
Abstract
Bone marrow transplantation is a treatment for various cancers and genetic diseases, and the only case of a cured HIV infection involved the use of this clinical procedure, highlighting the potential use of this therapy for curing many chronic diseases. However, little is known about how chronic viral infection influences lymphocyte reconstitution after bone marrow transplantation. To address this, we infected mice with chronic lymphocytic choriomeningitis virus, and performed bone marrow transplantation to assess lymphocyte reconstitution. Interestingly, we observed that adoptively transferred marrow cells exhibited preferential B cell differentiation in chronically infected mice. Moreover, donor marrow cells that were adoptively transferred into chronically infected mice differentiated into virus-specific CD8 T cells that were able to expand after PD-L1 blockade. Taken together, our data show that chronic viral infection induces a biased differentiation of bone marrow stem cells into B cells, and that exhausted virus-specific CD8 T cells generated de novo in this setting are rescuable by PD-1 blockade. These data contribute to the understanding of how chronic viral infection impacts lymphocyte reconstitution, and may provide valuable information to improve current hematopoietic transplantation regimens in chronically infected hosts.
Introduction
B
Chronic infections, such as HIV, HBV, HCV, and tuberculosis, affect many people worldwide, 12 and it is important to understand how persistent infections influence immune responses, since this may help to develop safer and more effective therapies. Prior reports have demonstrated that immune exhaustion during chronic infections and cancers can be partially reverted with checkpoint blockade, 13 –15 costimulation triggering, 16 T regulatory cell depletion, 17,18 cytokine modulation, 19 –22 or interferon blockade. 23,24 However, it is unknown whether hematopoietic transplantation can be combined with an immune rescue regimen, such as PD-1/PD-L1 blockade, which is also a treatment of choice for various cancers in humans. PD-1/PD-L1 blockade has been shown to induce the functional rescue of exhausted CD8 T cells that express CXCR5 and intermediate levels of PD-1, and it has been proposed that the efficacy of immune reinvigoration after blockade of this inhibitory pathway is dependent on costimulation. 25 –27 However, it is unknown whether de novo generated CD8 T cells generated during the chronic phase of a persistent infection can expand after PD-1/PD-L1 blockade.
In this study, we utilized the model of lymphocytic choriomeningitis virus (LCMV) infection in mice to investigate the effect of chronic viral infection on hematopoietic differentiation and de novo lymphocyte production. We show that acutely infected mice exhibit only transient changes in lymphoid tissues, especially at the peak of the effector T cell response, whereas chronically infected mice exhibit protracted involution of lymphoid tissues. Our data also demonstrate that chronic viral infection induces accelerated uptake and differentiation of donor bone marrow cells, and importantly, we show that de novo generated CD8 T cell responses generated in the chronic phase of a persistent viral infection can be rescued by checkpoint blockade.
Materials and Methods
Mice, immunizations, and treatments
Six- to eight-week old female C57BL/6 mice were purchased from Jackson Laboratories (Bar Harbor, ME). For acutely controlled viral challenges, mice were infected with LCMV Armstrong, whereas for chronic viral challenges, mice were infected with LCMV Cl-13. Viral infections were performed intravenously (i.v.) through the lateral tail vein at 2 × 106 PFU/mouse. To induce a more protracted lifelong LCMV Cl-13 infection, we utilized the stringent LCMV Cl-13 infection model developed by Drs. Matloubian and Ahmed. 28 In this model, mice were briefly CD4 T cell depleted (0.5 mg GK1.5 mAb) through the intraperitoneal route for 2 consecutive days before chronic viral challenge. For hematopoietic reconstitutions, we conditioned the recipient mice with busulfan. One day before bone marrow transfer, 0.6 mg of busulfan diluted in PBS was administered intraperitoneally. A total of 20 × 106 of congenically distinct bone marrow cells were infused 1 day after. All experiments were performed with approval of the Institutional Animal Care and Use Committee.
Virus titration
Titration of LCMV was performed on VERO E6 cell (ATCC) monolayers by plaque assay as previously shown. 29 Three 10-fold serial dilutions of mouse sera in Dulbecco's modified Eagle's medium without fetal bone serum (FBS) were gently distributed on top of the VERO E6 cell monolayers in six-well plates at around 90% confluency. Plates were incubated for 1 h rocking every 10 min, followed by overlay with a 1:1 solution of 1% agarose in 2 × 199 media (Gibco, Life Technologies) at 40°C, and after the media solidified, plates were transferred to a CO2 37°C incubator. Ninety-six hours later, a 1:1 solution of 1% agarose in 2 × 199 media with 1:50 neutral red at 40°C was pipetted on the wells. Plaques were enumerated the next day.
Antibodies and flow cytometry
Lymphocytes were stained with anti-CD8α (53–6.7 on PerCP-Cy5.5), -CD44 (IM7 on Pacific Blue), -PD-1 (RMP1-30 on PE), and -CD4 (RM4-5 on PerCP-Cy5.5). Antimouse flow cytometric antibodies were purchased from BD Pharmingen, except for CD44 (which was from Biolegend). Major histocompatibility complex (MHC) I monomers were obtained from the NIH Tetramer facility, and were then conjugated to fluores. Samples were acquired using Canto II flow cytometer (BD Biosciences) and analyzed using FlowJo (Treestar).
Statistical analysis
Analyses were done with two-tailed nonparametric Mann–Whitney U tests using GraphPad Prism software. Statistical analyses in survival plots were performed using the Mantel–Cox test.
Results
Lymphoid tissue homeostasis after acute and chronic viral infection
We first evaluated lymphoid tissue architecture after viral infection to assess how viral persistence influenced the sites where lymphocytes develop and undergo priming. We utilized the classical model of LCMV infection in mice; LCMV Armstrong results in an acute viral infection that is cleared within a week, and induces functional immune responses. Whereas LCMV Cl-13 results in a chronic viral infection and induces immune exhaustion. 11 We analyzed how these two distinct immune conditions influenced lymphoid tissue integrity by enumerating lymphocytes at lymphoid sites and performing H&E stains. Interestingly, viral infection resulted in a significant decrease in thymic cellularity independent of whether the viral infection persisted. By day 7 postinfection, both the acutely controlled LCMV Armstrong infection and the chronic LCMV Cl-13 infection induced a significant (p < .01) decrease in thymic cellularity relative to naive mice (Fig. 1A). However, LCMV Armstrong-infected mice showed a gradual rebound in thymocyte numbers after day 15 postinfection, whereas chronic LCMV Cl-13-infected mice showed a continuous decline in thymocyte counts (Fig. 1A). By day 15 postinfection, chronic LCMV Cl-13-infected mice exhibited pronounced thymic involution, evidenced by histological analyses (Fig. 1B). Since de novo T cell generation is shown to occur in the thymus, these data suggested that chronic viral infection may impair de novo T cell generation from hematopoietic precursors.
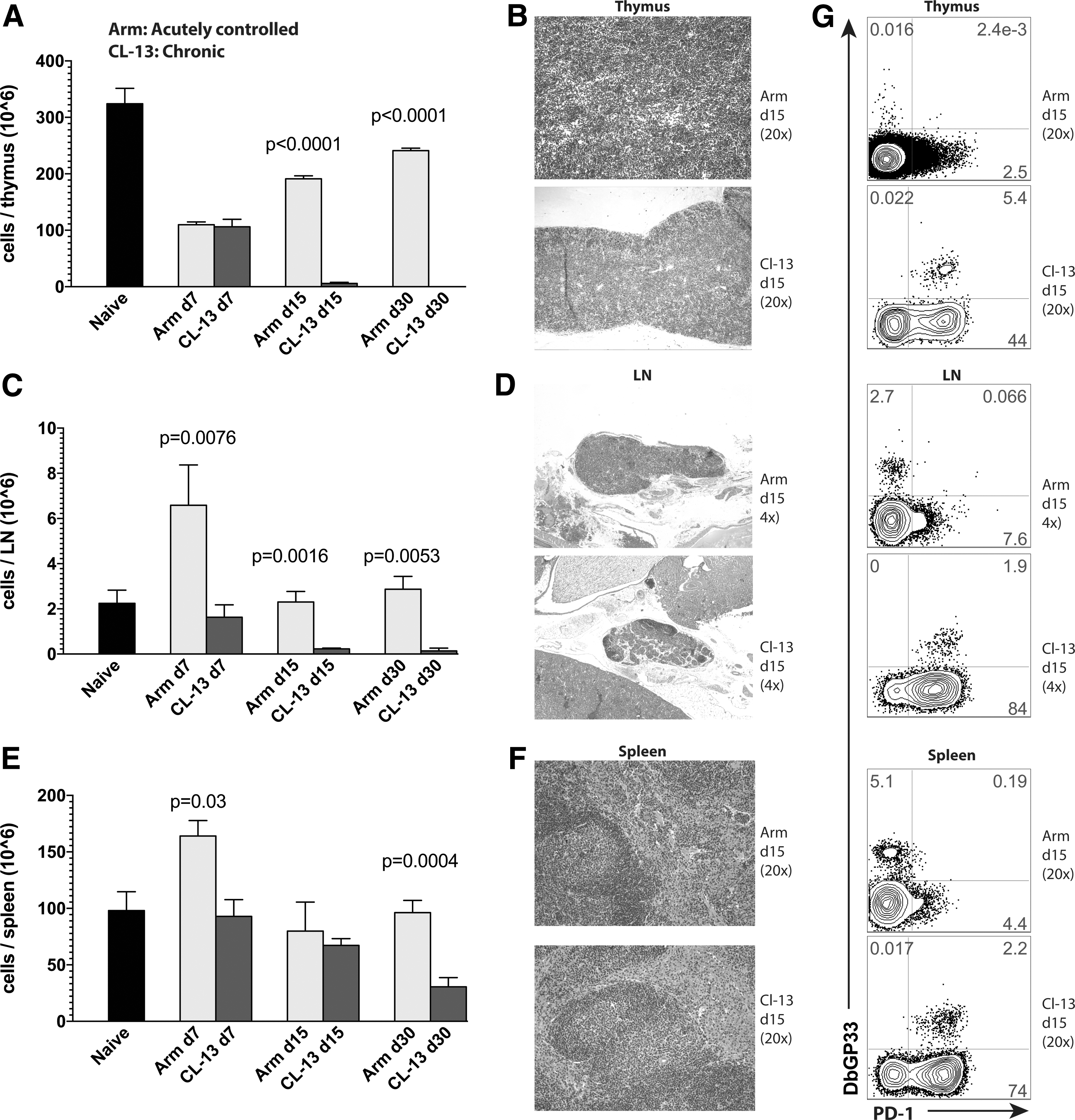
Acutely controlled viral infection induces transient changes in lymphoid tissues, whereas chronic viral infection induces protracted changes in lymphoid tissues.
In addition, we compared lymph node dynamics after viral infection. Acutely controlled LCMV Armstrong infection resulted in a threefold increase in inguinal lymph node cellularity by day 7 postinfection (p = .021, relative to naive mice), whereas chronic LCMV Cl-13 infection resulted in progressive lymph node involution (Fig. 1C). Lymph node involution was also evidenced histologically; chronic LCMV Cl-13-infected mice exhibited extensive areas of lymph node fibrosis at day 15 postinfection (Fig. 1D). Although spleen cell counts were not statistically different at day 15, by day 30 postinfection, chronically infected mice exhibited a 3.2-fold reduction (p = .0004) in splenocyte counts relative to acutely infected mice (Fig. 1E, F). Another interesting observation was that thymic involution in chronically infected mice was associated with high levels of exhausted (PD-1+) virus-specific CD8 T cells into the thymus, whereas mice that cleared viral infection exhibited negligible frequencies of virus-specific CD8 T cells in the thymus (Fig. 1G). However, all other sites showed increased frequencies of virus-specific CD8 T cells in mice that cleared viral infection relative to chronically infected mice (Fig. 1G), consistent with prior reports showing the effect of high antigen loads in inducing T cell exhaustion. 11,14,30,31 Another interesting observation is that the peak of the effector T cell response (day 7) after acute or chronic viral infection was associated with decreased frequencies of double positive (DP) thymocytes (Fig. 2). Importantly, mice that cleared the LCMV Armstrong infection recuperated their normal DP thymic precursor frequencies after day 15, whereas chronically infected mice continued to lose this population (Fig. 2).
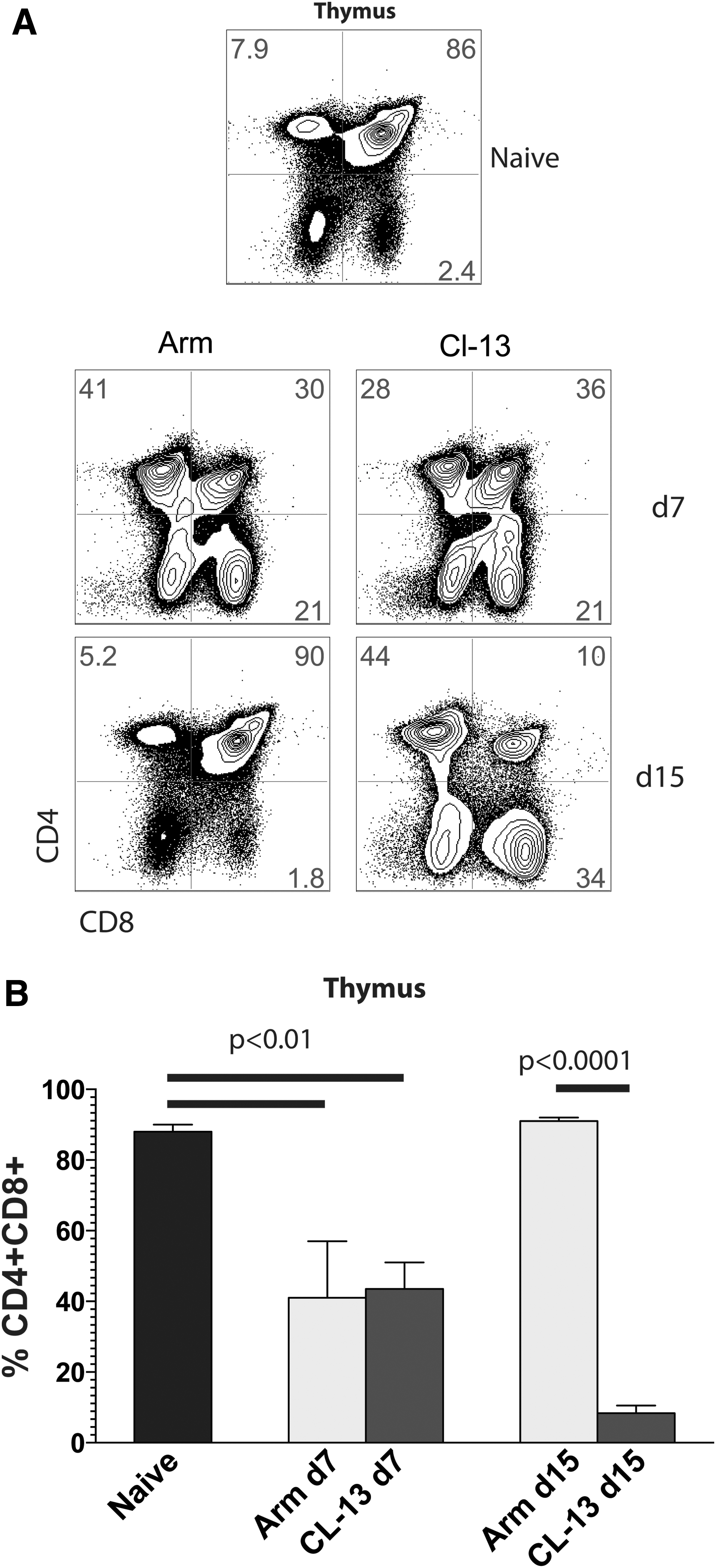
Dynamics of thymic differentiation after acute or chronic viral infection.
Altogether, these data demonstrated that an acutely controlled viral infection induced very transient changes in lymphoid tissues, with lymphoid sites rapidly becoming similar to those of uninfected mice. In contrast, persistent viral infection induced protracted changes in lymphoid tissues. The persistence of viral antigen induced severe involution of lymphoid tissues (especially the thymus, which appeared completely absent at day 30 postinfection). Altogether, these changes in lymphoid tissues suggested aberrant lymphocyte dynamics during chronic viral infection.
Chronically infected mice exhibit accelerated lymphocyte reconstitution
The data from above suggested that bone marrow transplantation in chronically infected mice may result in abnormal lymphocyte reconstitution. To rigorously address this, we used LCMV Cl-13 chronically infected CD45.2+ mice. After 60 days postinfection, we treated these mice intraperitoneally (i.p.) with 0.6 mg of busulfan, which partially depletes the mouse bone marrow, allowing space for donor marrow engrafting. 32 One day after busulfan conditioning, we injected i.v. 20 × 106 donor CD45.1+ bone marrow cells isolated from the femurs of naive mice to generate bone marrow chimeras. Since recipient and donor mice express different congenic markers (CD45.2 vs. CD45.1), we were able to distinguish between recipient (endogenous) and donor-derived cells by flow cytometry. Uninfected treatment-matched CD45.2+ mice served as experimental controls to compare hematopoietic transplantation relative to chronically infected mice (Fig. 3A).
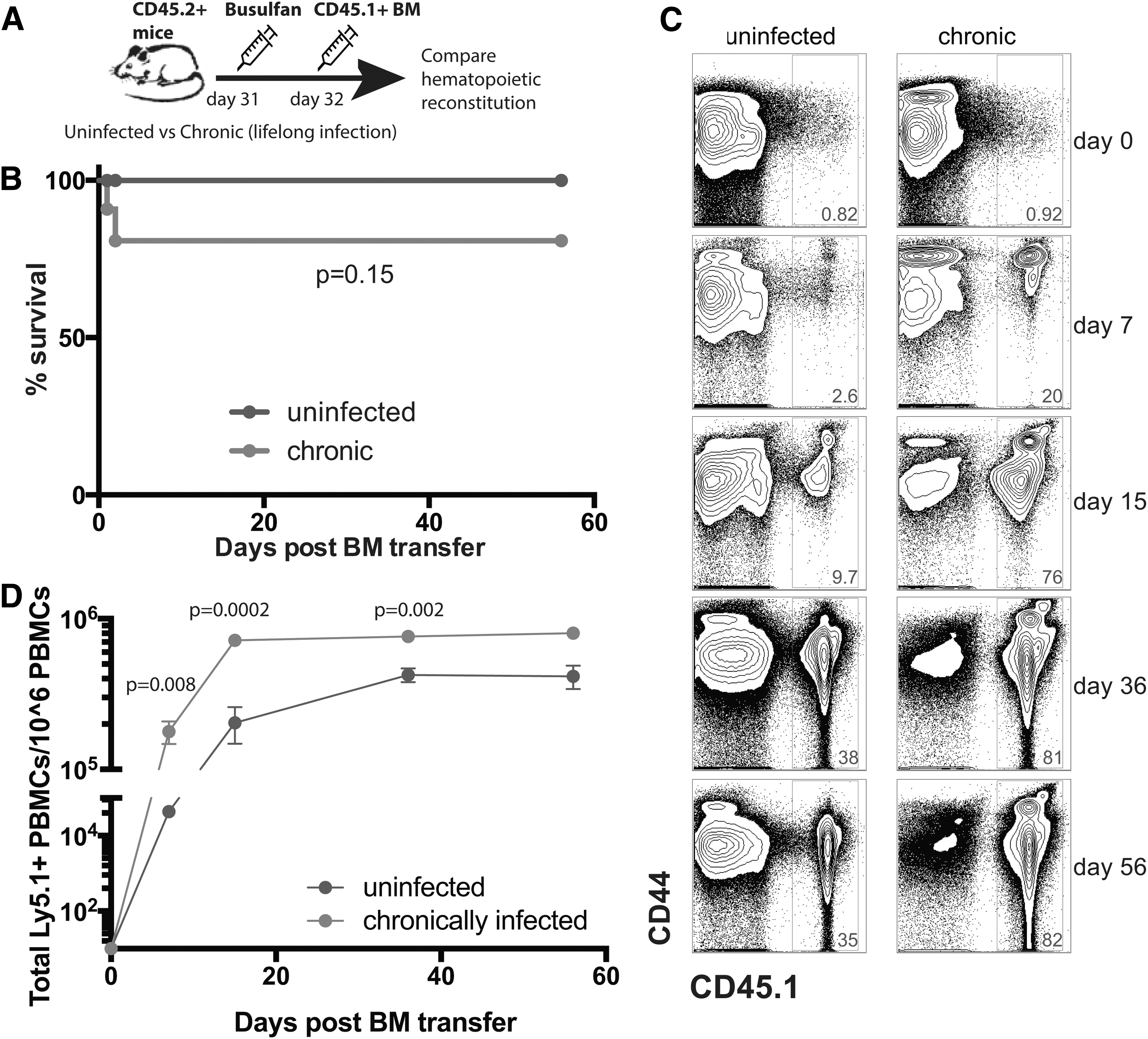
Accelerated differentiation of hematopoietic cells in chronically infected mice.
Bone marrow transplantation was well tolerated in uninfected mice with 100% survival, but it resulted in low-level mortality (∼20%) in chronically infected mice, but this was not statistically significant (p = .15, Fig. 3B). Interestingly, and contrary to our initial prediction, chronically infected mice showed accelerated lymphocyte reconstitution from hematopoietic precursors relative to uninfected mice (Fig. 3C, D). Altogether, these results showed that although chronically infected mice had severe lymphoid tissue atrophy, they were able to rapidly generate differentiated lymphocytes from hematopoietic precursors, suggesting a role for immune activation in influencing donor marrow uptake and lymphocyte turnover.
Biased B cell differentiation of donor-derived bone marrow cells in chronically infected mice
The data from above suggested rapid lymphocyte differentiation from hematopoietic stem cell precursors in chronically infected mice. To ascertain whether a specific lymphocyte subset accounted for the expanded hematopoietic reconstitution in chronically infected mice, we assessed the levels of donor-derived (CD45.1+) CD4 T cells (CD3+ CD4+), CD8 T cells (CD3+ CD8+), and B cells (CD3- B220+ CD19+) in peripheral blood mononuclear cells (PBMCs) at various time points after bone marrow transplantation. Chronically infected mice and uninfected mice exhibited similar levels of donor-derived CD4 T cells (Fig. 4A) and CD8 T cells (Fig. 4B). Interestingly, in both groups of mice (uninfected or infected), we noticed a pattern of faster CD8 T cell reconstitution relative to CD4 T cell reconstitution during the first week of transfer (p < .001), but at later time points donor-derived CD4 T cells were more predominant relative to donor-derived CD8 T cells (Fig. 4A, B). Although T cell reconstitution kinetics were similar in both groups of mice, chronically infected mice showed significantly greater levels of donor-derived B cells (Fig. 4C). This suggested that bone marrow transplantation during chronic viral infection results in a more pronounced de novo B cell production relative to normal mice. This may be explained by the greater frequencies of T follicular helper (Tfh) cells present during chronic viral infection (Fig. 4D).
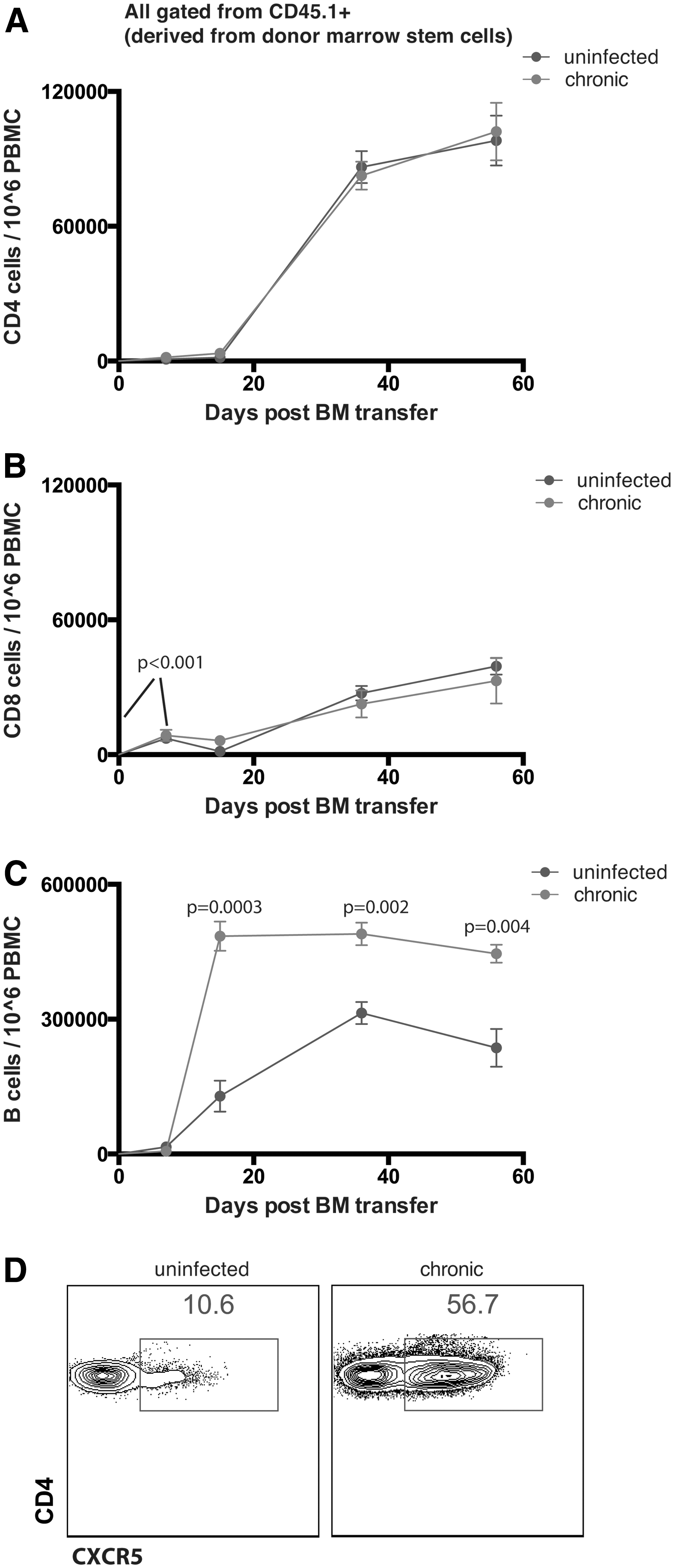
Biased differentiation of donor-derived marrow cells into B cells in chronically infected mice.
Bone marrow transplantation in chronically infected mice induces de novo virus-specific CD8 T cells that can be rescued by PD-L1 blockade
Bone marrow transplantation is a promising therapy for various cancers and chronic infections. Although continuous recruitment of de novo-generated T cells has been shown to occur during chronic viral infections, 32 –34 it is not clear whether CD8 T cell subsets derived from donor stem cells late after infection can be reinvigorated by PD-L1 blockade. To address this simple question, we utilized CD45.2+ chronically infected mice (LCMV Cl-13), and after 30 days we treated them with busulfan followed by adoptive transfer of 20 × 106 CD45.1+ bone marrow cells i.v. 1 day after. We then administered i.p. 200 μg of PD-L1 blocking antibodies or isotype control (every 3 days, five times), 50 days after bone marrow transfer, and we evaluated the de novo-generated donor-derived CD8 T cell response to LCMV after 2 weeks of anti-PD-L1 treatment (Fig. 5A).
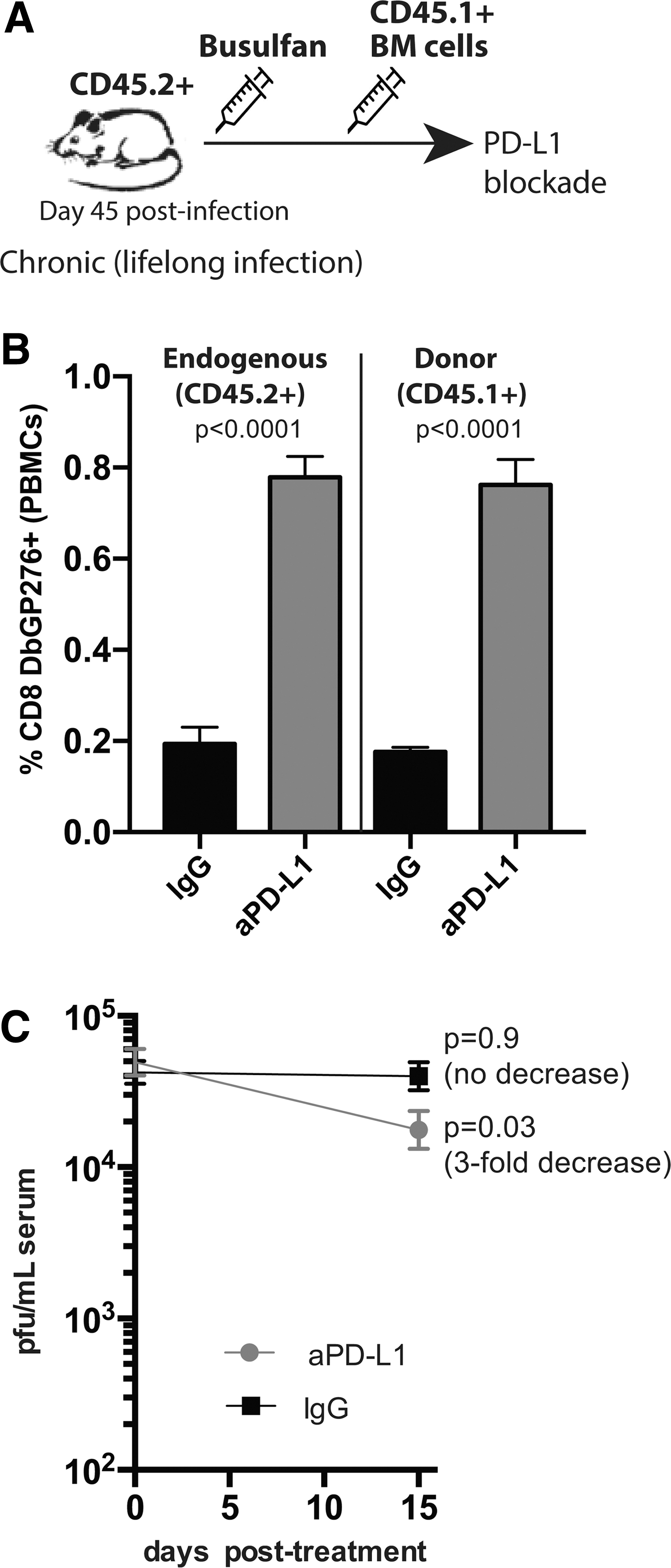
De novo virus-specific CD8 T cells derived from donor bone marrow expand after PD-L1 blockade during chronic viral infection.
Interestingly, PD-L1 blockade resulted in rescue of both the endogenous (CD45.2+) and the donor-derived (CD45.1+) virus-specific CD8 T cell response (Fig. 5B), and was able to reduce viral loads (Fig. 5C). These results demonstrate that hematopoietic transplantation can be combined with checkpoint blockade, and that de novo-generated CD8 T cells derived from donor stem cells can be rescued by PD-L1 blockade.
Discussion
Bone marrow transplantation is a widely utilized therapy consisting of the intravenous infusion of donor hematopoietic cells, which can be used to reconstitute the hematopoietic compartment of the recipient. This medical procedure represents a major advance in the treatment of various hematopoietic malignancies and genetic disorders. In addition, bone marrow transplantation is given to patients undergoing high-dose chemotherapy for the treatment of solid tumors or after whole body irradiation, since such palliative regimens can result in high hematogenous toxicity and ablation of endogenous hematopoietic cells. Moreover, the only reported cure of an HIV infection, also referred to as the “Berlin patient,” involved a bone marrow transplantation protocol, suggesting the potential wide use of this procedure for the treatment of chronic viral infection.
Although bone marrow transplantation is used in the clinic for the treatment of various diseases, very little is known about how chronic viral infection influences the dynamics of lymphocyte reconstitution after marrow transplantation. In this study, we assessed T cell and B lymphocyte reconstitution after bone marrow transplantation in chronically infected mice, and we made various key observations. We noticed that chronic viral infection induces severe involution of lymphoid tissues with complete involution of the thymus after day 15, which corresponded to a time when high levels of exhausted (PD-1+) virus-specific CD8 T cells infiltrated the thymus. Prior studies have suggested a role for lymphotoxins, interferons, and NK1.1+ cells in coordinating lymphoid involution and immunosuppression during chronic viral infection. 35 –37 Note that thymocyte atrophy has been shown to result in rapid apoptotic cell death of DP thymocyte precursors, 6 which is consistent with our results showing that when thymocyte counts decrease, DP cells are greatly reduced (Figs. 1A and 2).
We also report that busulfan, which is an alkylating agent that creates physical space in the marrow to allow for the engraftment of donor stem cells, followed by bone marrow transplantation, induces low-level mortality rates in 20% of chronically infected mice (p = .15). Although this difference in survival was not statistically significant, it was reproducible in two separate experiments. It is not clear what are the reasons for this low level mortality rates in chronically infected mice, but it may be due to the fact that chronically infected mice exhibit partial emaciation due to the constant cytotoxic response, which could increase their susceptibility to succumb after experimental manipulations. Interestingly, our study demonstrates that chronic viral infection accelerates the rate of lymphocyte reconstitution after bone marrow transplantation. Prior studies have demonstrated that stem cell mobilization and differentiation can be influenced by inflammatory cytokines, 38,39 and therefore, a possible reason for the accelerated hematopoietic reconstitution in chronically infected mice could be inflammation caused by the persistent viral infection. However, one cannot rule out that selective depletion of the recipient bone marrow is the reason for the increased levels of donor bone marrow in chronically infected mice. In fact, chronic infection is known to induce hematopoietic turnover and depletion of hematopoietic cells, 32,40,41 and thus, it is possible that the recipient bone marrow contains cell precursors that have already reached their Hayflick limit.
Of note, very high numbers of donor-derived (CD45.1+) stem cells differentiated into B cells in chronically infected mice. Prior studies have demonstrated that chronic viral infections, such as HIV, SIV, and chronic LCMV, induce biased Tfh cell differentiation. 42 –44 Since Tfh cells interact with B cells to induce their expansion and induce the antibody response, it is possible that the biased B cell differentiation after bone marrow transplantation during chronic viral infection is due to increased levels of Tfh cells.
In addition to bone marrow transplantation, another potential treatment for chronic infections and cancers is immune checkpoint blockade therapies, such as PD-1/D-L1 blockade, which have been shown to reinvigorate exhausted virus-specific CD8 T cells. 14,45 –47 However, it is not clear whether this checkpoint therapy can have an equal effect on endogenous or donor-derived CD8 T cells. It is also not clear whether PD-1 blockade rescues only the exhausted CD8 T cells that were primed during the early stages of chronic infection (before bone marrow transplantation). To address this question, we treated chronically infected mice (day 30 postinfection) with busulfan and injected bone marrow cells 1 day after treatment. Several weeks after marrow grafting, chronically infected mice were treated with PD-L1 blocking antibodies, and endogenous (CD45.2+) versus donor-derived (CD45.1+) CD8 T cell responses were compared.
Interestingly, our data show that PD-L1 blockade resulted in similar expansion of both endogenous (CD45.1-) and donor-derived (CD45.1+) LCMV-specific CD8 T cells. Since the donor bone marrow was infused during the onset of chronic viral infection, we can reasonably conclude that PD-L1 blockade can also rescue de novo-generated CD8 T cells. This demonstrated that hematopoietic transplantation can provide a new pool of virus-specific CD8 T cells that can be rescued by PD-L1 blockade. Although bone marrow transplantation followed by PD-L1 blockade appeared to be a safe dual treatment in chronically infected mice, the effect of this dual regimen in humans remains to be rigorously addressed. Our experiments involved the transfer of MHC-matched cells, but prior clinical studies have reported that blockade of the PD-1 pathway may exacerbate graft versus host disease (GVHD) after allogenic stem cell transfer. 48 –52 Therefore, future studies are needed to rigorously assess the safety of combined hematopoietic transfer and PD-1 blockade and the level of MHC match needed to improve the efficacy of these combined regimens. Taken together, our data highlight key features of how a chronic viral infection affects lymphocyte homeostasis. Future experiments will determine whether these observations can be generalized to HIV-infected patients who receive hematopoietic transplantation as part of a cure strategy or a treatment for HIV-associated malignancies. Overall, these data may shed light into the development of novel therapies aimed to regenerate lymphocyte subsets in settings of antigen persistence.
Footnotes
Acknowledgments
This work was supported by an NIH grant to P.P.M. (1K22AI118421) and the Chicago Third Coast CFAR grant to P.P.M. (P30 AI117943). The authors declare that no financial conflicts of interests exist.
Author Disclosure Statement
No competing financial interests exist.