
Abstract
Factors underlying HIV acquisition in women remain incompletely understood. This study evaluated ex vivo mucosal HIV-1BaL infection (ectocervix, endocervix), T cell frequencies and phenotype (ectocervix, endocervix, peripheral blood), and HIV-1BaL-induced tissue immune responses (ectocervix) in the proliferative and secretory phases of the menstrual cycle using samples obtained from women undergoing hysterectomies. Tissue infectivity (number of productively infected explants) and infection level following 500 and/or fifty 50% tissue culture infectious dose (TCID50) HIV-1BaL challenge were similar in the proliferative and secretory phases. Although not associated with infection outcomes, higher frequencies of HIV target CD4+α4β7+ T cells, and stronger HIV-1BaL-induced proinflammatory responses were detected in ectocervix in the secretory versus proliferative phase. Independently of the cycle phase, serum E2 concentrations were inversely associated with ectocervical and endocervical tissue infection levels following high-dose 500 TCID50 HIV-1BaL challenge, with frequencies of CD4+α4β7+ T cells in endocervix, and with HIV-induced interleukin (IL)2R and IL4 in ectocervix. Although serum P4 concentrations and P4/E2 ratios were neither associated with tissue infection level nor infectivity, high P4 concentrations and/or P4/E2 ratios correlated with high frequencies of CD4+α4β7+ T cells in ectocervix, low frequencies of CD4+CD103+ blood T cells, low CD4+LFA-1+ T cells in endocervix, and high proinflammatory (IL1β, IL17, tumor necrosis factor α) ectocervical tissue responses to HIV-1BaL. The data suggest an inhibitory effect of E2 on mucosal HIV infection, provide insights into potential mechanisms of E2-mediated anti-HIV activity, and highlight P4-associated immune changes in the mucosa.
Introduction
Asubstantial body of evidence implicates endogenous sex hormones in HIV acquisition. In particular, the luteal (or secretory) phase of the menstrual cycle, characterized by high serum progesterone (P4) levels, was suggested to be a “window of vulnerability” for HIV acquisition, with dampening of immune protection in the upper and lower female reproductive tract. 1 –3
Studies in rhesus 4 –7 and pigtail 8,9 macaques demonstrated increased susceptibility to vaginal simian human immunodeficiency virus (SHIV) infection in the luteal phase and during menses. Further supporting the “window of vulnerability” hypothesis, one report utilizing an ex vivo challenge of surgical cervical tissues with HIV-1BaL demonstrated an association between productive tissue infection and secretory phase of the cycle. 10 In contrast, a recent study utilizing ex vivo challenge of ectocervical and vaginal biopsies found no differences in tissue HIV-1BaL infection, vaginal transcriptome, vaginal immune cell populations, and activation status between the phases of the cycle. 11
We recently demonstrated that inhibition of ex vivo simian human immunodeficiency virus-reverse transcriptase (SHIV-RT) infection in cervical and vaginal tissues from rhesus macaques administered NNRTI MIV-150-containing intravaginal ring (IVR) was predictive of in vivo efficacy against vaginal SHIV-RT challenge. 7,12 Studies using ex vivo cervical challenge model in humans revealed that dapivirine concentrations in tissues from women using the dapivirine IVR are associated with significantly reduced ex vivo infection, 13 supporting the results of The RING and ASPIRE pre-exposure prophylaxis studies 14,15 and pointing to the relevance of tissue challenge models for HIV transmission research.
Understanding the factors underlying vaginal HIV acquisition in women may better target prevention strategies. Building on published data, this study was designed to explore additional indices of mucosal HIV susceptibility in proliferative and secretory phases of the cycle using surgical ectocervical and endocervical explants. Although ectocervical tissue samples can be obtained both through a biopsy procedure and from hysterectomy specimens, sufficient amounts of endocervical tissues for in vitro experiments are typically obtained from hysterectomy specimens. To be able to study both mucosae, hysterectomy specimens were chosen. We investigated relationships between the phase of the cycle, estradiol (E2) and P4 concentrations and (i) tissue infection level and infectivity, (ii) frequencies/phenotype of mucosal T cells, and (iii) HIV-induced responses in the mucosa (innate mediators).
Materials and Methods
Subjects
The project was approved by the Icahn School of Medicine at Mount Sinai Program for the Protection of Human Subjects (protocol #11-01380) and The Population Council IRB; cervical tissues, blood, and urine were obtained from women undergoing hysterectomies for nonmalignant conditions (menometrorrhagia, leiomyomas, chronic pelvic pain, and pelvic organ prolapse) at Mount Sinai Hospital, the primary teaching hospital of the medical school.
Subjects were enrolled after providing written informed consent. This is a subanalysis from the 33 subjects [age range 35–53 years old; 44.91 ± 4.71 (mean ± standard deviation)] who did not use either (i) hormonal contraception and/or (ii) any hormonal treatments for gynecological conditions within the 3 months before surgery. Specifically, subjects using continuous combined estrogen/progesterone and progesterone only oral contraceptives, oral progesterone/gonadotropin inhibitors, oral progesterone, gonadotropin inhibitors, hormonal intrauterine device (IUDs), gonadotropin inhibitor/IUDs, intravaginal combined estrogen/progesterone, and long-acting injectable progestational agents were all excluded from the current analysis. Among those subjects included in this substudy, the nonhormonal contraceptive methods included Essure (n = 3), tubal ligation (n = 2), tubal ligation/condoms (n = 1), and condoms (n = 2). Twenty-five subjects did not use contraception.
Age, race, phase of the cycle, histopathology report [cervical inflammation, metaplasia (for endocervical mucosa), parakeratosis (for ectocervical mucosa)], and HSV-2 status of subjects included in individual data sets are summarized in Supplementary Tables S1–S3.
The cycle phase was determined by assessing the histopathology of Hematoxylin and Eosin-stained sections of the endometrial mucosa by board-certified diagnostic gynecological pathologists who perform this assessment routinely. The proliferative phase group (n = 14) included n = 2 subjects with weakly proliferative endometrium. The secretory phase group (n = 16) included n = 3 subjects in late and n = 1 subject in early secretory phase. One subject had undetermined phase of the cycle, one had inactive endometrium, and one had menses. Inflammation was defined as the presence of white blood cells within the cervical tissue (epithelium and stroma of the mucosa).
Blood and surgical tissue collection
Blood samples were collected right before surgery. Cervical tissues not used for clinical diagnosis, which would otherwise be discarded, were collected and released by pathologists within 1–2 h of surgery. Only endocervix and ectocervix, excluding transformation zone, were utilized. A portion of the mucosa (∼5 mm pieces of ectocervix and endocervix) was immediately frozen in liquid nitrogen for E2 and P4 concentration measurement by radioimmunoassay (RIA). The remaining mucosae were placed in RPMI 1640 (Cellgro Mediatech, Manassas, VA) on ice. All samples were transported from the Mount Sinai School of Medicine to the Population Council laboratory within 1 h of tissue release by the pathologists.
HIV-1BaL
HIV-1BaL was generated as in Villegas et al. 16 Briefly, CD8-depleted peripheral blood mononuclear cells (PBMCs) cells were activated for 6 days with 10 nM retinoic acid (RA; Sigma-Aldrich, St. Louis, MO), 20 U/mL interleukin (IL)2 (NCI BRB Preclinical Repository, Frederick, MD) and 50 ng/mL of anti-CD3 mAbs (clone OKT3; e-Bioscience, San Diego, CA) in RPMI 1640 containing 10% fetal bovine serum (FBS; Life Technologies, Grand Island, NY). Activated cells were then challenged with 103 50% tissue culture infectious dose (TCID50) of HIV-1BaL (ABI, Eldersburg, MD) per 106 cells and cultured for 7 days in the presence of RA and IL-2. The viral TCID50 was determined in TZM-bl cells. 16 A single HIV-1BaL stock (2.37 × 105 TCID50/mL TCID50) was used in the study.
Ex vivo HIV-1BaL challenge of cervical mucosa
Tissues were processed for viral challenge as described in Villegas et al. 16 Ectocervical mucosa was challenged with HIV-1BaL on the day of tissue collection, as soon as it arrived to the laboratory. Briefly, 3 × 3 mm ectocervical explants from each donor were challenged with 500 TCID50 and/or 50 TCID50 (2–3 explants per challenge dose) in complete Dulbecco's modified Eagle's medium (cDMEM) [(DMEM; Cellgro Mediatech) containing 10% charcoal-stripped FBS (Atlanta Biologicals, Flowery Branch, GA), 100 μM nonessential amino acids (Irvine Scientific, Santa Ana, CA), 100 U/mL penicillin, and 100 μg/mL streptomycin (Cellgro Mediatech)] (cDMEM) in 96-well plate. Where feasible, Lamivudine (3TC) control (10 μM) was included. The charcoal-stripped FBS was used to avoid effects of steroid hormones present in FBS.
After ∼18 h of challenge at 37°C and 5% CO2, tissues were washed to eliminate input virus. During the virus washout procedure, plates were spun and the input virus in cDMEM was carefully aspirated followed by ×3 washes in phosphate-buffered saline and ×1 wash in cDMEM. Then tissues were cultured for 14 days. Supernatants were collected at D0 (last wash in cDMEM), 3, 7, 11, and 14 days of culture. Infection was monitored by HIV-1 gag quantitative reverse transcription PCR (qRT-PCR).
Endocervical tissue explants (3 × 3 mm) had abundant mucus produced during incubation with HIV-1BaL when challenged on the day of tissue collection. Mucus can hinder HIV movement 17 and interfere with the outcomes of the challenge.
Visually, overnight (ON) incubation of endocervical explants in cDMEM at 4°C followed by washout before the challenge resulted in less mucus production. During the washout procedure, plates were spun and cDMEM was carefully aspirated to avoid loss of migratory cells, followed by × 2 washes in cDMEM. Endocervical explants were kept in the same plate throughout the experiment. Subsequent challenge with 500 TCID50 HIV-1BaL resulted in productive infection and less carryover virus evidenced by lower D0 HIV gag copy numbers as compared with the challenge on the day of tissue collection (Supplementary Fig. S1). Based on these results, we chose to challenge endocervical tissues after ON incubation at 4°C/washout.
HIV qRT-PCR
The infection was monitored in the supernatants using HIV-1 gag qRT-PCR (lower limit of quantification [LLOQ] 2,000 copies/mL) as in Villegas et al. 16 using SYBR FAST One-Step qRT-PCR (5 μL of supernatant per reaction; Kapa Biosystems, Wilmington, MA).
The primers were forward primer 5′ GGTGCGAGAGCGTCAGTATTAAG 3′ and reverse primer 5′ AGCTCCCTGCTTGCCCATA 3′. Any value below LLOQ was set to 20001/√2 = 215.86. 18 Results were analyzed using the standard curve method, using pNL(AD8) (11346; NIH AIDS Research and Reference Reagent Program, Germantown, MD). The cycling conditions were one cycle at 42°C for 5 min, one cycle at 95°C for 5 min, and 40 cycles at 95°C for 3 s, and 60°C for 20 s. Dissociation curves were generated to verify the absence of nonspecific amplification. HIV-1 gag qRT-PCR was performed using the ViiA 7 real-time PCR system (Applied Biosystems, Carlsbad, CA).
Data were analyzed using the ViiA 7 software (Applied Biosystems). SOFT and CUM endpoint analyses of infection level based on HIV-1 gag copies on days 3–14 of culture were performed. 16,19,20 SOFT is an estimation of HIV-1 gag copies at the start of the stationary phase of virus growth and CUM is the cumulative number of HIV gag copies. 19,20
Tissue infectivity
Tissue infectivity was analyzed based on productive infection of explants. The explants were categorized as not productively infected if HIV-1 gag copies in tissue culture supernatants were below LLOQ 2,000 copies/mL at days 3, 7, 11, and 14. If HIV-1 gag copies at one or more postchallenge time points (days 3, 7, 11, and 14) were above LLOQ, explants were categorized as productively infected. Subjects were stratified in two groups: (i) all tissue replicates infected (“infected”) or (ii) at least one uninfected replicate (“uninfected”).
Tissue viability
Human ectocervical and endocervical tissues without gross pathological changes were obtained from routine hysterectomies through the National Disease Research Interchange (Philadelphia, PA) and transported ON on ice in RPMI medium. The 3 × 3 mm explants were prepared. 16 Explant viability following ON incubation in cDMEM at 4°C (vs. viability on the day when tissues were received) was tested using the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay. 21
Radioimmunoassay
Serum and tissues were kept at −80°C before RIA. E2 and P4 concentrations were determined using the ImmuChem Double Antibody 125I RIA Kits for 17β-estradiol (LLOQ 10 pg/mL) and for Progesterone (LLOQ 200 pg/mL) (MP Biomedicals, LLC, NY). Any value <LLOQ was set to 101/√2 = 5.09 or 2001/√2 = 42.37 pg/mL. 18 Around 97.98 ± 13.91 mg and 90.3 ± 14.6 mg [mean ± standard error of the mean (SEM)] of ectocervical tissue and endocervical tissues (n = 33 each), respectively, were extracted with solvent and used in the RIA.
HSV-2 immunoglobulin G AB herpeselect test
Serum was kept at −80°C before being assayed by Quest Diagnostics.
Luminex
Levels of immune mediators in ectocervical tissue culture supernatants collected ∼18 h post 500 TCID50 HIV-1BaL challenge (vs. no challenge) were measured using the Human Cytokine Magnetic 25-Plex Panel Kit (Thermo Fisher Scientific, Waltham, MA). The analytes measured were: IL-1RA, IL-1β, IL-2R, IL-2, IL-4, IL-5, IL-6, IL-7, IL-10, IL-12, IL-13, IL-15, IL-17, tumor necrosis factor (TNF)α, Eotaxin, GM-CSF, interferon (IFN)-α, IFN-γ, CCL2, CCL3, CCL4, CCL5, CXCL8, CXCL9, and CXCL10. Each pair of samples (challenged and not challenged) were analyzed in the same Luminex plate.
Flow cytometry
PBMCs were isolated from whole blood using Ficoll-Hypaque density gradient centrifugation. A portion of ecto- and endocervical tissues (∼5 × 5 mm each) was digested in collagenase IV (1 mg/mL; Worthington Biochemical), hyaluronidase (1 mg/mL; Sigma), DNase I (10 U/mL; Roche), and bovine serum albumin (1 mg/mL; Calbiochem) for 1 h at 37°C. Cells were passed through a cell strainer (40 μm).
PBMCs and mucosal cells were stained with Aqua Dead Cell Stain (Thermo Fisher Scientific) and anti-CD3-V450 (clone UCHT1; BD Biosciences, San Jose, CA), -CD4-PE-Cy7 (SK3; BD), -CCR5-AF700 (HEK/1/85a; BioLegend), -CD45RA-BV605 (HI100; BD), -CCR7-PerCP-Cy5 (150503; BD), -LFA1-FITC (G43-25B; BD), -CD54 (HA58, BD), -CD103-APC (B-Ly7; eBioscience), and α4β7-PE (Act-1; NHP reagent resource) Abs. Anti-CD54 Abs were conjugated with APC-Cy7 using the APC-Cy7 Labeling Kit (Novus Biologicals). Samples were acquired on a BD LSRII Flow Cytometer and analyzed using the FlowJo 8.8.6 software.
Statistics
Mann–Whitney test was used to analyze (i) E2 and P4 concentrations, (ii) frequencies of blood and tissue T cells, and (iii) soluble mediators (HIV-1BaL induced/baseline fold change) in proliferative and secretory phases of the cycle.
Log-normal generalized linear models were utilized for analysis of (i) cervical tissue infection level (log-transformed SOFT and CUM values) following 500 and 50 TCID50 challenge, (ii) relationships between serum hormonal concentrations, P4/E2 ratios, and infection level (SOFT and CUM values) accounting for 2–3 tissue replicates used in the ex vivo challenge assay. The revealed significant associations between serum E2 and tissue infection levels were adjusted for potential confounding parameters: age, race, endometrial histology, metaplasia (for endocervix), parakeratosis (for ectocervix), cervical inflammation, and HSV-2 status.
Mixed effects logistic regression analysis was performed to analyze relationships between hormonal concentrations or P4/E2 ratios and tissue infectivity.
Chi-Square statistics, including Fisher's exact test, was calculated to assess the effect of challenge dose and phase of the cycle on tissue infectivity.
Linear regression analysis was used to determine relationship between serum hormonal concentrations, P4/E2 ratios, and (i) subject age, (ii) T cell frequencies (log-transformed), and (iii) soluble mediators (HIV-1BaL induced/baseline fold change). Grubbs' test was used to identify outliers.
Results
Tissue infection following 500 and 50 TCID50 HIV-1BaL challenge
The ectocervical explants were challenged with 500 and/or 50 TCID50 and endocervical explants were challenged with 500 TCID50 of HIV-1BaL due to limited amount of tissue. Examples of HIV-1BaL growth kinetics over the 14 days of culture are shown in Figure 1. Moving forward, all infection results are expressed using the SOFT and CUM endpoints encompassing days 3–14 data.
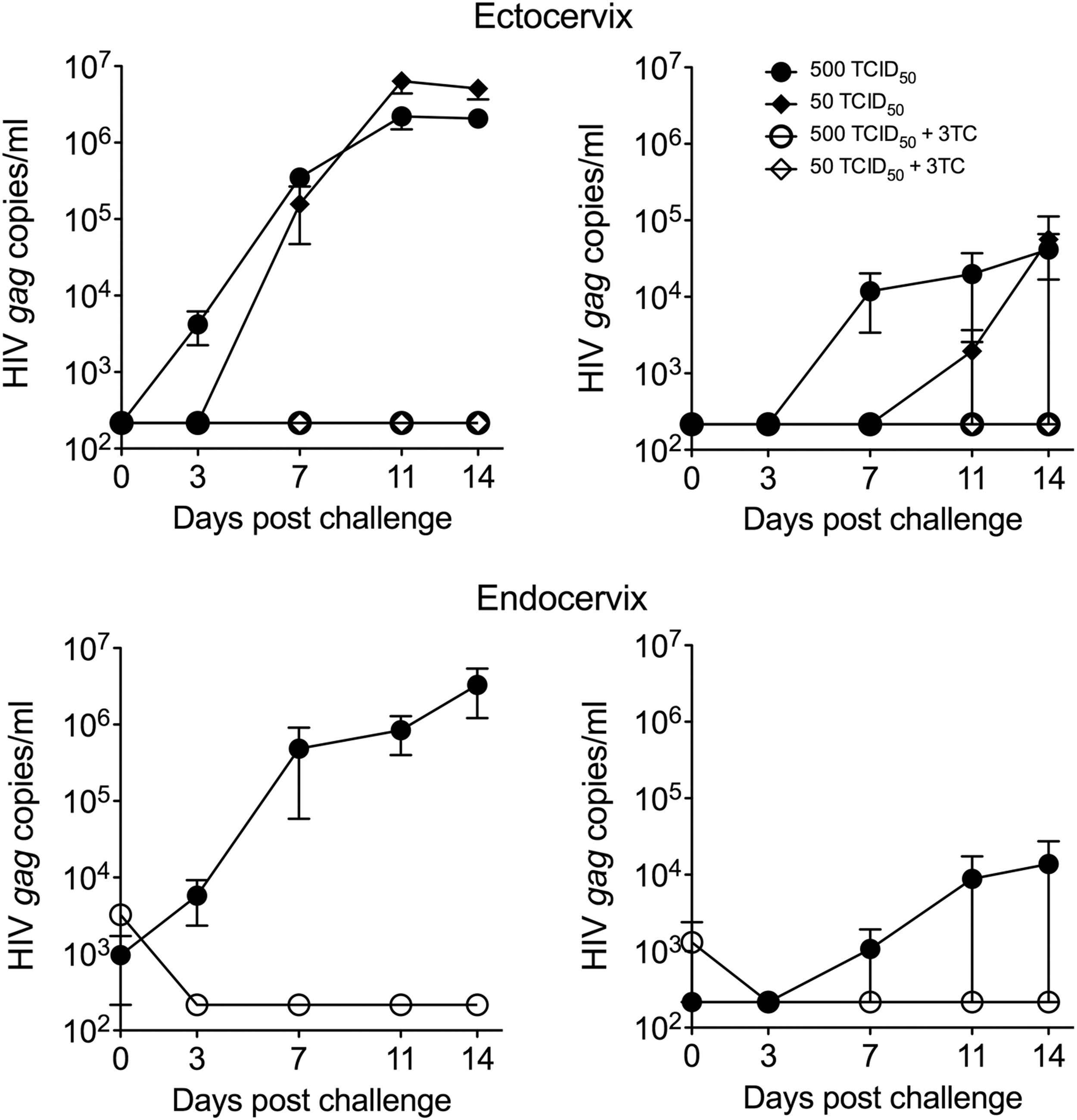
Viral growth kinetics following challenge with HIV-1BaL. Ecto- and endocervical explants were challenged with 500 or 50 TCID50 of HIV-1BaL or with HIV 500 TCID50 and 10 μM 3TC (two to three explants). Shown are examples (individual experiments) of viral growth kinetics (HIV gag copy numbers) over 14 days of culture (mean ± SEM of replicates) from four subjects (unpaired ectocervix and endocervix tissues) demonstrating higher and lower tissue infection levels. 3TC, Lamivudine; TCID50, 50% tissue culture infectious dose; SEM, standard error of the mean.
Ectocervical tissue infection level after challenge with 500 TCID50 HIV-1BaL was higher versus challenge with 50 TCID50 (Fig. 2) [p < .001 for SOFT (estimate 1.8197; 95% confidence interval (CI) 1.1658–2.4736)/CUM (estimate 1.7378; 95% CI 1.1447–2.3308)]. A lower infection level in endocervix as compared with ectocervix after 500 TCID50 HIV-1BaL challenge was noted, however, this is likely due to differences in the challenge protocols (challenge on the day of tissue recovery for ectocervix vs. after ON incubation at 4°C for endocervix). No decrease in tissue viability after ON incubation at 4°C was detected (Supplementary Fig. S2).
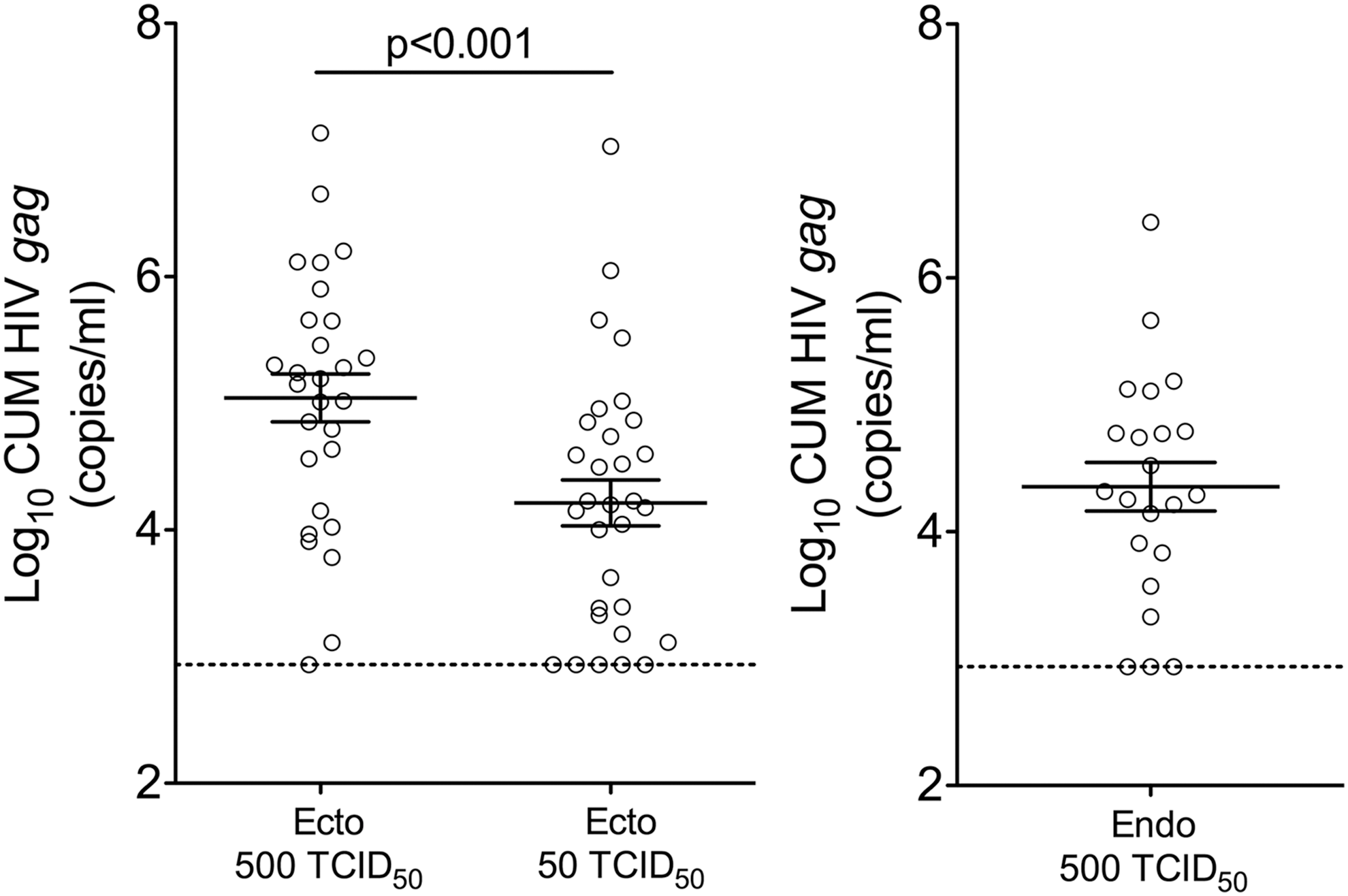
Tissue infection level following challenge with 500 and 50 TCID50 HIV-1BaL. Cumulative (CUM; days 3–14) analyses of HIV gag copies (mean ± SEM) are shown. Each symbol represents infection level in individual subject (mean of replicates). The dashed lines represent the LLOQ. LLOQ, lower limit of quantification.
Significantly more subjects had all ectocervical tissue replicates infected after 500 TCID50 (24/28) versus 50 TCID50 (17/31) challenge (p = .01). In the endocervical tissue challenge data set, 16/22 subjects had all tissue replicates infected. Of note, the same four subjects had uninfected ectocervical explants after 50 and 500 TCID50 challenge. Three of these subjects were in proliferative phase of the cycle and one had menses. Out of these four subjects, three had parakeratosis, one had metaplasia, one was HSV-2+. None of the subjects had cervical inflammation. Endocervical tissue challenge data were available from three of these four subjects. Two of them had tissues uninfected post 500 TCID50 challenge.
Cycle phase, subjects age, and hormonal concentrations
Hormonal concentrations in serum, ectocervix, and endocervix for all subjects were measured and are indicated in the Supplementary Table S4. Subjects were categorized into two groups based on the proliferative and secretory phases according to histological assessment of endometrium.
Serum P4 concentrations were higher in the secretory [8.08 ng/mL ±1.88 (mean ± SEM) vs. proliferative (0.98 ng/mL ±0.74)] phase of the cycle (p = .0002). The increased P4 levels detected in subjects 1041Z and 1056O in the proliferative phase are consistent with ovulation. 22 One subject (1064W) in the secretory phase had undetectable P4 levels. Serum E2 concentrations did not differ in the proliferative (158.5 pg/mL ±40.41) and secretory (175.5 pg/mL ±34.57) phases. No association between subject age and hormonal concentrations in serum were detected analyzing all subjects or subjects in the proliferative or secretory phase separately (p > .3).
E2 and P4 concentrations were undetectable in most of the tissue samples. P4 was detected only in two ectocervical tissue samples.
Cycle phase, hormonal concentrations, and tissue infection
Infection levels in the proliferative and secretory phases of the cycle were similar for both ecto- and endocervical tissues (p > .05 for SOFT/CUM) (Fig. 3). In accord with these results, no effect of the phases of the cycle on ecto- and endocervical tissue infectivity was noted (Supplementary Table S5).
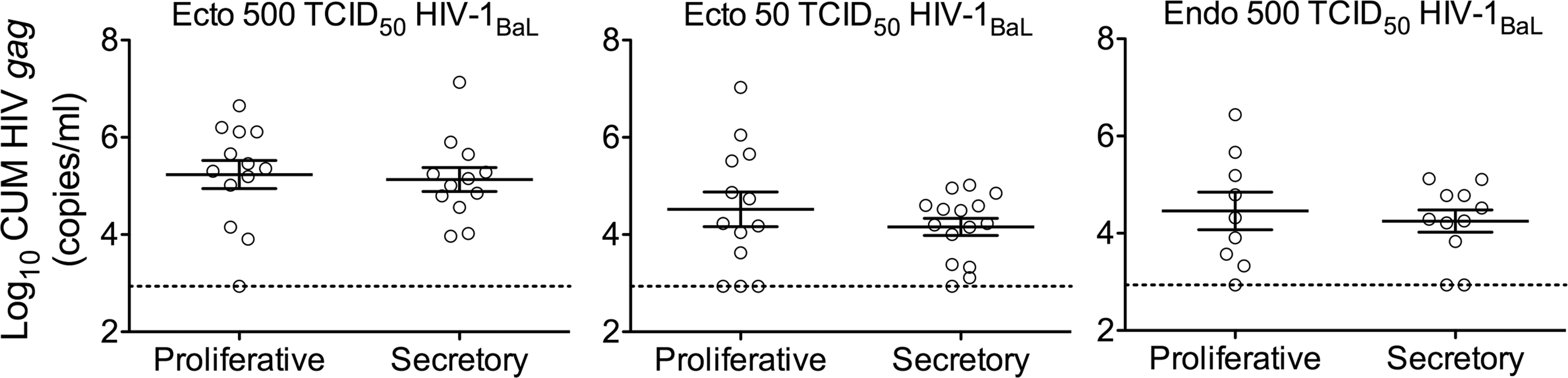
Tissue infection level in the proliferative and secretory phases of the cycle. Subjects were stratified based on the proliferative and secretory phase of the cycle. CUM analyses of HIV gag copies (mean ± SEM) are shown. Each symbol represents infection level in an individual subject (mean of replicates). The dashed lines represent the LLOQ.
In addition, we aimed to assess whether serum E2 and P4 concentrations and P4/E2 ratios are associated with tissue infection outcomes, irrespectively of the phase of the cycle.
Serum E2 concentrations were inversely associated with infection levels in ectocervical and endocervical tissues after 500 TCID50 HIV-1BaL (but not 50 TCID50) challenge (Table 1; Fig. 4). The associations remained significant adjusting for background parameters (Table 1). One exception was an adjustment of E2 effect for inflammation in CUM (but not SOFT) analysis following 500 TCID50 challenge of ectocervix, which resulted in p = .052, suggesting that cervical inflammation could have been a confounding factor in this analysis. P4 concentrations or P4/E2 ratios were not associated with tissue infection level (Table 1; Fig. 4). No associations between E2, P4, or P4/E2 ratios and tissue infectivity were noted either (Supplementary Table S6).

Serum E2, P4 concentrations and P4/E2 ratios and tissue infection level. CUM analyses (mean ± SEM) are plotted relative to serum E2, P4 concentrations and P4/E2 ratios. Shown are symbols representing infection level in individual subjects (mean of replicates). Regression lines are shown for descriptive purposes (i.e., trend). The dashed lines represent the LLOQ. Only significant p-values are indicated.
Relationship Between E2, P4 and P4/E2 Ratios and Tissue Infection Level
Significant p-values are indicated in bold.
Subjects included in the analysis. Hormonal measurements were available for 27/28, 30/31, and 22/22 subjects in Ecto 500, Ecto 50, and Endo 500 TCID50 data sets, respectively.
TCID50, 50% tissue culture infectious dose; CI, confidence interval.
Cycle phase, hormonal concentrations, and T cell frequencies and phenotype
Using PBMCs and an unchallenged portion of mucosa, we compared blood and tissue T cell frequencies in the proliferative and secretory phases of the cycle. The T cell gating strategy in PBMCs and tissues is demonstrated in Supplementary Fig. S3.
T cell subsets were defined within CD4+ T cells based on CCR7 and CD45RA expression [naive, CD45RA+CCR7+; central memory (Tcm), CD45RA−CCR7+; effector memory CD45RA−CCR7− (Tem); terminally differentiated effector memory (TEMRA), CD45RA+CCR7−]. Expressions of CCR5, CD103, CD54, LFA-1, and α4β7 were defined within CD4+ T cells. In addition, the frequency of CD4+CD45RA−α4β7+ memory cells was analyzed (total CD45RA−α4β7+ and CD45RA−α4β7high).
Expectedly, most of CD4+ T cells isolated from ecto- and endocervix had Tem phenotype and were CCR5+. Also, the tissue CD4+ T cells had higher frequencies of CD103+, CD54+-expressing cells, and lower frequencies of naive T cells (Fig. 5). The analysis revealed higher frequencies of CD4+α4β7+ T cells in ectocervix during the secretory versus proliferative phase (Fig. 5). No other significant differences in frequencies of T cells between the phases of the cycle were noted.
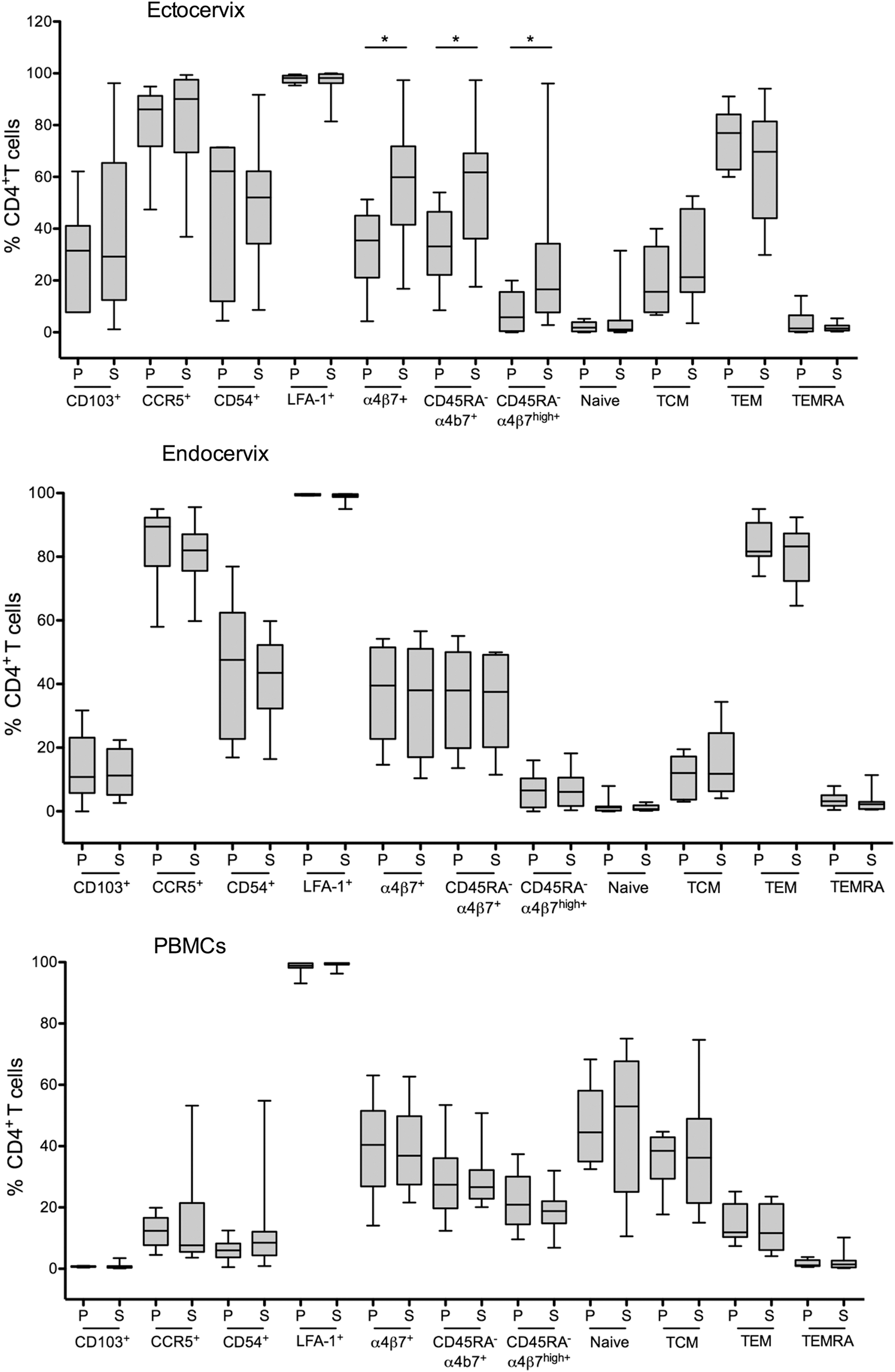
Frequencies of blood and tissue T cells in the proliferative and secretory phases of the cycle. Subjects were stratified based on the proliferative and secretory phase of the cycle. Frequencies of CD4+ T cells subsets expressing indicated markers are shown for ectocervix [n = 6 proliferative (P), n = 14 secretory (S)], endocervix (n = 8 P, n = 10 S) and PBMCs (n = 9 P, n = 12 S). *Indicates p < .05. PBMC, peripheral blood mononuclear cells.
Additionally, effects of serum E2 and P4 concentrations and P4/E2 ratios on T cell phenotype/frequencies were explored (Fig. 6). E2 concentrations were inversely associated with CD4+α4β7+ T cells in the endocervix. In ectocervix, P4/E2 ratios were directly associated with CD4+α4β7+ T cells. Subjects with high P4 and/or P4/E2 ratios also had decreased frequencies of CD4+CD103+ and CD4+LFA-1+ T cells in blood and endocervix, respectively. It needs to be noted that frequencies of CD4+CD103+ T cells in blood were extremely low (0.7% ± 0.14; mean ± SEM; Fig. 6). No associations between hormonal concentrations/ratios and either functional T cell subset frequencies or frequencies of CD54 or CCR5-expressing cells were detected.
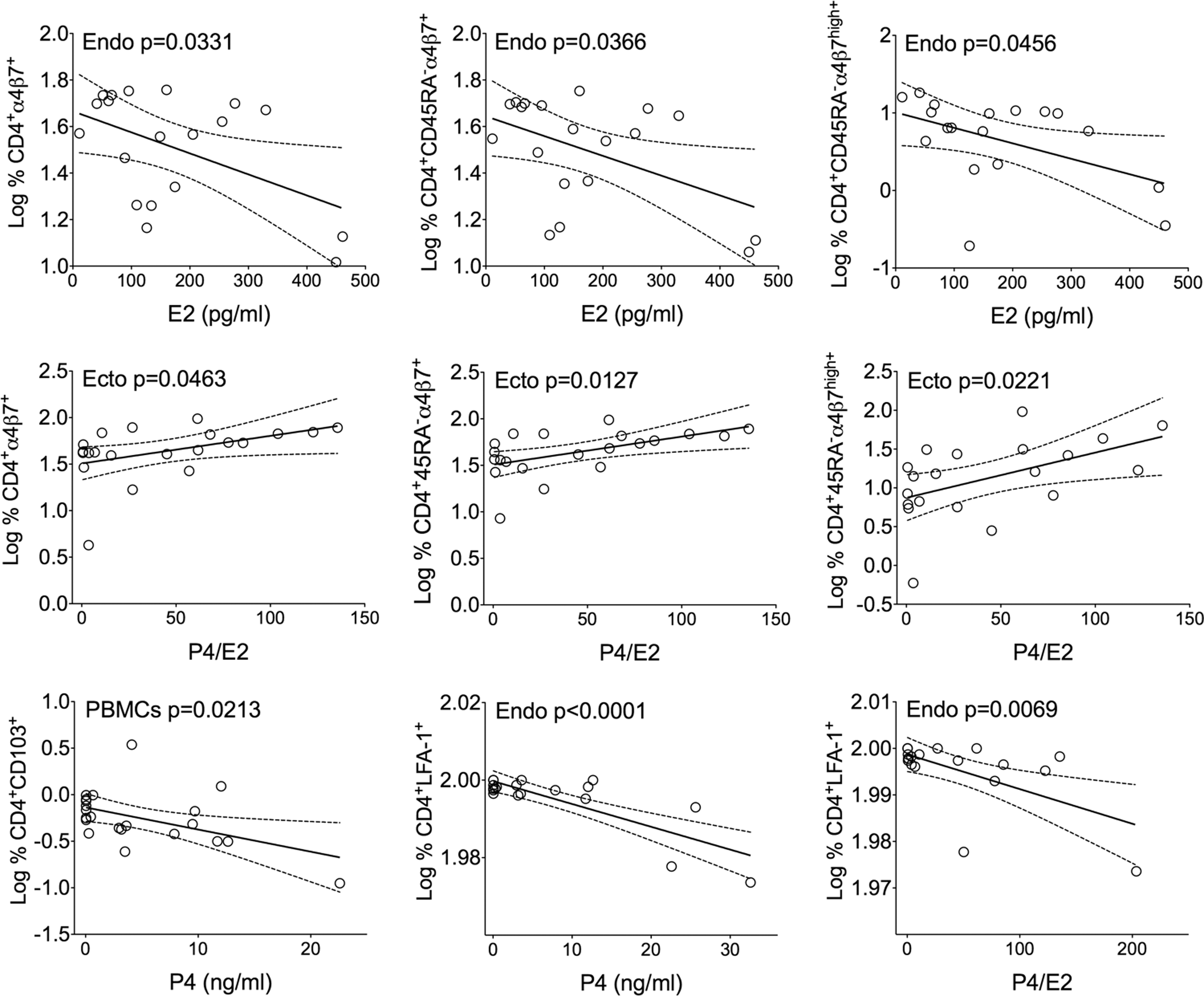
Significant associations between E2, P4 concentrations, P4/E2 ratios, and Log % CD4+ T cells. Frequencies of PBMCs and tissue T cells expressing CD103, α4β7, or LFA-1 are plotted relative to E2, P4 concentrations, and P4/E2 ratios. Each symbol represents an individual subject. Linear regression analyses and 95% CIs are shown. CI, confidence intervals.
Cycle phase, hormonal concentrations, and innate mediators profile in response to HIV-1BaL in ectocervix
Soluble mediators' concentrations after exposure of ectocervical mucosa to 500 TCID50 HIV-1BaL (vs. no viral exposure control) were analyzed by Luminex. Two (IL2 and IL5) out of 25 analyzed mediators were not detected. Subjects in the secretory phase of the cycle had stronger induction of proinflammatory IL6, IL7, and CXCL8 and subjects in proliferative phase had stronger anti-inflammatory IL1RA response (Fig. 7). E2 and P4 concentrations predicted different immune mediator profiles in response to HIV-1BaL exposure (Fig. 8). High E2 concentrations were associated with low IL2R and IL4 post HIV exposure. High P4/E2 ratios were associated with high proinflammatory response (IL1β, IL17, and TNFα).

HIV-1BaL-induced changes in soluble mediators profile in the proliferative and secretory phases of the cycle in ectocervical mucosa. Tissue culture supernatants were collected 18 h postexposure to HIV-1BaL (vs. medium control) and analyzed by 25-plex Luminex. Shown are HIV-1BaL-induced responses (HIV-1BaL induced/baseline fold change; mean ± SEM) for subjects in the proliferative (P; n = 11) and secretory (S; n = 13) phases of the cycle.

Significant associations between E2 and P4/E2 ratios and HIV-induced responses in ectocervical mucosa. Virus-induced responses in tissue cultures (fold changes in the concentrations of individual mediator relative to untreated medium control) are plotted relative to E2 and P4/E2 ratios. Outliers were identified in IL2R, IL1β, IL17, and TNFα data sets, but they did not affect the significant associations revealed by linear regression analyses. Linear regression analyses and 95% CIs are shown. IL, interleukin; TNFα, tumor necrosis factor α.
Discussion
The study revealed similar ectocervical and endocervical tissue infection level/infectivity with HIV-1BaL in the proliferative and secretory phases of the cycle. These results are consistent with a recent report by Thurman et al. 11 utilizing ectocervical and vaginal biopsies, which demonstrated a lack of association between the phases of the cycle and ex vivo tissue infection after challenge with HIV-1BaL. An earlier report by Saba et al. demonstrated productive HIV-1BaL infection ex vivo (combined ecto-/endocervix hysterectomy specimens) only from women in the secretory phase. 10
A number of methodological differences could have contributed to these discrepant results. Our study involved (i) separate analysis of ectocervical and endocervical tissues, and (ii) overall much lower (“high” 500 and “low” 50 TCID50) tissue challenge doses/explant compared with 2.1 × 106 TCID50 HIV-1BaL/explant challenge used in the Saba et al. report. 10
The tissue collection in narrow periods during the menstrual cycle (e.g., close to ovulation at the peak of E2 during proliferative phase and at the peak of P4 concentrations during secretory phase) could have potentially revealed differences in tissue infection/infectivity. Our results demonstrating an inverse association between serum E2 concentrations and endocervical and ectocervical infection support this possibility and suggest E2-mediated protection against HIV acquisition. The analysis was adjusted for multiple covariates (race, age, endometrial histology, parakeratosis, metaplasia, histopathological signs of cervical inflammation, and HSV-2 status). Of note, inflammation could have been a confounding factor in the analysis of association between E2 and HIV-1BaL infection level (CUM analysis) after 500 TCID50 HIV-1BaL challenge of ectocervix.
The inverse association between serum E2 concentrations and tissue infection level is concurrent with in vivo data demonstrating protection afforded by E2-containing implants and estriol vaginal cream treatment against vaginal simian immunodeficiency virus (SIV) acquisition in ovariectomized rhesus macaques, 23,24 and with ex vivo ectocervical tissue infection inhibition following topical administration of E2-containing vaginal cream in postmenopausal women. 25
Similar to the report by Thurman et al., 11 no association between E2 concentrations and ectocervical tissue infection following low 50 TCID50 challenge was detected. We theorize that the lack of association between serum E2 concentrations and infection level after low-dose challenge, leading to less robust infection, is possibly due to E2-mediated regulation of infection once it is established. Supporting this argument, E2 concentrations were not associated with tissue infectivity in our study. P4 concentrations or P4/E2 ratios did not predict tissue infection level or infectivity.
More pronounced proinflammatory (high IL6, IL7, and CXCL8 and low IL1RA) response to HIV in ectocervical tissues from subjects in secretory phase was detected. IL6 is known to induce HIV expression in vitro. 26 In cervicovaginal lavage fluids from HIV-infected women IL6 was linked with increased HIV shedding, suggesting that this proinflammatory factor induces HIV gene expression in the genital tract. 27 In ectocervical explants enhanced secretion of IL6 is associated with increased HIV transcription. 28
IL-7, a cytokine involved in thymopoiesis as well as in the homeostasis and survival of mature T cells, 29 –31 was shown to enhance HIV replication in cervicovaginal explants through inhibition of apoptosis and promoting of CD4 T cell proliferation. 32
The role of CXCL8 during HIV infection is not fully understood. CXCL8 is a known chemotactic factor for neutrophils, basophils, T cells, and monocytes 33 –37 and CXCL8 signaling was identified among top pathways associated with the luteal phase of the cycle in a vaginal fluid proteome study. 38 Contrasting results on the effects of CXCL8 on HIV replication in T lymphocytes were reported. 39 –41 In ectocervical explants, CXCL8 was shown to decrease HIV transcription. 41
IL-1RA was previously reported to block IL-1-induced HIV expression in U1 cells. 42,43
Overall, detected immune mediator responses induced by HIV are congruent with inflammatory and immune-activated state described in the secretory phase vaginal fluid proteome, 38 which can promote HIV acquisition. However, the data also indicate potentially opposing effects of HIV-1BaL-induced cytokines during mucosal infection, which could have contributed to the lack of differences in ex vivo HIV-1BaL tissue challenge outcomes between proliferative and secretory phases of the cycle in our study.
Frequencies of CD4+CCR5+ T cells in ectocervical and endocervical mucosa in our study did not vary between the phases of the cycle and are concurrent with data in vaginal 25,44 and ectocervical 45 mucosa using immunohistochemistry and immunofluorescence. In contrast, a recent report utilizing endocervical cytobrush specimens and using flow cytometry demonstrated increased frequencies of CD4+CCR5+ T cells in luteal versus follicular phase. 46 No differences in the frequencies of CD103, CD54, LFA-1-expressing CD4+ T cells, and functional T cell subsets were observed between the phases of the cycle in our study.
Analysis of mucosal CD4+ T cells expressing α4β7, a gut-homing integrin 47 –50 binding with high affinity to HIV gp120, 51 and the expression on peripheral blood T cells that predicts HIV acquisition, 52 revealed higher frequencies of CD4+α4β7+ T cells in ectocervix in the secretory phase. However, higher frequencies of CD4+α4β7+ T cells in ectocervix in the secretory phase were not predictive of infection ex vivo, which could have been due to contribution of HIV-1BaL-induced immune responses with opposing effects on HIV infection, as discussed above.
E2 concentrations were inversely associated with frequencies of CD4+α4β7+ T cells in the endocervix and P4/E2 ratios were directly associated with frequencies of CD4+α4β7+ T cells in the ectocervix. Together with the inverse association of E2 levels and tissue infection, these data suggest a new potential mechanism underlying E2-mediated protection against HIV infection in the cervix, adding to several other proposed mechanisms. 53 –55
These data also suggest opposing roles for E2 and P4 in the regulation of α4β7+ T cell mucosal trafficking and, potentially, during HIV acquisition. Of note, injection of a high-dose (30 mg) exogenous progestin depot medroxyprogesterone acetate (DMPA) in rhesus macaques was previously shown to regulate T cell trafficking and to increase frequencies of CD4+α4β7+ T cells in endocervical mucosa. 56 DMPA is known to increase susceptibility to vaginal SIV/SHIV acquisition, enhance genetic complexity of replicating virus, and peak viremia. 57,58 DMPA is also known to decrease serum E2. 59
Other significant findings in our study included inverse associations between P4 and P4/E2 ratios and LFA-1 expression in endocervix. LFA-1 expression on target cells increases susceptibility to HIV-1 infection through cell-free virions. 60,61 P4-associated changes in frequencies of CD4+α4β7+ and CD4+LFA-1+ T cells suggest that P4 may have contrasting effects during HIV acquisition.
High E2 concentrations were predictive of low IL2R and IL4 levels in tissue culture post HIV exposure. IL4 is known to have a dual effect on HIV infection by downregulating CCR5 expression on T cells (but not on macrophages) and activating HIV expression through transcriptional mechanism. 62 In contrast, subjects with high P4/E2 ratios had high levels of proinflammatory cytokines (IL1β, IL17, and TNFα).
Our study had several limitations. Given that our study cohort were women undergoing hysterectomies, the participants were primarily midlife women and, therefore, the results may not be generalizable to younger women who are at greater risk of HIV transmission. However, midlife women are still at significant risk of HIV acquisition. 63 –65 Understanding effects of the menstrual cycle during vaginal HIV acquisition in older premenopausal women may help to target prevention strategies in this important population.
With increasing age and during transition to menopause, E2 and P4 levels decrease among other menstrual cycle-related changes, 66,67 representing a caveat in our study. However, among our study participants (ranging in age from 35 to 53), we did not detect an association between subject age and hormonal concentrations in serum.
Limited amount of available tissues precluded analysis how E2 impacts tissue susceptibility to HIV-1 per se (i.e., median TCID50). Also, using different HIV-1BaL challenge protocols for ectocervix and endocervix precluded comparisons of challenge outcomes between different mucosae. To address some limitations inherent to the usage of hysterectomy specimens, 68 our study included women who did not use hormonal contraception and presurgical hormonal therapy for gynecological conditions.
Another limitation in the study was that viral stock was not purified, and, although we used very small volume (2.1 μL/explant; 1:100 dilution in the explant culture medium), soluble factors present in the viral stock could have influenced analysis of tissue responses to virus exposure.
In summary, the results suggest a link between serum E2 concentrations and cervical tissue infection, provide insights into possible mechanisms of the E2-mediated anti-HIV effect in the mucosa and a potential use of E2 treatment as an HIV prevention strategy.
Footnotes
Acknowledgments
The authors thank Nina Derby and Lyle McKinnon for critical reading of the article. This project was funded by the National Institutes of Health grant RO1AI110370.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.