
Abstract
The cytoplasmic tail (CT) of the HIV-2 envelope glycoprotein (Env) includes amino acid (aa) sequences that are similar to lentiviral lytic peptides (LLP) described in other lentiviruses. Within the putative LLP-2 region, we previously observed insertions of 3 or 7 aa in sequences deduced from plasma viral RNA of symptomatic HIV-2-infected individuals. Based on these observations, we reproduced the insertions in a molecular clone to assess their impact on replicative fitness and cell death in vitro. Using a molecular clone of the HIV-2ROD reference strain, site-directed mutagenesis experiments allowed the generation of plasmids with the insertion L791 TAI or L791 QRALTAI in the Env protein. The clone with 7 aa insertion enhanced viral release 8 to 11 times in infected T cells and cell viability was impaired by more than 20%, compared with the wild-type HIV-2ROD virus. The effect of the 3 aa insertion was milder, with a nonsignificant trend to enhance viral replication and cell death compared with the wild-type virus. Interestingly, the insertions in the Env proteins did not induce a significant increase of viral infectivity, as revealed by the infectivity assay using TZM-bl cells. The insertions in the Env CT observed in vivo from disease progressors may, therefore, be involved in the higher viral load observed in these individuals. This study may open the way to the development of a prognostic marker related to the HIV-2 infection progression.
T
The CTs of HIVs are dispensable for binding and fusion activities but these domains are essential for antero- and retrograde trafficking, viral assembly, incorporation in newly synthesized virions, and conformational structure of Env glycoprotein. Interestingly, Env glycoprotein has also been reported to impact viral replication and release via, for instance, activation of NF-κB pathway and antagonism of BST-2/Tetherin in the case of HIV-2. 4,5
HIV-2 infection is characterized by lower transmission rates, lower plasma viral loads, and better immune control compared with HIV-1 disease. In contrast to HIV-1 patients, the majority of HIV-2 patients do not progress to AIDS and 70%–80% are called long-term nonprogressors. 6,7 In a previous study, our research group observed insertions of 3 or 7 amino acids (aa) in env sequences deduced from plasma viral RNA of several HIV-2-infected individuals. A cohort of 72 HIV-2 patients from Belgium and Luxembourg was established and among these, 23 patients were selected because of their detectable viral load (>100 copies/mL). Among these 23 progressors, the insertions of 7 (QRALTAI) or 3 (TAI) amino acids were observed in the Env CT from four different patients (17%). Noticeably, the patients infected with virus harboring the 7 aa insertion in the Env CT displayed the highest viral load of this cohort (>35,000 copies/mL). 8 The only exception was a patient infected with a virus harboring an insertion of 7 aa LRALTAT in Env CT and showing a low viral load (405 copies/mL). Patient characteristics and available clinical information are summarized in Table 1.
ART, antiretroviral therapy; N.A., not available.
To confirm the impact of these insertions on viral replication in vitro, we reproduced the insertions by site-directed mutagenesis using a molecular clone of HIV-2ROD encoding a complete envelope cytoplasmic domain (pKP59-HIV-2ROD, 9 GenBank: M15390.1). We generated two different mutants of HIV-2ROD: a clone containing the insertion L791 TAI and another clone containing the insertion L791 QRALTAI in the envelope CT (Fig. 1A). The sequences of primers used in inverse PCR were as follows: F Env 7aa 5′-cag aga gca cta aca gca atc aga gac tgg ctg aga ctt aga aca gcc ttc ttg-3′ or F Env 3aa 5′-aca gca atc aga gac tgg ctg aga ctt aga aca gcc ttc ttg caa tat ggg tgc-3′ (inserted nucleotides are in italic) and R Env: 5′-gag att ctg gta gat gag ttg gag ggt cag gaa gct cct gga tag-3′. Molecular clones were transfected in HEK293T cells, and the produced viral particles were purified by using a sucrose cushion 3 days post-transfection. We used our quantitative reverse transcription PCR (RT-qPCR) assay to quantify the number of viral RNA copies from a fraction of viral stocks following a protocol previously described. 9 H9 lymphocyte T cells were infected with wild-type HIV-2ROD (WT), HIV-2ROD Env L791 TAI, or HIV-2ROD Env L791 QRALTAI viruses at a multiplicity of infection of three (MOI = 3, based on the number of viruses calculated by the RT-qPCR assay). After 3 h of infection, inocula were removed; infected cells were carefully washed three times with PBS and then incubated in complete RPMI culture medium. At 3 and 6 days postinfection, virus-containing supernatants were harvested and the number of viral copies released from infected cells was determined by RT-qPCR assay. We observed that viral replication and release ability of the HIV-2ROD Env L791 QRALTAI virus were enhanced by 8- to 11-fold compared with that of wild-type HIV-2ROD virus (Fig. 1B). We also investigated the expression of viral proteins by western blot analysis to check the enhanced viral production in H9 cells. To this end, H9 cells were infected with the different viruses at a multiplicity of infection of three (MOI = 3) and Maraviroc and bicyclam AMD-3100 (a CCR5 and a CXCR4 competitive antagonist, respectively) were added the next day. Two days postinfection, cells were harvested and lysed. The cell lysates were clarified by centrifugation at 20,000 × g for 15 min at 4°C, and protein sample loading buffer was added. Samples were boiled, resolved by electrophoresis on 12% polyacrylamide gels, and transferred to nitrocellulose membranes. Membranes were blocked with 5% BSA in TBS supplemented with 0.1% of Tween-20 for 1 h and probed with a rabbit polyclonal anti-HIV-2 gp120 antibody (AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH), a mouse monoclonal anti-HIV-2 gag p26 antibody (USBiological), or a rabbit monoclonal GAPDH antibody to detect endogenous proteins (Cell Signaling Technology). As shown in Figure 1C, HIV-2 Env and p26 expression levels were increased in H9 cells infected with HIV-2ROD Env L791 QRALTAI viruses compared with HIV-2ROD WT.

Impact of 3 and 7 amino acid insertions in the HIV-2 envelope cytoplasmic tail on viral replication and release.
It is important to note that these insertions also modify the second exon of the HIV-2 rev gene. Since the Rev protein is implicated in viral RNAs exportation from the nucleus to the cytoplasm, the insertion of 7 aa (showing the most important effect on the HIV-2 replication) may conceivably enhance viral RNAs transport, thereby improving virus production. To test this hypothesis, we constructed molecular clones for HIV-2ROD Δrev and HIV-2ROD Δrev Env L791 QRALTAI, and we obtained vectors encoding the synthetic gene of wild-type rev or rev with the corresponding 7 aa insertion (Fig. 1D; Eurofins Genomics, Germany). The different molecular clones for HIV-2ROD Δrev were transfected alone in HEK293T cells or transcomplemented with plasmids encoding wild-type Rev or Rev with 7 aa insertion. Forty-eight hours later, accumulated viral particles in culture supernatants were quantified by RT-qPCR assay. Of note, HEK293T cells transfected with the molecular clone for HIV-2ROD Δrev generated nearly no viral particles, given the absence of Rev proteins. Interestingly, the production of HIV-2ROD Δrev (encoding wild-type Env protein) transcomplemented with Rev with the 7 aa insertion did not enhance viral production and release, whereas the production of HIV-2ROD Δrev Env L791 QRALTAI viruses along with the expression in trans of wild-type Rev boosted viral production and release (Fig. 1E). These results demonstrate that the enhanced viral production is only due to the modification in the HIV-2 Env sequence, and not in the Rev sequence. Further, these results corroborate previous observations that all the HIV-2 patients infected with HIV-2 virus harboring this 7 aa insertion QRALTAI in the Env CT exhibit a high viral load and disease progression. 8
Given that insertions are located in the putative Env LLP-2 region of HIV-2, we next sought to determine whether cell viability could be impacted after expression of the Env CT mutants. Indeed, it has been reported in HIV-1 that the LLPs, three conserved amphipathic α-helical segments in the Env CTs of HIVs, induce apoptosis or cytolysis of infected cells by disturbing cell membrane integrity. 3,10 Therefore, at day 4 postinfection, we evaluated cell viability of the infected H9 T cells by using a CASY cell counter that was able to discriminate living cells from dead ones. Whereas 77% of cells infected with HIV-2ROD WT virus were still living, only 55% of cells infected with HIV-2ROD Env L791 QRALTAI virus were living cells. In the case of HIV-2ROD Env L791 TAI, there were no significant differences regarding cell viability compared with the infection with wild-type HIV-2ROD virus (Fig. 2A). Thus, the 7 aa insertion in the HIV-2 Env CT causes a higher mortality, possibly due to the presence of additional charged residues in the putative LLP-2 region or may be due to enhanced viral transmission followed by cell death.
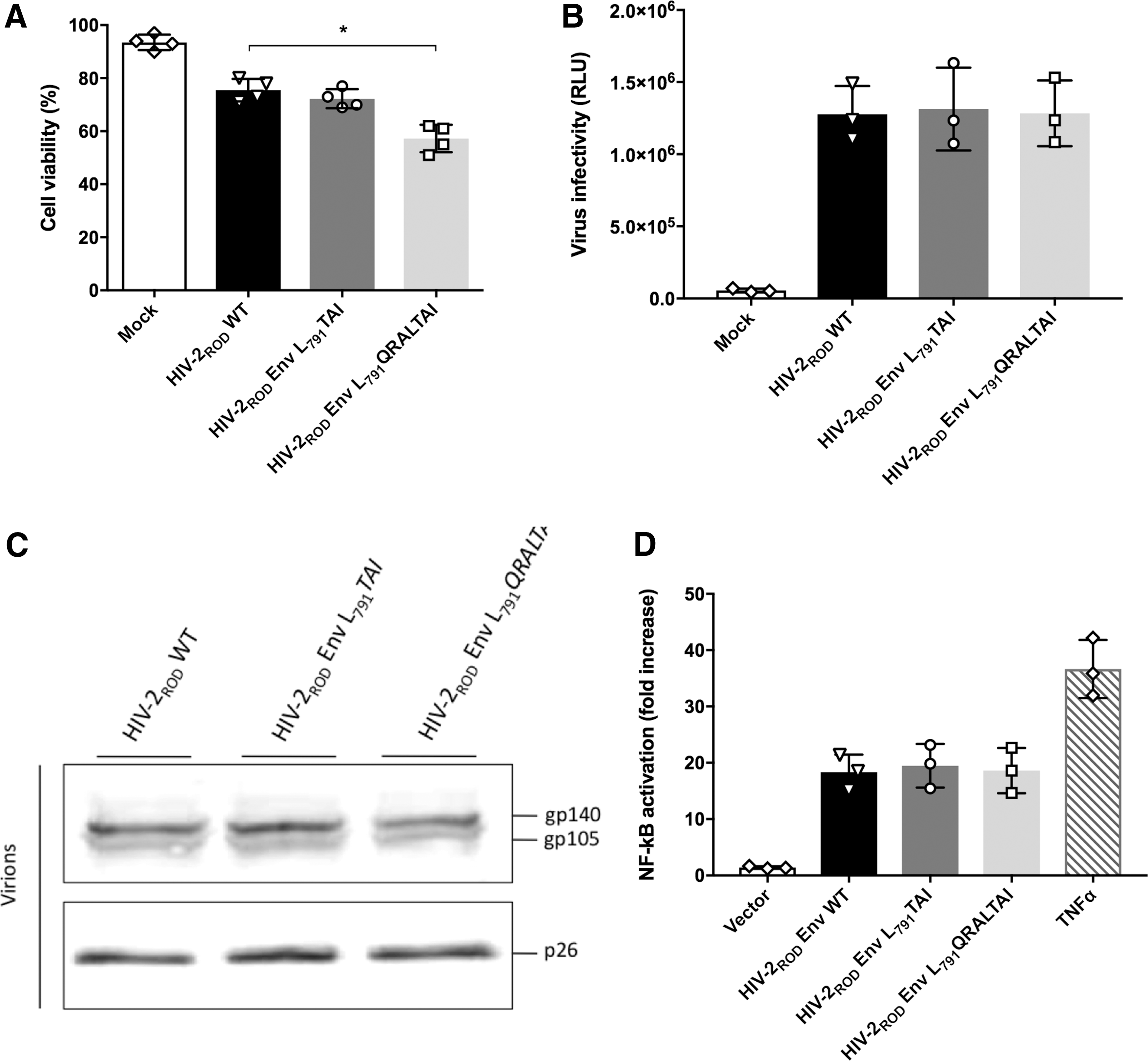
Impact of 3 and 7 amino acid insertions in the HIV-2 envelope cytoplasmic tail on cell viability, viral infectivity, Env incorporation, and NF-κB activation.
To better define mechanisms involved in the enhanced viral replication and release observed with the HIV-2ROD Env L791 QRALTAI virus, we hypothesized that the 7 aa insertion can promote fusogenicity and quantity of Env glycoproteins at the virion surface and, consequently, may favor binding and entry processes during early steps of infection. Therefore, we tested viral infectivity of HIV-2ROD Env L791 TAI and HIV-2ROD Env L791 QRALTAI viruses and we compared this ability with that of wild-type HIV-2ROD virus. We used TZM-bl reporter cells to test viral infectivity of the three different viruses. These cells contain a luciferase reporter gene under the control of an HIV long terminal repeat (LTR) promoter and can be used to assess viral infectivity. TZM-bl cells were seeded in a 24-well plate and infected with wild-type HIV-2ROD, HIV-2ROD Env L791 TAI, or HIV-2ROD Env L791 QRALTAI viruses at a multiplicity of infection of three (MOI = 3) during 3 h. After this step of infection, inocula were removed, adherent TZM-bl cells were gently washed three times with PBS, and fresh culture medium was added to the wells. To avoid multiple cycles of infection and test only viral infectivity capacity, we added Maraviroc and bicyclam AMD-3100 in the culture medium, and we performed luciferase reporter assays 28 h later. As presented in Figure 2B, viral infectivity of the three viruses remained equal, suggesting that the insertions did not promote infectivity. However, insertions in the Env CT may impede or promote the Env incorporation per virion, thereby explaining the improved viral fitness of the HIV-2ROD Env L791 QRALTAI viruses. To observe and compare the Env incorporation ratio of these different viruses, we performed a western blot of the infection-derived viral fraction. Of note, we used equivalent amounts of virus to complete the western blot analysis. No real differences were observed in Env incorporation ratios, although a slight decrease was seen with HIV-2ROD Env L791 QRALTAI viruses (Fig. 2C). As fusogenicity and Env incorporation may compensate each other, it could still be that a lower Env incorporation may balance a higher fusogenicity to maintain a similar viral infectivity.
Our results suggest that the improvement in viral replication induced by the insertion of 7 aa in Env CT is due to a postintegration mechanism. Indeed, Env glycoproteins may impact and promote viral replication. In respect to this, we recently reported an important role of HIV-2 Env in activation of the NF-κB signaling pathway and this effect can induce LTR-driven proviral genes transcription. 5 Therefore, it is tempting to speculate that insertions in Env CT could lead to a higher activation of NF-κB, resulting in a higher viral production in infected cells. We used our HEK293T reporter cell line stably expressing the luciferase gene under the control of NF-κB binding sequences to assess the NF-κB activation ability of the different Env glycoproteins. 5 We cloned wild-type HIV-2ROD Env, Env L791 TAI, or Env L791 QRALTAI coding sequences into pcDNA3.1 expression vector and we transfected these plasmids in HEK293T-NF-κB cells. Twenty-four hours later, we performed a luciferase reporter assay, allowing us to evaluate the effect of these insertions on NF-κB activity. We observed no increased capability of the mutated Env glycoproteins in activation of NF-κB (Fig. 2D).
Overall, our in vitro results validate in vivo observations that the 7 aa insertion in HIV-2 Env CT induces enhanced viral replication and cell mortality but the mechanisms underpinning this viral fitness and phenotype remain to be defined. We believe that this study provides new insights into the role of the Env CT in the HIV-2 replication and pathogenesis.
Footnotes
Acknowledgments
The authors thank Dr. Patrick Goubau for the fruitful discussions concerning this study and for the proofreading of the article. They also thank Dr. Isabel Barahona (Instituto Superior de Ciências da Saúde-Sul, Portugal) for providing the pKP59-HIV-2ROD molecular clone. Finally, they acknowledge Dr. Robert Gallo and Dr. John C. Kappes for providing the H9 cells and the TZM-bl cells, respectively (AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH). They also acknowledge Dr. Raymond Sweet for providing the rabbit antiserum to HIV-2 gp120 (AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH).
Author Disclosure Statement
No competing financial interests exist.