
Abstract
Sexual contact is the main route of transmission of HIV type 1 (HIV-1) pandemic in China. New unique recombinant forms (URFs) were increasingly characterized in sexual populations in recent years. We reported two URFs identified from two male HIV-1-positive patients who were infected through sexual exposure in Shaanxi, China. Phylogenetic analyses and bootscan analyses revealed that the near full-length genomes of the two URFs were having four recombinant breakpoints, with two CRF07_BC fragments inserted into CRF01_AE backbones. The CRF01_AE fragments of the two URFs clustered with previously reported a cluster 5 lineage of CRF01_AE. The four recombinant breakpoints of the two URFs were quite similar. The emergence of the new CRF01_AE/07_BC URFs indicated the complexity genetic variability and the active epidemic of HIV-1. Much more attention should be paid to monitor the emergency of recombinant strains.
T
The dominant routes of transmission of HIV/AIDS epidemic in China have shifted from contaminated former paid blood donation and injecting drug use to sexual contact. 4 From Jan to Oct in 2017, about 97.3% newly diagnosed HIV-1 cases in Shaanxi province were infected through sexual exposure. 5 Risky sexual behavior of sexual active populations, such as having multiple sexual partners and unprotected sexual intercourse, created the opportunities for dual and multiple HIV infections. New HIV-1 CRFs had been continuously reported among homo- or heterosexually infected population in recent years. 6,7 CRF01_AE and CRF07_BC are the two predominant strains among MSM and heterosexuals in China. 8,9 Several CRF01_AE/07_BC URFs were identified across China, including Shaanxi province. 10 In the present study, two new CRF01_AE/07_BC recombinant forms were characterized to have four similar breakpoints in common, which were different from previously reported CRF01_AE/07_BC recombinants.
Whole blood samples were collected from two male HIV-positive patients, SN139 from Xianyang during an HIV-1 molecular epidemiology survey in May 2015 and M62 from Xi'an during a longitudinal observational study among MSM in November 2015. Xi'an and Xianyang are two cities both located in Northwestern China, with the resident population of 9,000,000 and 4,000,000, respectively. Xianyang is located a few kilometers upstream (west) from Xi'an. The informed consent was signed before the collection of samples. SN139 and M62 started to receive antiretroviral therapy (ART) in August and December 2015, separately. The CD4+T cell count of patient SN139 increased from 235 cells/μL in June 2015 at the baseline of ART to 559 cells/μL in March 2016. The CD4+T cell count of patient M62 stayed a comparable high level, 693 cells/μL in November 2015 at the baseline of ART and 740 cells/μL in December 2017. More detailed information about the two patients is summarized in Table 1. This study was reviewed by the Institutional Research Ethics Community of the Shaanxi Center for Disease Control and Prevention.
ART, antiretroviral therapy.
Near full-length genome (NFLG) sequences of both SN139 and M62 were amplified, because recombinant breakpoints were identified at the previous phylogenetic analysis of pol gene fragment. RNA was extracted from 280 μL plasma using QIAamp Viral RNA Mini Kit (QIAGEN, Valencia, CA) and then reversely transcribed into cDNA using SuperScript III Reverse Transcriptase (Invitrogen, Carlsbad, CA). NFLG was amplified using near-endpoint diluted cDNA template by nested polymerase chain reactions (nest-PCRs) with TAKARA LA Taq (TAKARA). The PCR conditions of both rounds were the same, including an initial denaturation of 94°C for 2 min followed by 10 cycles of 94°C for 10 s and 68°C for 8 min 30 s and then 20 cycles of 94°C for 10 s and 68°C for 8 min 30 s with the cumulative addition of 20 s at 68°C with each successive cycle, followed by a final extension at 68°C for 20 min. The positive PCR products were sequenced by an ABI 3730XL sequencer (Applied Biosystems, Foster City, CA), and then, the chromatogram data were assembled by Sequencher v. 5.1 (Gene Codes Corporation, Ann Arbor, MI). Phylogenetic tree and subregion tree analyses were conducted by MEGA 6.05 using the neighbor-joining method with 1,000 bootstrap replications. Similarity plot and Bootscan analyses were performed using software SimPlot v3.5.1 based on the neighbor-joining method using the Kimura two-parameter model of 500 replicates with a 300 bp window size moving in a step size of 10 bp.
The NFLG sequences of SN139 and M62 were 8991 and 8973 bp corresponding to HXB2 nucleotide positions: 622–9601 and 635–9601. In the phylogenetic tree (Fig. 1), the two URFs were clustered with YA1996,
11
which was a CRF01_AE/07_BC URF previously reported in Beijing, however, independently from CRF01_AE branch. Similarity plot (Supplementary Fig. S1) and Bootscan analyses (Fig. 2) revealed a CRF01_AE parent bone with two CRF07_BC fragments inserting into it, with the reference sequences (CRF01_AE and CRF07_BC as parents, subtype J as out-group). The genomic maps of the two URFs were generated using the recombinant HIV-1 drawing tool (

A neighbor-joining phylogenetic tree of SN139 and M62 was constructed using MEGA 6.05 with reference sequences where the solid circles represent SN1139 and M62. The stability of each node was assessed by bootstrap tests with 1,000 replicates, and the bootstrap values >70% are shown at the corresponding nodes.
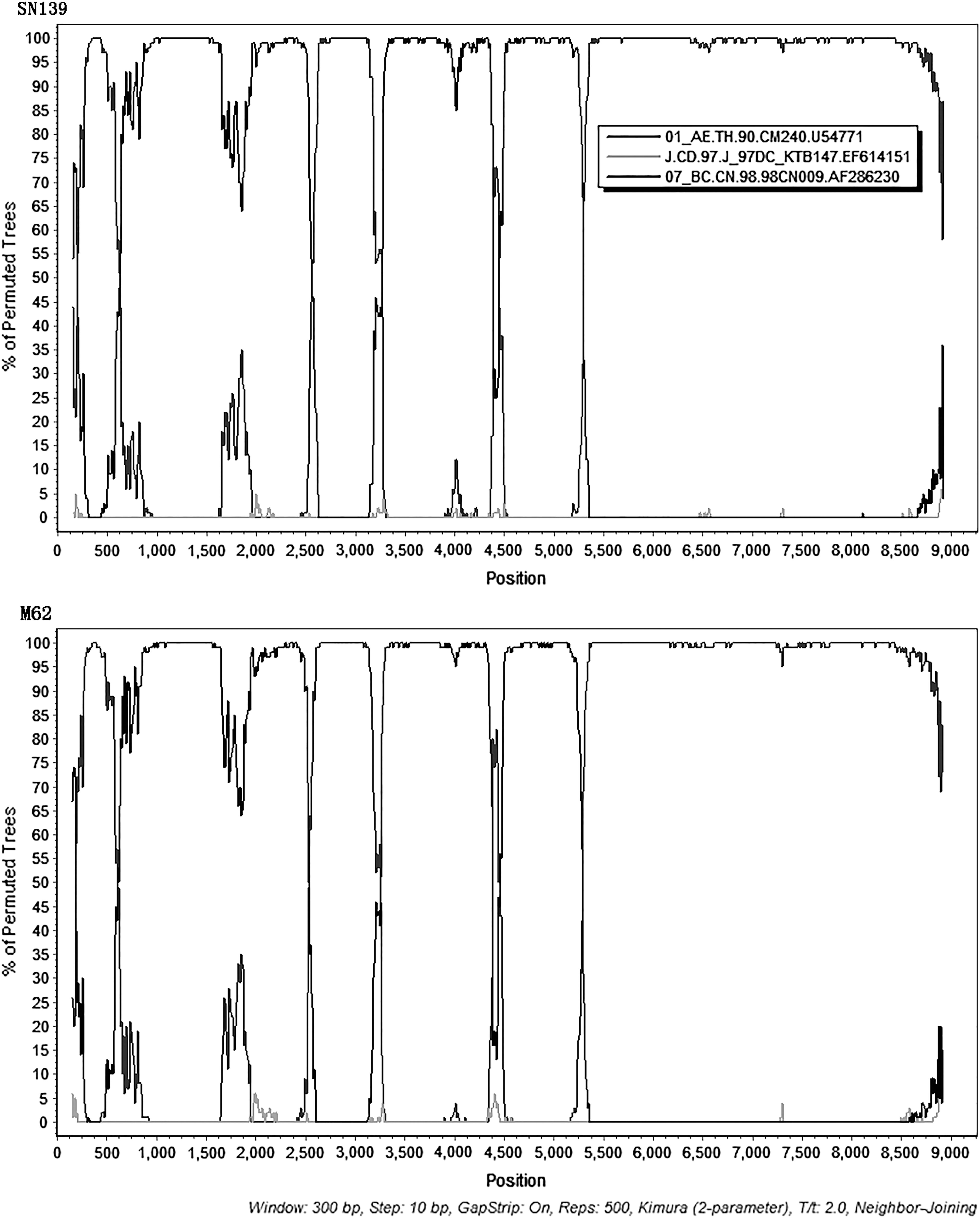
Bootscan analyses of the near full-length genome sequences of SN139 and M62. CRF01_AE (CM240) and CRF07_BC (98CN009) are used as putative parental reference sequences, and subtype J (KTB147) is used as an out-group. The parameters of bootscan analyses are as follows: window size of 300 bp; step size by 10 bp; tree algorithm, neighbor; distance model, Kimura; bootstrap replicate, 500.
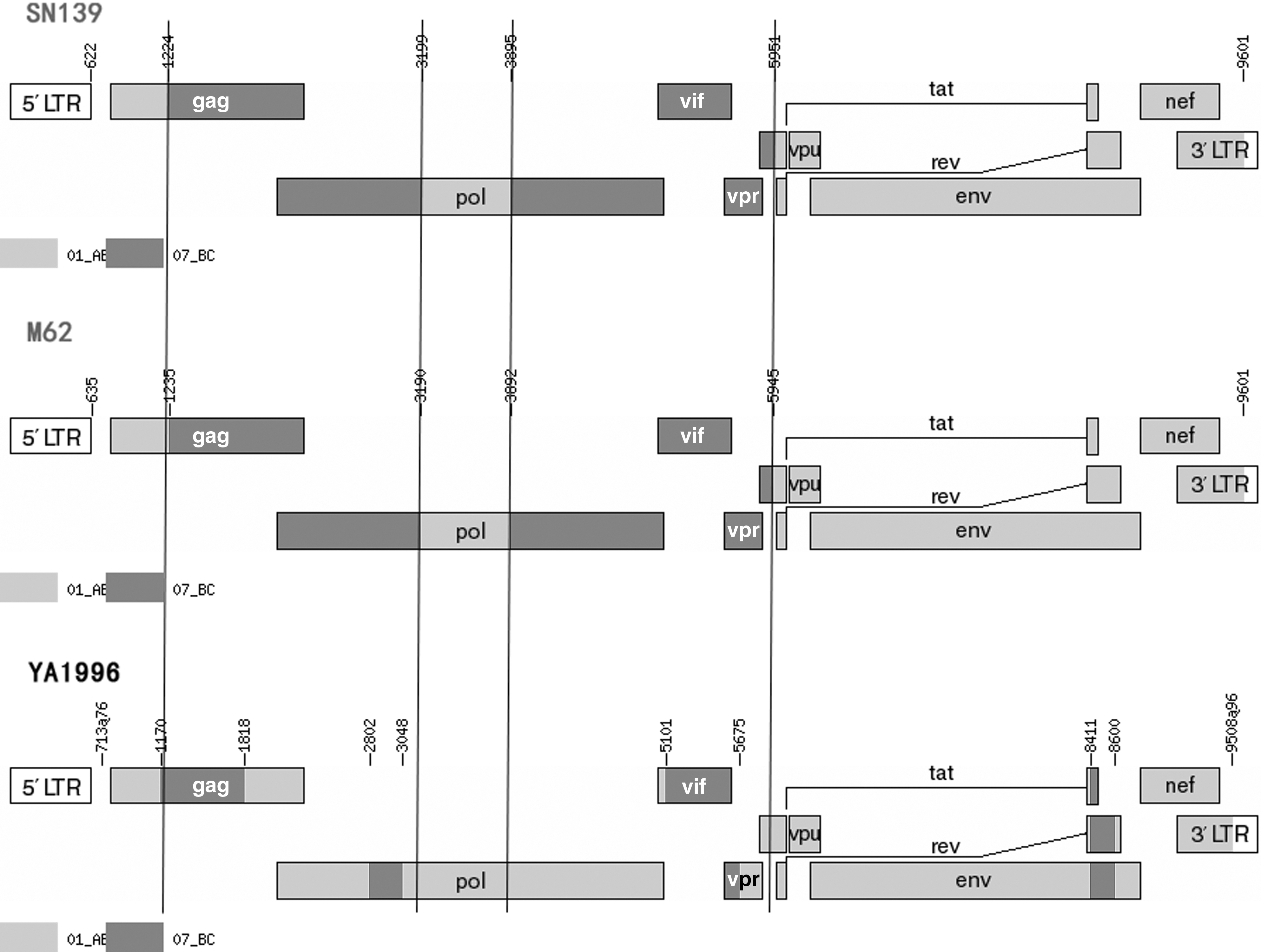
Genetic maps of SN139 and M62, created using the recombinant HIV-1 drawing tool (
The mosaic structures of the two NFLGs were displayed as follows, SN139: ICRF01_AE (HXB2 nt 622-1223), IICRF07_BC (HXB2 nt 1224-3198), IIICRF01_AE (HXB2 nt 3199-3894), IVCRF07_BC (HXB2 nt 3895-5950), and VCRF01_AE (HXB2 nt 5951-9601); M62: ICRF01_AE (HXB2 nt 635-1234), IICRF07_BC (HXB2 nt 1235-3189), IIICRF01_AE (HXB2 nt 3190-3891), IVCRF07_BC (HXB2 nt 3892-5944), and VCRF01_AE (HXB2 nt 5945-9601). Bootscan analyses revealed that the two URFs had four similar breakpoints (HXB2 nt 1229 ± 6, nt 3194 ± 5, nt 3893 ± 2, nt 5947 ± 3). One breakpoint located in gag and vpu gene, respectively, and two located in pol gene. The subregion NJ trees confirmed these breakpoints (Supplementary Fig. S2). Region I, III, and V of the two NFLGs were clustered with CRF01_AE references. Region II and IV were clustered with CRF07_BC references.
Region I, III, and V of the two NFLGs were further phylogenetically analyzed with seven distinct clusters of CRF01_AE from China. 12 Subregion trees indicated that the three CRF01_AE regions were clustered with the cluster 5 lineage, which was found among MSM and heterosexuals in the northern China. It is suggested that the parental origin of CRF01_AE regions may be from sexual populations of northern China. The two HIV-1 patients in this study were infected through sexual contact. The emergence of recombinant strains might have resulted from their unsafe sexual behavior.
In this study, we reported two novel CRF01_AE/07_BC recombinant forms from HIV-infected males through sexual transmission. The two URFs had four similar breakpoints. The emergence of new URFs indicated the complexity of local HIV epidemic among sexual populations. Since CRF01_AE and CRF07_BC were two predominant genotypes among risk groups in China, 8 recombinant forms of CRF01_AE/07_BC strains could be much more diverse than those that had been characterized. The molecular surveillance of HIV-1 diversity should be continuously conducted.
Sequence Data
The NFLG sequences of SN139 and M62 have been deposited in GenBank with accession numbers MH396609 and MH396608, respectively.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.