
Abstract
The main clinical feature of human T cell leukemia virus-1 (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) is slowly progressive spastic paraparesis with bladder dysfunction. HAM/TSP is induced by chronic inflammation in the spinal cord, mainly the lower thoracic cord. A long-standing bystander mechanism, such as the destruction of surrounding tissues by the interaction between infiltrated Th1-like, HTLV-1-infected CD4+ T cells and HTLV-1-specific CD8+ cytotoxic T cells (CTL), is probably critical for the induction of chronic inflammation. Although the HTLV-1-infected CD4+ T cells in HAM/TSP appear to play a crucial role in the initial pathogenesis of HAM/TSP, the exact mechanisms of how these cells acquire their function as the first responders in the pathogenesis of HAM/TSP still remain unresolved. Herein, we propose the importance of the activation of both outside-in signals from integrin signaling and inside-out signals for integrin signaling in the HTLV-1-infected CD4+ T cells of HAM/TSP patients.
Introduction
H
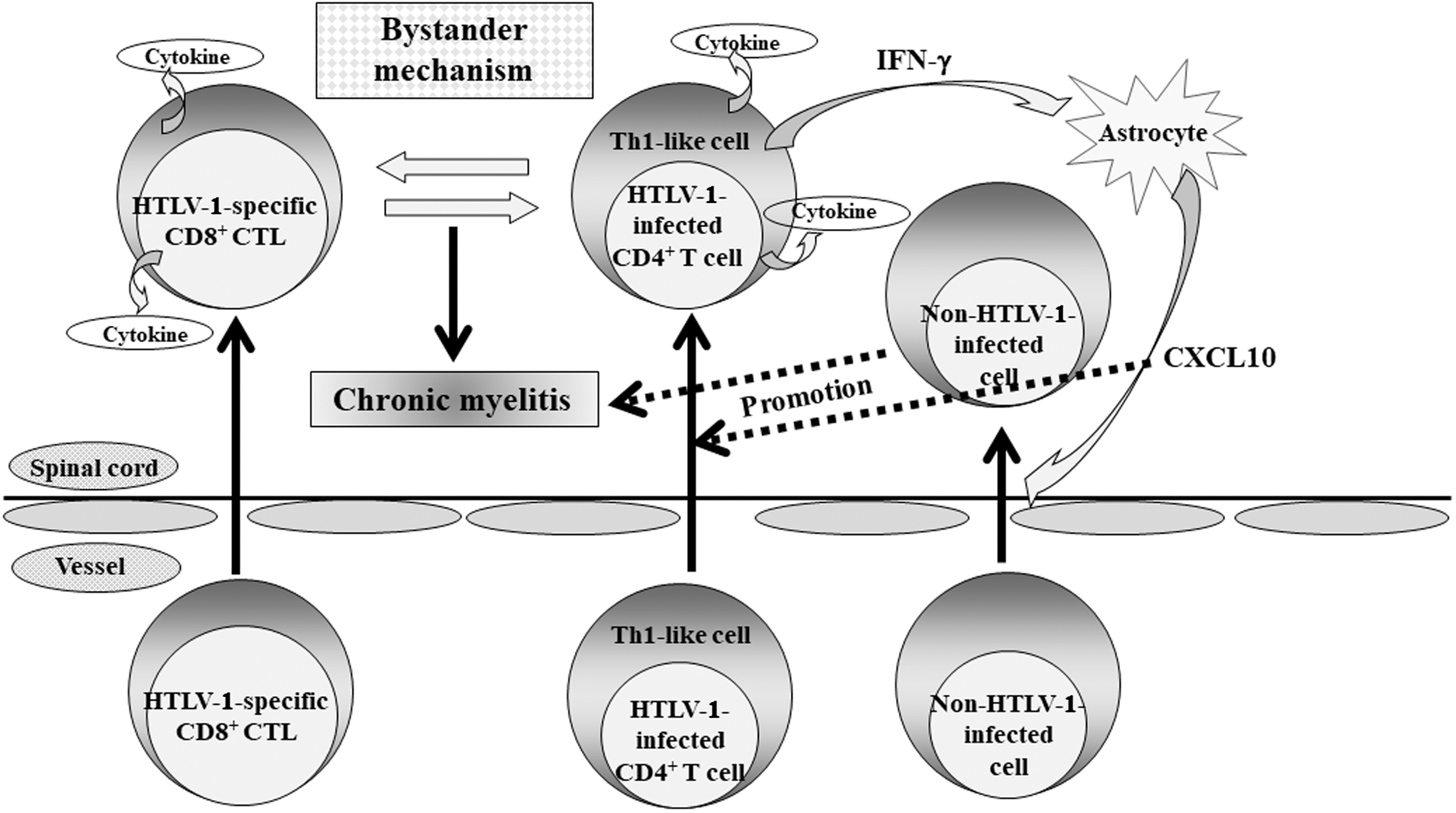
Proposed mechanism for the development of HAM/TSP. In the induction of chronic myelitis, a long-standing bystander mechanism, such as the destruction of surrounding tissues by the interaction between infiltrated Th1-like, HTLV-1-infected CD4+ T cells and HTLV-1-specific CD8+ CTL, which enter the spinal cord to make an attack on HTLV-1-infected CD4+ T cells, is probably critical. In addition, a positive feedback loop formed through the expression of chemokine CXCL10, which is a ligand of CXCR3, by astrocytes via interferon-γ (IFN-γ) from infiltrated HTLV-1-infected cells might be involved in the maintenance and promotion of chronic myelitis. CTL, cytotoxic T cells; HAM/TSP, HTLV-1-associated myelopathy/tropical spastic paraparesis; HTLV, human T cell leukemia virus-1.
HTLV-1-infected CD4+ T cell adherence to and transmigrating activity through vascular endothelial cells in HAM/TSP patients
Given the previously described functions of HTLV-1-infected CD4+ T cells as the first responders in HAM/TSP, it seems likely that their adherence to and transmigrating activity through vascular endothelial cells (ECs) may have a key role as the first trigger of HAM/TSP development. Studies demonstrated that expression of several kinds of adhesion molecules, such as intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecue-1, and cell adhesion molecule 1 etc., is upregulated in HTLV-1-infected cells. 9 –12 Very recently, it was shown that increased expression of activated leukocyte cell adhesion molecule, which might be related to the transmigration of T cells across the blood-brain barrier, is observed on the surface of HTLV-1-infected cells. 13 Selectin and its ligands, which are expressed on ECs and T cells, respectively, play an important role during rolling, the initial attachment of T cells to ECs that subsequently leads to T cell transmigration into the tissues. 14 Sialyl Lewisx antigen (sLex) is a ligand of selectin, 15 and T cells expressing sLex have the potential to transmigrate into the tissues. Although HTLV-1 tax can transactivate the expression of fucosyltransferase VII, which is a key element in regulating the synthesis of selectin ligands, 16 we previously reported a significant increase of the sLex+ cell population, in which HTLV-1-infected cells are concentrated, in the peripheral blood CD4+ T cells of HAM/TSP patients. 17 This finding indicates that the amount of HTLV-1-infected CD4+ T cells with the ability to transmigrate into the spinal cord is increased in the peripheral blood of HAM/TSP patients. The potential transmigration of HAM/TSP patient HTLV-1-infected CD4+ T cells into the tissues is supported by the exaggerated transmigrating activity through a reconstituted basement membrane of vessels that is observed for this cell population but is not observed for CD8+ T cells. 18
Activation of small GTPases in the HTLV-1-infected cells of HAM/TSP patients: upregulated status of the outside-in signal from integrin signaling
Although HTLV-1 preferentially infects CD4+ T cells, 2 we previously reported the significantly increased adherence to ECs and subsequent transmigration through ECs of activated CD4+ T cells with heightened lymphocyte function antigen-1 (LFA-1, αLβ2) expression compared with that of CD8+ T cells in the peripheral blood T cells of HAM/TSP patients. 19,20 This suggests that signaling induced by integrin/ligand interaction is upregulated in the peripheral blood CD4+ T cells of HAM/TSP patients. Integrin/ligand signaling induces the activation of small GTPases leading to the rearrangement of cytoskeletal components. 21 –23 Therefore, small GTPases may be activated in the HTLV-1-infected cells of HAM/TSP patients. It was previously demonstrated that HTLV-1 tax regulates cell adhesion and migration by the interaction with small GTPases, such as Cdc42, Rac, and Rho. 24 We analyzed the activity of small GTPases, such as Cdc42, Rac, and Rho, by using pull-down assays in three HTLV-1-infected T cell lines derived from HAM/TSP patients and three HTLV-1-infected T cell lines derived from other origins, which were used as controls. 25 These experiments revealed that all small GTPases were strongly activated in all of the cell lines derived from HAM/TSP patients.
Interestingly, of these small GTPases, the difference in the degree of activation between HTLV-1-infected T cell lines derived from HAM/TSP patients and the control cell lines was the most obvious in Cdc42. Unlike Rac or Rho, Cdc42 plays an important role in the polarization of the cytoskeleton following integrin-mediated activation, 26 and as a major regulator of cell polarity, Cdc42 is involved in cell migration. 27 Therefore, the activation of small GTPases, particularly Cdc42, in the HTLV-1-infected cells of HAM/TSP patients, which indicates a polarized status, suggests that these cells have upregulated transmigrating activity into the tissues. That is, the outside-in signal from integrin signaling appears to be strongly upregulated in the HTLV-1-infected cells of HAM/TSP patients, suggesting that this activity functions as one of the first triggers in the development of HAM/TSP (Fig. 2). In addition, our data suggest that activation of this signaling might also be involved in the efficient spread of HTLV-1 to new cells 25 because HTLV-1 spreads to cells through the virological synapse, which is composed of an integrin and its ligand and is achieved by polarization of the cytoskeleton, such as via the microtubule-organizing center. 28 Although the equilibrium condition between immune surveillance and HTLV-1 29,30 or the increased proliferation of HTLV-1-infected cells 31 are proposed as reasons for the induction of high HTLV-1 proviral load in HAM/TSP patients, the efficient cell-to-cell spread of HTLV-1 by activation of small GTPases in HTLV-1-infected cells might also partially function for the induction of this status.

Upregulated status of both the outside-in signal from integrin signaling and the inside-out signal for integrin signaling in the HTLV-1-infected CD4+ T cells of HAM/TSP patients. Activation of both outside-in integrin signaling and inside-out integrin signaling induced by the activation of small GTPases, such as Cdc 42 and Rap1, respectively, might upregulate polarization and cytoskeletal rearrangement, leading to an upregulation of cell transmigration activity into the tissues. In addition, an activating factor for CXCR4 signaling other than the SDF-1α produced from HTLV-1-infected cells might be involved in the activation of Rap1 in an autocrine manner.
Activation of CXCR4 signaling in the HTLV-1-infected cells of HAM/TSP patients: upregulated status of the inside-out signal for integrin signaling
In addition to outside-in signaling, integrins are also capable of conducting inside-out signaling, and this mechanism also plays a role in the HTLV-1-infected cells of HAM/TSP patients. Rap1, a small GTPase that functions in a complex with the regulator of adhesion and cell polarization enriched in lymphoid tissues (RAPL), is a potent stimulator of integrins and facilitates the inside-out signal for integrin signaling, leading to increased adhesion to integrin ligands, polarization of cell shape, and facilitation of cell migration. 32 When we analyzed the activity of Rap1 by using pull-down assays in an HTLV-1-infected T cell line derived from an HAM/TSP patient (HL) and an HTLV-1-infected T cell line derived from an HTLV-1 carrier (CL) as a control, we detected Rap1 activation, such as bound Rap1-GTP, in HL cells but not in CL cells. 33 Rap1 is activated by various external stimuli. 34 For example, activation of signaling by C-X-C chemokine receptor type 4 (CXCR4), a major coreceptor for human immunodeficiency virus-1 entry, 35 can induce Rap1 activation, thereby leading to initiation of the inside-out signal for integrin signaling. 36
HTLV-1 spreads to new cells through the virological synapse, which is composed of integrins on the target cells and their ligands on the HTLV-1-infected cells. 28 Therefore, the activity of integrin/ligand signaling can be evaluated by using a co-cultivation system of HTLV-1-infected T cell lines with reporter cells to measure the efficiency of HTLV-1 intercellular transmission. The finding that HL cells had a higher HTLV-1 intercellular transmission efficiency compared with CL cells suggests that HL cells are producing a substance that is activating integrin/ligand signaling. Upregulation of ICAM-1 expression in HTLV-1-infected cells might be related to the efficient spread of HTLV-1 to new cells. 9 However, because Rap1 activation was observed in HL cells, 33 we analyzed the involvement of CXCR4 signaling in the efficiency of HTLV-1 intercellular transmission in this culture system. Pretreatment of the reporter cells with an anti-CXCR4 blocking antibody induced a significant reduction in the HTLV-1 intercellular transmission in HL cells but not in CL cells. These data suggest that HL cells produce the factor inducing activation of the inside-out signal for integrin signaling through Rap1 activation via the activation of CXCR4 signaling in the reporter cells. Additionally, this factor appears to be involved in the efficient HTLV-1 intercellular transmission from HTLV-1-infected cells.
In general, the activation of CXCR4 signaling is induced by stromal cell-derived factor-1α (SDF-1α), 36 which is one of the C-X-C chemokines. However, the finding that SDF-1α was not detected in the HL cell culture supernatant suggests that HL cells are producing a different factor (something other than SDF-1α) that activates CXCR4 signaling leading to Rap 1 activation. Given that Rap1 activation was observed in HL cells in the absence of SDF-1α, another HL cell-derived activating factor for CXCR4 signaling might activate Rap1 in an autocrine manner and facilitate the inside-out signal for integrin signaling. In summary, along with the outside-in signal from integrin signaling, the inside-out signal for integrin signaling also appears to be strongly upregulated in the HTLV-1-infected cells of HAM/TSP patients, suggesting that this activity also functions as one of the first responses in the development of HAM/TSP (Fig. 2).
Other HTLV-1-associated diseases
Several inflammatory diseases, such as uveitis, alveolitis, arthritis, mononeuropathy, myositis, inclusion body myositis, and Sjögren syndrome, occasionally occur in conjunction with HAM/TSP or HTLV-1 infection. 37 The pathogenesis of these inflammatory diseases in conjunction with an HTLV-1 infected status is still obscure. However, pathological studies in HTLV-1-associated myositis detected HTLV-1 proviral DNA and HTLV-1 tax expression in infiltrating cells in the perimysium but not in the muscle fiber. 38 In addition, HTLV-1 tax-specific CTLs were also detected in the perimysium. 39 These findings appear similar to the neuropathological findings of the spinal cord in HAM/TSP patients, suggesting that a bystander mechanism may also be at work in the muscle tissues. Therefore, HTLV-1-infected CD4+ T cells may function as the first responders even in the pathological process in the muscle tissues.
In our studies of HAM/TSP patients, we found that Sjögren syndrome occurs at a very high frequency, in 6 out of 10 HAM/TSP patients. 40 Furthermore, immunohistological studies revealed T cell infiltration in the labial salivary glands of all examined HAM/TSP patients, 40 suggesting that the T cells of HAM/TSP patients have transmigrating activity into the salivary glands, even if its occurrence is not severe enough to induce the pathological changes that cause Sjögren syndrome. Ectopic germinal center (GC) formation in the labial salivary glands, which might be associated with autoantibody production and salivary destruction, is one of the characteristic pathological features of Sjögren syndrome. 41 We analyzed the existence of ectopic GCs in the labial salivary glands of HTLV-1-seropositive Sjögren syndrome patients, HAM/TSP patients with Sjögren syndrome, and HTLV-1-seronegative Sjögren syndrome patients and found that the amount of ectopic GC was significantly lower in HTLV-1-seropositive Sjögren syndrome patients as compared with that in HTLV-1-seronegative Sjögren syndrome patients. Interestingly, ectopic GCs were not detected in all nine of the HAM/TSP patients with Sjögren syndrome. 42 This finding indicates that Sjögren syndrome with HAM/TSP is not based on GC formation but might be based on T cell infiltration into the salivary glands. Thus, these results suggest that the main pathogenesis of Sjögren syndrome occurring in conjunction with HAM/TSP is not simply a “combination of both diseases” but might instead be based on the increased transmigrating activity into the tissues of HTLV-1-infected CD4+ T cells in HAM/TSP patients as described above.
Conclusions
In conclusion, we propose the importance of integrin signaling activation in the HTLV-1-infected CD4+ T cells as the first response in the development of HAM/TSP. This activity is composed of both outside-in from integrin signaling and inside-out for integrin signaling, which are induced by the activation of small GTPases, such as Cdc42 and Rap1, respectively; furthermore, this activity appears sufficient for allowing HTLV-1-infected CD4+ T cells to function as the first responders in the development of HAM/TSP (Fig. 2). Lastly, the activation of integrin signaling may also act as the first trigger in the development of other HTLV-1-associated diseases in HAM/TSP patients. Therefore, therapeutic approaches toward downregulation of the activity of these small GTPases might contribute to a successful treatment against HAM/TSP.
Footnotes
Acknowledgments
We thank Katie Oakley, PhD, from Edanz Group for editing a draft of this article. This work was supported in part by the Japan Agency for Medical Research and Development (AMED), a Health and Labor Sciences Research Grant on Intractable Diseases (“Neuroimmunological Diseases” and “Clinical research for HAM/TSP toward identification of new biomarkers and establishment of therapeutic strategies”) from the Ministry of Health, Labor and Welfare of Japan, and a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology in Japan.
Author Disclosure Statement
No competing financial interests exist.