
Abstract
We reported a simple genome editing approach that can generate human immunodeficiency virus-1 (HIV) coreceptor defective cells, which may be useful for latent viral eradication treatment. Samples of bone marrow leftover after diagnostic procedures and crude bone marrow from aviremic HIV patients were subjected to zinc finger nuclease-mediated stop codon insertion into chemokine receptor 5 (CCR5) loci. Locked nucleic acid-based polymerase chain reaction was used to estimate the amount of insertion in the expandable CD34+ cells. The results showed that about 0.5% of CD34+ cells carried stop codon insertions in CCR5 loci. Cells edited using this simple protocol have the potential to be infused back into the bone marrow.
Introduction
E
Chemokine receptor 5 (CCR5) is an important coreceptor that allows HIV to enter targeted cells. Natural dysfunction of CCR5 due to a homozygous 32-bp deletion in CCR5 is known to impart some level of resistance to HIV infection and to slow the rate of progression. 6,7 This variant is believed to have naturally been selected among Caucasian populations of Eastern Europe as a result of pandemics of vaccinia and/or small pox. 8 This was demonstrated by HIV eradication in a patient with chronic HIV who received a CCR5-defective bone marrow transplantation (BMT) for the treatment of acute myeloid leukemia. 9,10
Subsequent attempts to replicate BMT using CCR5delta32 donors as a strategy to cure HIV have, however, been unsuccessful. Following this discovery, researchers have attempted to find ways to provide patients with their own lymphocytes, modified to resist re-emerging virus; disruption of CCR5 has been attempted using various genome editing techniques and in various cells. 11 –24 Most of the work relies on an error-prone DNA repair method, nonhomologous end joining, that tends to generate uncontrollable insertion/deletion mutations.
In 2015, we demonstrated the possibility to integrate a stop codon into the CCR5 locus via zinc finger nuclease (ZFN)-mediated homologous recombination (HR) in bone marrow-derived mesenchymal stem cells, which can directly be converted to hematopoietic stem cell (HSCs)-like cells. 25 However, to date, direct cell conversion to HSCs has not been achieved in clinical practice. 26,27 In this study, we tested the efficacy of HR-mediated stop codon insertion into bone marrow HSCs, which are better suited to self-transplantation.
Materials and Methods
Patients
All study protocols were approved by the Ethics Committee of Lerdsin General Hospital. Informed consent was obtained from all subjects. Aviremic chronic HIV patients (HIV-1 RNA under 50 copies/mL of plasma) who were clinically stable and had not changed ARV protocol for more than 6 months, who were without any coexisting disease, and who regularly followed up at Lerdsin General Hospital were invited to donate bone marrow samples. Bone marrow leftover from diagnostic aspiration (non-HIV patients) was obtained in agreement with the attending physician. Bone marrow aspirations were performed using the standard procedure by experienced clinicians.
Genome editing and crude bone marrow expansion
CCR5-specific ZFN was purchased from Sigma Aldrich (St. Louis, MO). This ZFN was designed to target the third exon of CCR5. 11,25 A pair of ZFN mRNAs were synthesized using the MassageMax T7 mRNA Transcription and Poly(A) Polymerase Tailing Kit (Epicentre; Madison, WI). The d-stop donor that carried the universal stop codon “TAGATAGTTAG” flanking the region −733 bp upstream of the left-hand ZFN-binding site to 1,038 bp downstream of the right-hand ZFN-binding site has previously been described. 25 Nucleated cells were isolated from 3 to 5 mL of heparinized bone marrow and were transfected with ZFN mRNA and d-stop donor plasmid. Cells were plated on Matrigel-coated flasks and cultured in CD34+ culture medium (Iscove's modified Dulbecco's medium supplemented with 10% fetal bovine serum and 100 units/mL penicillin and G-streptomycin (Gibco; Grand Island, NY), 50 ng/mL Flt3, 50 ng/mL thrombopoietin, and 50 ng/mL stem cell factor (R&D Systems)) for 5 days. Flow cytometry for CD34 was performed at day 0 and 5. CD34-positive cells were isolated from expansion cells at day 5 using a magnetic-activated cell sorting system (Miltenyi Biotec, Auburn, CA) according to the manufacturer's protocol (Fig. 1a.).
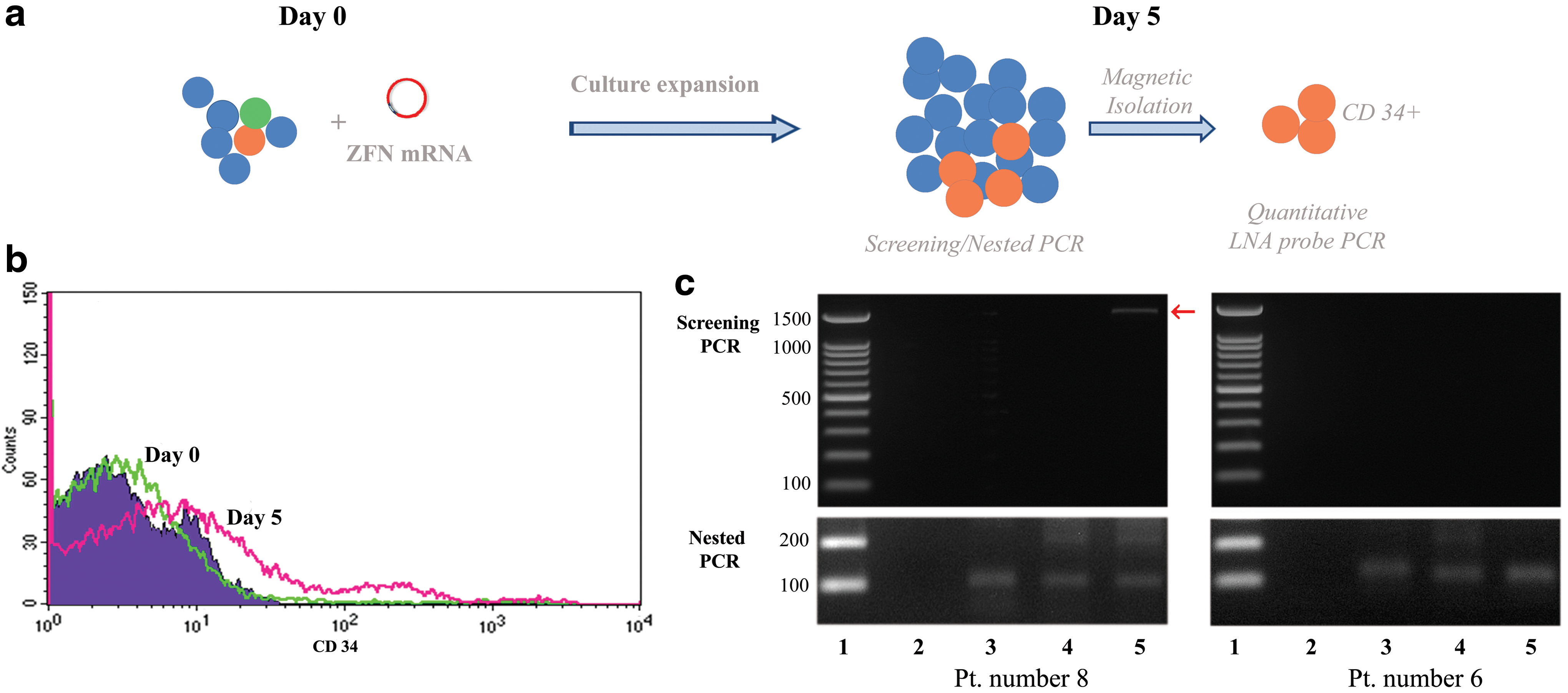
Experimental protocol and PCR results.
Polymerase chain reaction and locked nucleic acid probe polymerase chain reaction
The primer sequence for polymerase chain reaction (PCR) with the locked nucleic acid (LNA) probe was as follows: Forward 5′-TTAAAAGCCAGGACGGTCAC-3′ and Reverse 5′-TGTAGGGAGCCCAGAAGAGA-3′. LNA probes (sequence 56-FAM/AGT+TAG+CCA+TAC+AGTCCAGT/36-TAMSp) for detecting stop codon insertion were synthesized from Exiqon (Exiqon, Inc., Woburn, MA). The extracted genomic DNA was used to perform real-time PCR using a Platinum® Quantitative PCR SuperMix-UDG (Invitrogen, Carlsbad, CA).
Results
Isolation of bone marrow nucleated cells
Nucleated cells were isolated from five leftover bone marrow aspiration specimens of non-HIV patients and four fresh aspirations from aviremic patients with HIV by Ficoll–Hypaque. Flow cytometry analysis showed that the average percentage of CD34+ cells was 2.08 ± 1.66 in leftover bone marrow and 1.29 ± 0.77 in samples from aviremic HIV-positive volunteers.
Proliferation of edited bone marrow cells
Within 20 min after bone marrow aspiration, 1 × 106 isolated nucleated cells were transfected with ZFN mRNA and D-stop plasmid by nucleoporation as previously described. 25 To test the survival and proliferation of the edited cells, we plated cells into CD34+ culture medium for 5 days. The total cell number was counted, and the percentages of CD34-positive cells were determined by flow cytometry. The results showed that 5-day culture in reprogramming media increased the total cell number by ∼175% and increased the average CD34+ fraction by 11.8-fold (Fig. 1b.).
Screening and nested PCR
Genomic DNA was extracted from expansion cells. PCR amplification using screening primers and confirmation using the nested primers showed that four of five leftover bone marrow samples and three of four samples from aviremic HIV patients showed insertion of the stop codon sequence in CCR5 loci in expansion cells. (Fig. 1c.).
LNA probe PCR for quantitation of copy number
CD34+ cells were isolated from expanded nucleated cells by magnetic isolation. DNA extracted from expanded CD34+ cells was next subjected to testing with LNA probe PCR. The standard curve was constructed using dilution of the d-stop primer. The average stop-codon copy number was 128 ± 209 copies per 105 CD34+ cells (134 ± 250 and 57 ± 90 copies per 105 CD34+ cells for leftover bone marrow and aviremic HIV-positive subjects, respectively) (Table 1). In aviremic HIV-positive subjects, the calculated quantity of expandable CD34+ that harbored the stop-codon (presumed to indicate heterozygosity) was ≈2,300 cells per 105 of starting CD34+ cells.
NHL, non-Hodgkin lymphoma; PCR, polymerase chain reaction; HIV, human immunodeficiency virus-1.
Discussion
Although the debate on the optimal latent viral eradication mechanism continues, allogenic BMT of CCR5 -defective HSCs is the only treatment shown to cure an HIV infection. Genome editing of patients' own cells is a potential way to source CCR5-defective HSCs without immunologic incompatibility barriers. However, simple cell replacement with HIV-resistant cells might not lead to a complete cure, 28,29 because in chronic infection, the virus has already integrated its DNA into the genome of latent cells. Recently the “shock and kill” concept, in which the latent integrated provirus is intentionally activated, has been postulated as a strategy for reservoir eradication. 5,30 –32 As part of this strategy, virus-resistant cells may provide benefit by reducing de novo infection during proviral activation. In addition, HIV-resistant cell replacement also has potential among uninfected persons as a prophylactic treatment against HIV-1. These maneuvers theoretically can prevent primary HIV-1 infection among uninfected persons who subsequently become exposed. In this work, we showed the efficacy of ZFN-mediated stop codon integration into CCR5 loci in potential patients' own cells, using a simple protocol. Without the need for cell isolation or complex cell stimulation, the edited cells survived and showed a considerable proliferation rate. At day 5 of culture expansion, we observed that ∼50 cells per 105 CD34+ cells harbored a stop codon at the CCR5 locus. This number may be relatively low compared to the required dose for entire bone marrow regeneration in clinical practice, however, clonal generation of edited self-cells is different to bone marrow replacement. 33 One interesting approach is to directly inject these cells back to the bone marrow. Recent studies suggested that direct cell infusion into the marrow, that is, intrabone marrow injection, showed effective engraftment at lower doses. 34 –38 Our protocol requires only mRNA and a promoter-less plasmid. None of the materials in the process was contaminated by bovine or other animal substances. The overall bone marrow aspiration and editing process required less than 30 min, allowing the edited crude bone marrow cells to be injected back to the bone marrow in the same setting as the aspiration. In addition, the process may be repeated on multiple rounds and visits to generate the desired amount of CCR5-defective cells.
In summary, we reported a method for ZFN-mediated stop-codon integration into CCR5 loci of HIV-positive patients' bone marrow. Cells in the edited CD34+ fraction survived with a substantial proliferation rate. We therefore postulate that these CD34+ cells should expand if they are immediately injected back into the bone marrow. This approach might be beneficial for HIV treatment.
Footnotes
Acknowledgments
We thank Prof. Pokrath Hansasuta (Division of Virology, Department of Microbiology, Faculty of Medicine, Chulalongkorn University, Bangkok) for his helpful advice. We are grateful to Dr. Ryoji Sassa (Okasaki, Nagoya) for his generous support. This work was supported by Lerdsin Foundation and SS Manufacturing. Kamanotip Chaikomon was supported by the 90th and 100th Anniversary of Chulalongkorn University Funds.
Author Disclosure Statement
No competing financial interests exist.