutralizing antibodies from plasma of an HIV elite controller using an Env-tailored phage display library. 40,52 Although this epitope overlaps with that of the autoreactive mAb 2F5, antibodies purified from patient's plasma with the EC26-2A4 epitope did not show autoreactivity with cardiolipin. 40 To evaluate the EC26-2A4 epitope further in view of vaccination purposes, in this study we first determined the EC26-2A4 core epitope to define the essential contact residues of the EC26-2A4 purified plasma antibodies with the aim to maintain the potential to induce neutralizing antibodies upon vaccination, while minimizing the risk of inducing autoreactive 2F5-like antibodies.
We further analyzed the presence of antibodies against the optimized epitope in patient sera from two cohorts, a treated cohort from the HIV Center of the Goethe University in Frankfurt and an untreated HIV-positive long-term nonprogressor (LTNP) cohort from ANRS (Agence Nationale de Recherche sur le SIDA et les Hepatites Virales; ALT/CO15) in France to analyze potential associations of the presence of epitope-specific antibodies with clinical parameters.
Materials and Methods
Patient samples and ethics statement
This study was performed as retrospective subanalysis of the ongoing Frankfurt HIV Cohort Study, for which patient data and serum samples are being collected prospectively since 1987. 53 It was approved by the data protection officer of the state of Hesse and the Frankfurt University Ethics Committee (Umbrella-Vote-No. 270/09), including special subproject Approval No. 3/HIVERA (February 17, 2014). Patients gave written informed consent to be admitted since 2009. All cohort individuals were treated in concordance with the Declaration of Helsinki, national and institutional standards. Samples from 979 patients were included in this study.
This study also enrolled 71 LTNP HIV-1+ patients from the French ALT cohort (ANRS CO15). As previously reported, ALT cohort members met the following inclusion criteria: HIV seropositivity for at least 8 years and CD4 cell counts greater than 600/mm3 for the past 5 years, whatever the viral load, but without symptoms or antiretroviral therapy. 54,55 The CO15 ALT cohort is funded and sponsored by ANRS and was approved by the Ethics Review Committee of Ile de France-VI (Paris, France). All LTNP patients provided written informed consent, and all methods were performed in accordance with relevant guidelines and regulations indicated by the Declaration of Helsinki.
Epitope mapping by spot synthesis peptide arrays
Peptide arrays were synthesized by Fmoc chemistry at activated PEG spacers on cellulose membranes by automated parallel peptide synthesis on a MultiPep RS instrument (Intavis) according to the manufacturer's instructions. 56,57 After rehydration, washing, and blocking with 5% (w/v) skimmed milk powder in PBS (MPBST) supplemented with 0.05% Tween-20 for 3 h at room temperature (RT), membranes were incubated with 500 ng/ml 2% MPBST diluted affinity-purified plasma antibodies or mAb 2F5 at 4°C overnight. After washing with PBST, bound antibodies were detected with an HRP-conjugated α-human IgG antibody (1 h, RT) and visualized using an ECL Kit (Thermo Fisher Scientific).
Affinity purification of EC26-2A4ΔM specific plasma antibodies
One milligram Dynabeads M-280 Streptavidin (Thermo Fisher Scientific) was incubated with 2 μg biotinylated EC26-2A4ΔM peptide in 500 μl PBS for 90 min at RT. Unbound EC26ΔM peptide was removed by three washing steps with 1 ml PBST (PBS/0.05% Tween-20). Subsequently, 1–1.5 ml heat inactivated patient serum (56°C, 1 h) was incubated with the beads overnight at 4°C on a rotator. After five washing steps with PBST, bound epitope specific antibodies were eluted with 300 μl elution buffer (Pierce™ IgG Elution Buffer) for 30 min at RT on a rotator. Finally, eluted antibodies were neutralized by adding 30 μl 1 M Tris-HCl pH 9.0. Antibody concentrations were determined by NanoDrop.
ELISA for the detection of epitope-specific antibodies in patient or mouse sera
Microtiter plates (MICROLON High Binding; Greiner) were coated with 100 ng/well EC26-2A4 or 3S peptide in PBS in duplicates at 4°C overnight. For the biotinylated EC26-2A4ΔM peptide, we used 10 ng/well and precoated streptavidin plates (200 ng/well). After three washing steps with PBST (PBS/0.05% Tween-20), plates were blocked with 5% (w/v) skimmed MPBST for 2 h at RT and subsequently incubated with patient plasma (diluted 1:100) or mouse sera (1:100–1:312,500 in fivefold serial dilutions) for 2 h at RT. Plates were washed thrice with PBST and incubated with HRP-conjugated anti-human or anti-mouse IgG antibodies, respectively, for 1 h at RT. After five washing steps, the plates were developed using 100 μl/well SureBlue TMB substrate (KPL, Inc.), stopped with 100 μl 1 N HCl, and read at 450 and 650 nm as a reference.
Analysis of EC26-2A4ΔM-specific antibodies for autoreactivity by ELISA
Purified epitope-specific plasma antibodies were tested for autoreactivity against cardiolipin and phosphatidylserine by ELISA using Anti-Cardiolipin IgG Test and Anti-Phosphatidylserine IgG Test (Bio-Rad) according to the manufacturer's instructions. Monoclonal neutralizing antibody controls and patient derived EC26-2A4ΔM epitope-specific plasma antibodies were tested at 10 μg/ml.
Mouse immunization
Six-week-old NMRI mice (Charles River) were primed with 50 μg codon optimized HIV-1JRFL DNA intradermally and after 4 weeks boosted three times every 4 weeks, each with 50 μg EC26-2A4ΔM peptide coupled to KLH (Peptides & Elephants, Germany) in monophosphoryl lipid adjuvant. Control mice received only KLH. Five mice were analyzed per group. Blood samples were collected 2 weeks after the last boost and before DNA priming. Mouse sera were heat inactivated at 56°C for 30 min and analyzed for epitope specific binding antibodies by ELISA and for HIV neutralizing antibodies in the TZM-bl assay. All animal experiments were performed according to the institutional guidelines.
Neutralization studies
Neutralization studies were performed with HIV-1 pseudoviruses in the standardized TZM-bl assay as previously described. 58,59 All serum samples were heat inactivated at 56°C for 1 h before use. Briefly, pseudoviruses were produced by cotransfection of 293T cells with an env expressing vector and an env-deficient backbone vector (pSG3Δenv) and titered on TZM-bl cells. Pseudovirus stocks [15,000 relative luminescence units (RLU)] were preincubated with serial dilutions of serum or purified antibodies for 1 h at 37°C followed by addition of 1 × 104 TZM-bl cells per well of a 96-well plate for 48 h in duplicates. After cell lysis, RLU were determined with a luminometer (BMG Labtech GmbH). The 50% inhibitory dose (IC50) was defined as the reciprocal serum dilution causing 50% reduction in RLU compared to virus control wells.
Results
Optimization of the EC26-2A4 core epitope for differential binding to neutralizing antibodies from plasma EC26 and autoreactive mAb 2F5
In previous work we have narrowed down the original 29 amino acids (aa) long EC26-2A4 epitope to an 8 aa core epitope EC26-2A4MZ with the sequence 659ELLELDKW666, which is essential for binding of epitope-specific neutralizing antibodies purified from EC26 plasma. 40 As mAb 2F5 with an epitope overlapping the EC26-2A4MZ core epitope (Fig. 1) is associated with autoreactivity against phospholipids like cardiolipin, in this study we optimized the EC26-2A4MZ peptide sequence to achieve reactivity with EC26 antibodies, but not with mAb 2F5. We first performed an alanine walk through the EC26-2A4MZ peptide sequence to identify essential residues important for binding EC26 plasma derived antibodies. This alanine walk identified L660, L663, D664, and K665 as crucial for binding, with only minor contributions of E659 and W666 (Fig. 2A).
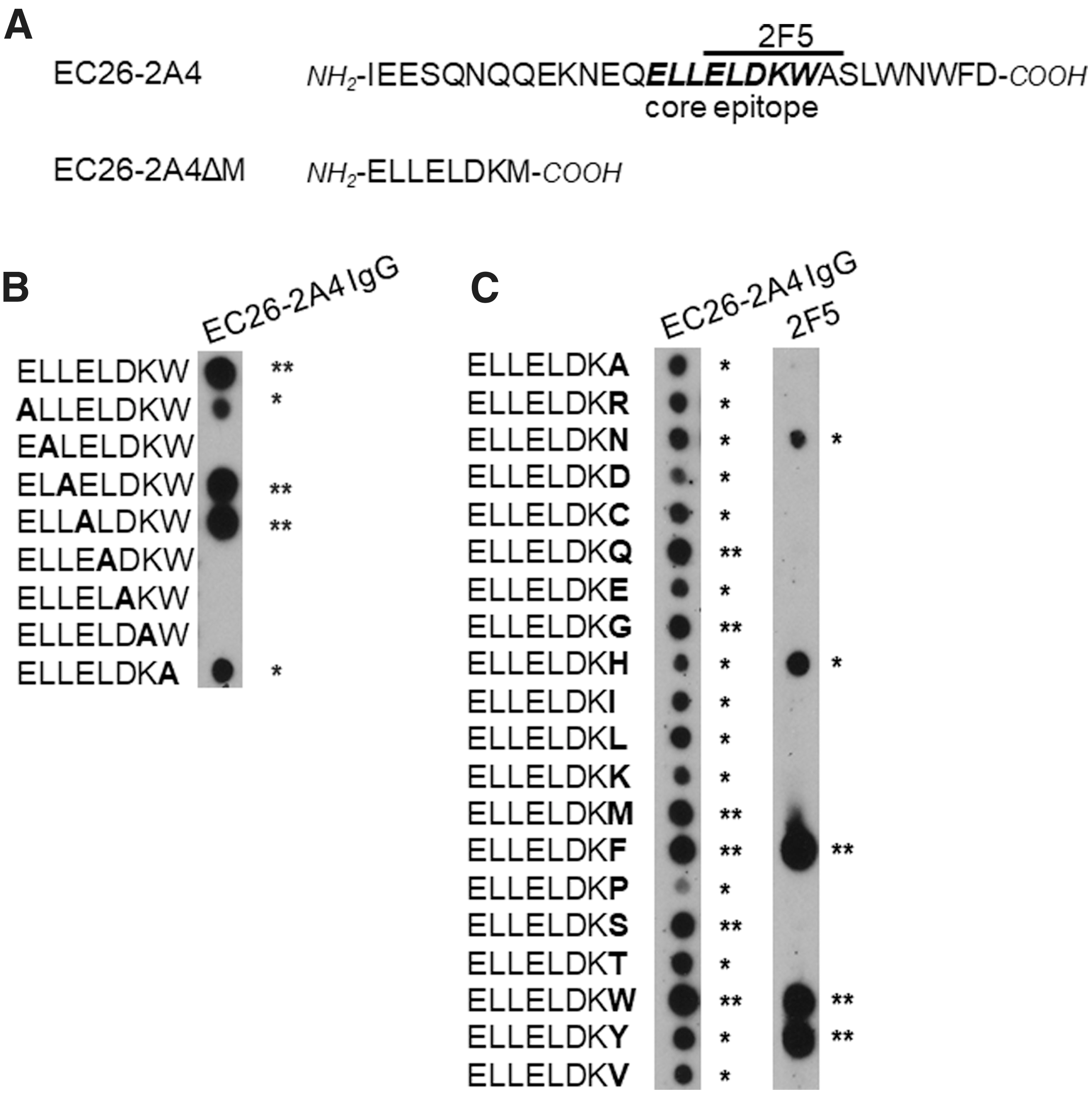
Optimization of the EC26-2A4 core epitope for differential reactivity with EC26 plasma antibodies and mAb 2F5 on peptide arrays.
In contrast, W666 as aromatic amino acid is absolutely essential for the reactivity with autoreactive mAb 2F5 and is known to mediate contact to the plasma membrane juxtaposed to the 2F5 MPER epitope and therefore likely to be involved in autoreactivity. We therefore exchanged W666 in the EC26-2A4MZ core for all other 19 amino acids to identify peptides with differential binding to epitope-purified EC26-2A4 antibodies and mAb 2F5 (Fig. 2B). Among four peptides showing strong binding to EC26-2A4 antibodies and no binding to mAb 2F5 (Fig. 2C, last amino acid position Q, G, M, or S), peptide EC26-2A4ΔM with the sequence ELLELDKM was analyzed further, as it consistently showed the observed reactivity pattern.
EC26-2A4ΔM induces HIV-neutralizing antibodies upon immunization of mice
Mice were vaccinated in an env-DNA prime/peptide boost approach with the EC26-2A4ΔM and the original EC26-2A4 peptide coupled to KLH. Compared to the preimmune sera used as control, both peptides generated a strong peptide-specific antibody response in all immunized mice, as measured by ELISA; for two of the five mice immunized with the short EC26-2A4ΔM peptide, titers even reached levels comparable to those of the mice immunized with the long peptide EC26-2A4 (Fig. 3A). Furthermore, sera from mice immunized with the optimized peptide EC26-2A4ΔM neutralized all three heterologous tier 1 HIV-1 strains analyzed with comparable IC50 titers as obtained with the original longer epitope peptide EC26-2A4 (Fig. 3B and Ref. 52 ).
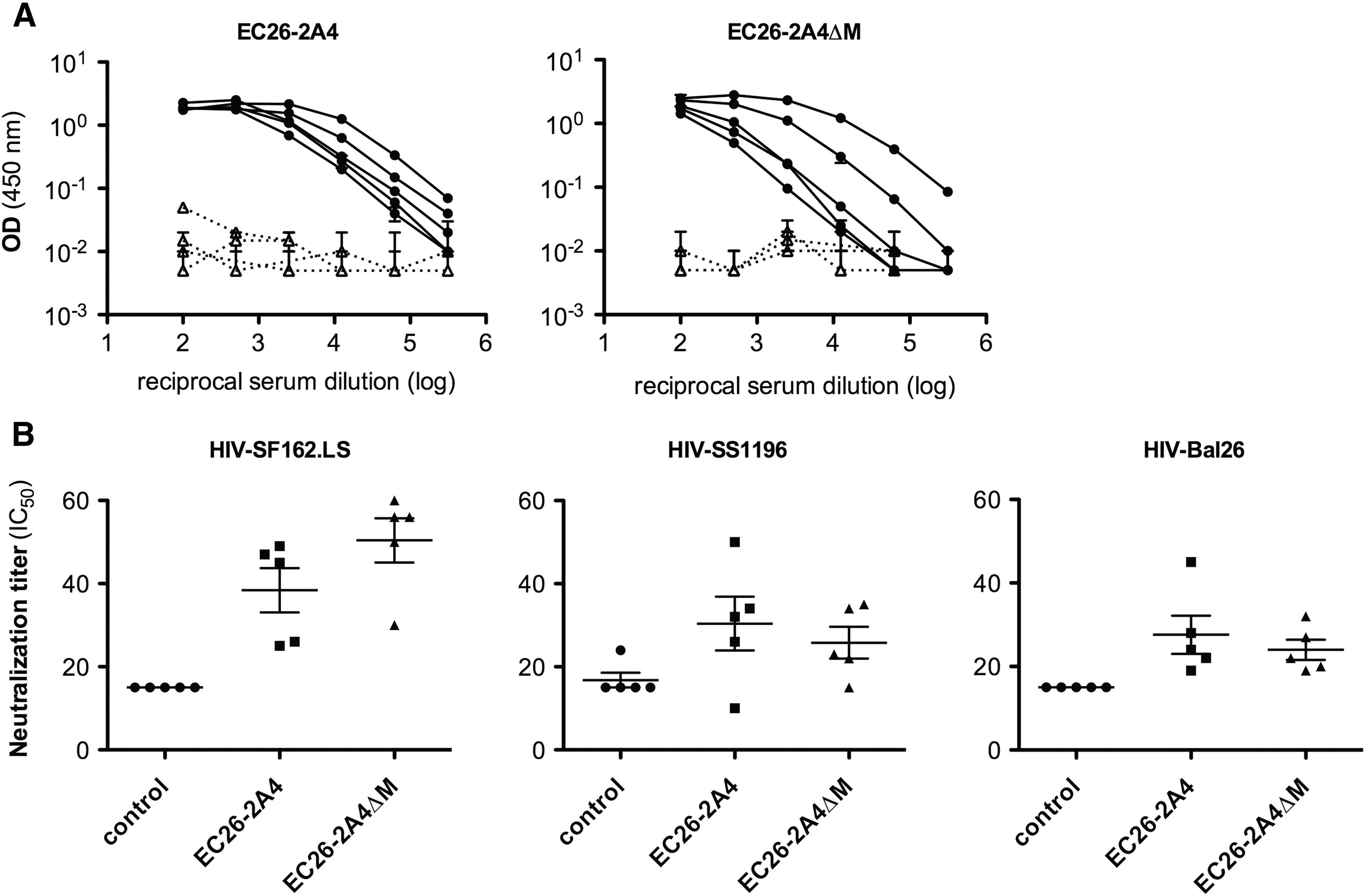
The EC26-2A4ΔM epitope induces neutralizing antibodies upon vaccination of mice.
No neutralizing antibodies were induced against the tier 2 strain THRO04156.18, which was neutralized by the original EC26 patient plasma 40 and the only one tested here due to the limited amount of mouse sera. No autoreactive antibodies against cardiolipin were induced in mice vaccinated with the EC26-2A4 and the EC26-2A4ΔM peptides as measured by commercial ELISA (data not shown).
Prevalence of EC26-2A4ΔM binding antibodies in stored plasma samples from HIV-positive cohorts
To estimate the clinical relevance of antibodies targeting the EC26-2A4ΔM epitope, we first screened 979 stored plasma samples from the HIV Center in Frankfurt for the presence of such antibodies by ELISA (Table 1). These plasma samples were analyzed from an early (<6 months after diagnosis) and a later timepoint (2–5 years after diagnosis). Overall, 83 plasma samples (8.5%) reacted with the peptide. Interestingly, the plasma samples being positive at both timepoints originated from patients with the highest CD4 nadir (Fig. 4A). Otherwise, there was no correlation between the presence of epitope-specific antibodies and clinical parameters like viral load and CD4 cell number, which might be attributed to the fact that these patients received various regimens of antiviral therapy potentially hiding such effects.

Analysis of EC26-2A4ΔM antibodies present in sera from HIV-infected patients and correlation with clinical data.
Median values are indicated.
ART, antiretrovirtal therapy.
For this reason, we also analyzed 71 plasma samples from the French ANRS CO15 ALT LTNP cohort comprising untreated patients 55 for the presence of antibodies against the EC26-2A4ΔM peptide alone and in conjunction with the 3S epitope in gp41, which has been previously associated with a protective effect in this cohort. 60,61 Interestingly, 19 of the 71 plasma samples (26.8%) showed reactivity with the EC26-2A4ΔM peptide, whereas only about 8.5% reacted in the treated cohort. Although there was a correlation between the presence of EC26-2A4ΔM antibodies in the LTNP cohort and reduced viral load in these patients (Fig. 4B), this effect was mostly due to the fact that 9 out of the 19 reacting sera also had antibodies against the 3S epitope (Fig. 4C).
EC26-2A4ΔM purified plasma antibodies show neutralizing activity and no autoreactivity with phospholipids
We purified epitope-specific plasma antibodies from eight samples of the German cohort reacting with the EC26-2A4ΔM peptide to evaluate their neutralizing activity and potential autoreactivity with cardiolipin and phosphatidylserine, two antigens known to cross-react with MPER mAbs 4E10 and 2F5. Out of the eight sera tested, seven neutralized between one and five pseudoviruses, including some tier 2 strains (Table 2). The neutralizing activity was comparable to that of the EC26 plasma antibodies previously purified with the original longer EC26-2A4 peptide. 40 There was also no cross-reactivity of the purified plasma antibodies from the eight samples with cardiolipin and phosphatidylserine (Fig. 5), the latter being even lower compared with the original plasma EC26 antibodies. 40 As expected, mAb 4E10 showed strong reactivity with both antigens, whereas mAb 2F5 reacted with cardiolipin, but not with phosphatidylserine.
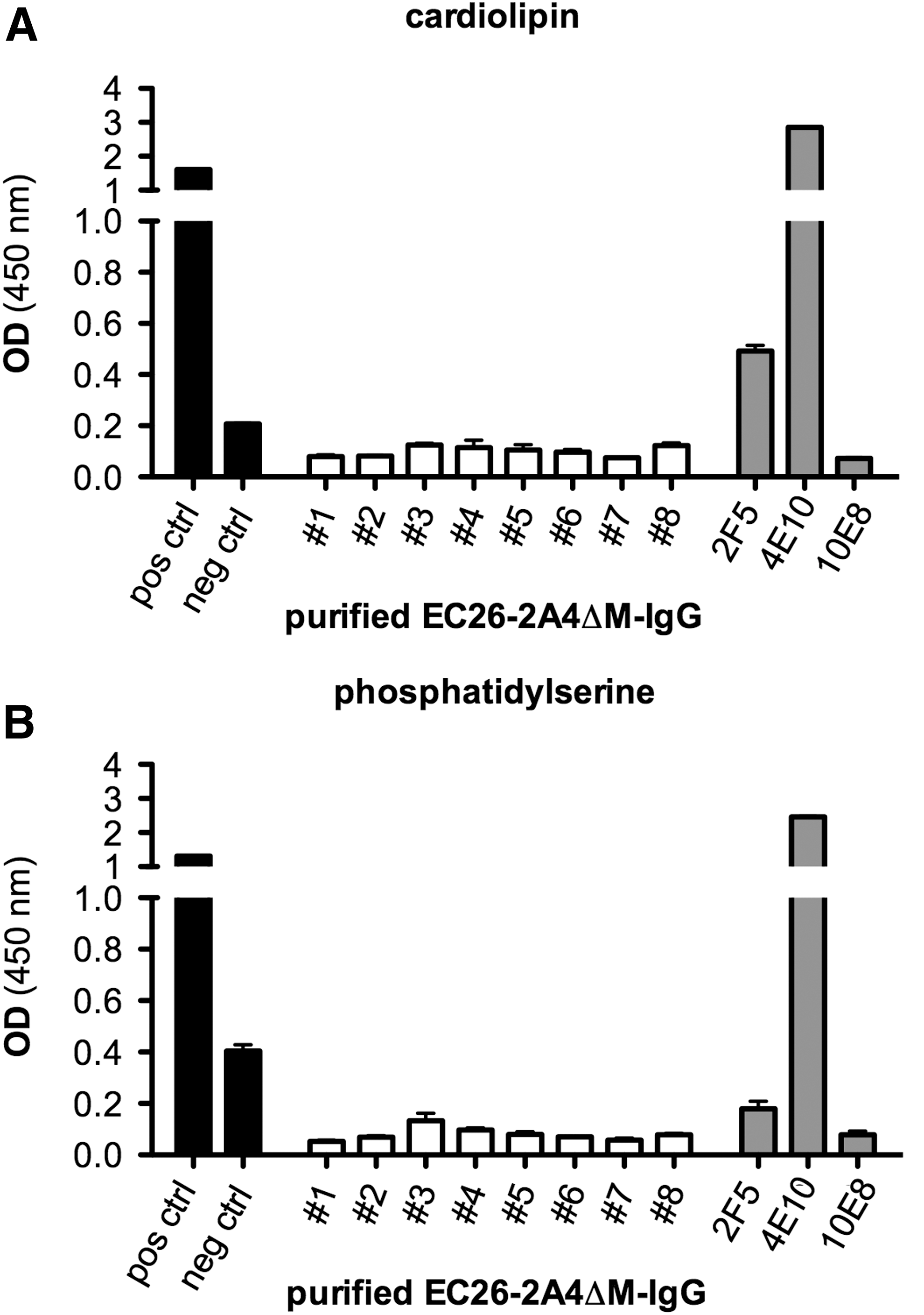
Purified EC26-2A4ΔM-specific antibodies do not show autoreactivity with phospholipids. ELISA reactivity of purified EC26-2A4ΔM antibodies (10 μg/ml) with
Mean IC50 values derived from the TZM-bl neutralization assays are indicated in μg/ml. Neutralization activity (IC50 ≤ 20 μg/ml) is marked in gray.
ART, antiretrovirtal therapy; n.d., not determined.
Discussion
BnAbs against HIV-1 need time to develop, as they require a mutual process of selection and adaptation to the coevolving HIV-1 quasispecies in the patient, finally resulting in bnAbs with peculiar features such as high affinity maturation, long HCDR3 loops, and so on. 62,63 However, bnAbs against HIV-1 are generated in an immune system that is damaged from very early on due to chronic hyperactivation, and B cell responses in particular are compromised due to loss of CD4 helper T cells, a shift from naive to memory B cells and prematured class switch. 64 For this reason, some years ago, we chose to study HIV neutralizing antibodies from HIV controllers (LTNPs and elite controllers) with a much less compromised immune system, after proving that their plasma samples showed better neutralizing activity against a panel of HIV-1 strains than control plasma samples from progressors with comparable viral load and CD4 cell count. 40,65,66 In the course of these studies we identified a new epitope in the MPER, EC26-2A4, which is targeted by neutralizing antibodies present in plasma from the elite controller EC26. 40,52
The gp41 MPER is a major target for bnAbs and as such is in focus of vaccine development since many years. 30,37 –39,67,68 MPER epitopes for bnAbs are generally exposed on fusion intermediates within the timeframe between receptor-mediated activation of the native trimeric Env spike and membrane fusion. 34,69 Furthermore, the MPER epitopes are partially covered by membranes due to their juxtaposition to the gp41 transmembrane domain. Therefore, bnAbs against MPER epitopes often contact nearby membrane envelope components with their long HCDR3 loops resulting in better extraction of the actual target epitopes. 31,47,70 Due to this fact, MPER bnAbs are often associated with autoimmunity.
In this study, we further analyzed and optimized the EC26-2A4 epitope in view of vaccination purposes. By defining the essential amino acids in the EC26-2A4MZ core epitope, which are responsible for reactivity with EC26 plasma antibodies, while not reacting with the autoreactive mAb 2F5 with an overlapping epitope, we could define a new optimized epitope, EC26-2A4ΔM. We could show that this epitope is able to induce neutralizing antibodies against tier 1 strains upon vaccination in mice. Although the original plasma from the EC26 patient neutralized 10/11 tier 2 viruses tested, 40 mice sera did not neutralize the only tier 2 strain tested, THRO04156.18. This was not unexpected, as mice usually do not show tier 2 neutralizing activity upon vaccination and even monkeys and rabbits so far show neutralizing antibodies, at best, against autologous tier 2 strains in recent studies, where native-like Env spikes have been used for immunization. 71
We further analyzed the presence of antibodies against the optimized EC26-2A4ΔM epitope in HIV-positive patient sera to study their clinical relevance. There was no evident correlation between EC26-2A4ΔM antibodies and viral load or CD4 count in the German cohort. This was not surprising, as potential effects may have been skewed by the antiviral treatment. However, despite the fact that patients from this cohort were infected for many years and on several antiretroviral treatments, an interesting observation was that the 33 patients having anti-EC26-2A4ΔM antibodies at the early and late timepoint of sampling had the highest CD4 nadirs. In clinical practice, CD4 cell nadir of untreated individuals may not only predict the magnitude of viral reservoirs and T cell recovery during treatment, 72,73 it furthermore determines the clinical stage of HIV infection according to CDC. 74
A recent randomized clinical trial revealed a clinical benefit for HIV-infected individuals starting antiretroviral therapy immediately at high (>500/μl) CD4 cell count nadir, compared to deferred therapy initiation at lower nadir cell count (<350/μl). 75 This could indicate some clinical relevance of CD4 cell count nadir for the development of antibodies targeting the EC26-2A4ΔM epitope. To elucidate this further, we also analyzed patient sera from a nontreated LTNP cohort (ANRS CO15 ALT cohort) for the presence of EC26-2A4ΔM specific antibodies. Although a much higher proportion of sera (about 27%) reacted with EC26-2A4ΔM in this cohort and an association with lower viral load in the corresponding patients was observed, a more detailed analysis showed that this association was due to the fact that about 50% of the EC26-2A4ΔM positive sera also contained antibodies against the gp41 3S peptide, which was previously associated with lower viral load in this cohort. 60 After analyzing sera single positive for EC26-2A4ΔM antibodies, there was no association with viral load or CD4 counts. Unfortunately, there are no data on CD4 nadir available for the French cohort.
Due to the neutralizing potential of EC26-2A4ΔM antibodies, in the absence of autoreactivity, and the capacity of antibodies against the 3S peptide to reduce CD4 depletion by downregulating the activating NKp44L expression on uninfected CD4+ T cells, 61 both gp41 epitopes in combination may represent an ideal dual vaccine to achieve both, protection/reduction of infection and preservation of CD4 T cells.
Footnotes
Acknowledgments
The authors thank Patricia Schult-Dietrich for expert technical assistance and Ralf Dürr for his help to initiate this project. This project was funded by the European HIVERA project HIVDUAVAX through the German Federal Ministry of Education and Research (01KI1306) and the French Agence Nationale de Recherche sur le SIDA et les Hepatitis Virales (ANRS; no. 009/JTC 2013). The Georg-Speyer-Haus is supported by the Federal Ministry of Health and the Ministry for Higher Education, Science and the Arts from the state of Hessen.
Author Disclosure Statement
No competing financial interests exist.