
Abstract
Female sex hormones, the octamer-binding transcription factor 4 (OCT4), and human endogenous retroviruses (HERVs) are all involved in the development of breast cancer. However, whether there are cross talks between these factors to promote breast cancer is still unknown. Using the T47D human breast cancer cell line, we have found that estradiol and progesterone synergistically activate HERV-K through nuclear receptors. The progesterone receptor (isoform B) binds a progesterone-response element (PRE) in a long terminal repeat (LTR5HS) of HERV-K. There is another transcription factor-binding element in the LTR, the octamer motif, which is required for the hormones to activate gene transcription downstream of the LTR. Gel shift assays and co-immunoprecipitation indicate that the progesterone receptor (PR) and the OCT4 transcription factor interact on the protein level. Methylation of the PRE enhances binding of the PR. These findings help to elucidate the previously unknown cross talks among the sex hormones, OCT4, and HERVs in contributing to breast cancer proliferation and tumorigenesis, which may be useful in guiding further development of cancer therapies.
Introduction
Breast cancer is mediated, in part, by dysregulated cellular responses to female sex hormones. The role of estrogens in breast cancer development is well established, but the role of progesterone remains unclear. 1 Normally, the actions of progesterone depend on prior existence of estrogen to stimulate the expression of progesterone receptors (PRs). There are several isoforms of PRs. The 120 kDa PR-B binds the progesterone-response element (PRE) in the promoter region of target genes to activate transcription, while the N-terminally truncated 94-kDa PR-A antagonizes PR-B. 2 The 60-kDa PR-C is further truncated at the N-terminus and its expression and physiological function in the breast gland are still largely unstudied. 3
The main mechanism by which progesterone promotes cell proliferation is through mitogen-activated protein kinase (MAPK) pathways, which enable the PR to regulate growth-related genes such as CCND1, which do not contain a PRE (see Lange and Yee 4 for review). In addition, MAPKs phosphorylate PR-B itself to enhance its function, forming a positive feedback loop. 5 Two parallel pathways are known to transduce signals between the ligand-bound PR and the cell proliferation machinery: a rapid, transient pathway that involves interactions between the estrogen receptor (ER), PR, and c-Src to phosphorylate Ras and subsequently ERK1/2, and a sustained pathway through Wnt-1, the epidermal growth factor receptor, c-Src, Ras, and ERK1/2, with the sustained pathway being dominant. 6,7
Recently, the Env protein of human endogenous retrovirus type K (HERV-K) was found to induce cell proliferation, migration, invasion, tumor formation, and epithelial to mesenchymal transition through the ERK1/2 pathway. 8,9 Since female sex hormones are known to activate expression of HERV-K, 10 –12 it is possible that Env serves as a mediator of the progrowth effects of progesterone. It most likely works upstream of Wnt-1, since Wnt-1 knockdown completely blocks ERK activation by progesterone, 7 and the mouse WNT-1 gene is known to be activated in mammary tumors by insertion of the mouse mammary tumor virus (the mouse orthologue of HERV-K). 13 HERV-K Env was also found to upregulate the expression of H-Ras and K-Ras. 8
With a canonical PRE, LTR5HS of HERV-K is a potential target of PR binding. LTR5HS elements at the 5′ end of proviruses drive transcription of viral genes, while solitary LTR5HS elements can drive transcription of downstream host sequences. 14,15 In embryonic stem cells, LTR5HS is activated by DNA demethylation and the octamer-binding transcription factor 4 (OCT4). 14 As a gatekeeper of stem cell-like state and a preventer of differentiation, OCT4 is overexpressed in breast cancer cells and is correlated with cell proliferation, metastatic ability, and poor prognosis. 16,17 The molecular mechanisms by which OCT4 contributes to tumorigenesis are still unclear.
The goal of this study was to investigate the mechanisms of hormonal activation of HERV-K elements, including the involvement of hormone receptors, viral regulatory sequences, DNA methylation, and the OCT4 transcription factor. Our results point to the coordinated actions of PR and OCT4 on LTR5HS to activate expression of downstream viral or host sequences in breast cancer cells.
Materials and Methods
Cell lines and culture
T47D breast cancer cell line was maintained in RPMI 1640 with 10% fetal bovine serum. Primary mammary epithelial cells were obtained from ATCC and were maintained in Mammary Epithelial Cell Basal Medium (ATCC) supplemented with the Mammary Epithelial Cell growth kit (ATCC). All cells were cultured at 37°C and 5% CO2.
Extraction of RNA, reverse transcription, and quantitative polymerase chain reaction
Total cell RNA was extracted from cultured cells using the RNeasy Plus Mini Kit (Qiagen). Complementary DNA was prepared using the Quantitect Reverse Transcription Kit (Qiagen). Real-time polymerase chain reaction (PCR) was performed using the LightCycler 96 (Roche) and SsoAdvanced Universal SYBR Green Supermix (Bio-Rad).
A pair of slightly degenerate primers targeting the pol gene of class II HERVs (HERV-K), shown to amplify HML-2 and HML-3 subtypes of HERV-K,
18
was used. Another pair of primers targets the env gene of HERV-K10 in the HML-2 subgroup.
19
Primers for OCT4 (POU5F1) were purchased from
All primer sequences are shown in Supplementary Table S1.
HERV genes and the YWHAZ gene were amplified at 95°C 15″, 57°C 15″, 72°C 45″, and 78°C 5″ for 40 cycles. Fluorescence was read at 78°C. The OCT4 gene was amplified using a touchdown PCR: 95°C 15″, touchdown (with annealing temperatures decreasing from 61°C to 58°C in 30 cycles) 45″, and 78°C 5″ for 50 cycles.
Plasmid and constructs
Entire long terminal repeats (LTRs) were cloned into luciferase reporter plasmids. A solo LTR5HS at 12q13.13 between exons 5 and 6 of the SLC4A8 gene that drives expression of antisense RNA 13 was amplified from genomic DNA of T47D cells using CloneAmp HiFi PCR Premix (Clontech). The primers were 5′-ACTGAAGAGGTAGGGGCACT-3′ and 5′-GCTAGGCATGGGGTTATGAA-3′. Similarly, a reversely oriented solo LTR13A in the first intron of the ITPR3 gene was amplified using primers 5′-GTGGGGCAGGTGACTATCAA-3′ and 5′-GTTGGCACAGGGTGGATTCT-3′. The amplicons were cloned into pMetLuc2-Reporter (Clontech) using the In-Fusion HD Cloning Plus Kit (Clontech).
Mutant and wild-type LTR5HS from HERV-KCON cloned into pGL3 were kindly donated by Dr. Joanna Wysocka. In the mutant, the octamer motif was replaced with a NotI site. 14
Transfections and reporter assays
For luciferase reporter assays, T47D cells were treated with estradiol (10 nM; Sigma-Aldrich) for 24 h followed by a combination of estradiol (10 nM) and progesterone (1 μM; Sigma-Aldrich) for 15 h. Immediately after progesterone was added, cells were co-transfected with the pMetLuc2-Reporter construct and an equal amount of a control plasmid that constitutively expressed a secreted alkaline phosphatase (pSEAP2-Control). The Xfect transfection reagent (Clontech) was used. Activity of secreted Metridia luciferase and alkaline phosphatase was quantified using the Ready-To-Glow™ Dual Secreted Reporter Vector Kit (Clontech). Activity of intracellular firefly luciferase was quantified using the Steady-Luc Firefly HTS Assay Kit (Biotium).
siRNA knockdown
Small interfering RNA against the human OCT4 (POU5F1) gene was purchased from Origene, along with the transfection reagent, siTran 1.0. Cells were transfected with siRNA or random control RNA twice in consecutive days. Estradiol was added to 10 nM immediately after the second transfection (day 2), and progesterone was added on day 3 at 1 μM, 15 h before harvest. Total cell RNA was harvested 48 h after the second transfection on day 4. Knockdown efficiency was evaluated with quantitative reverse-transcription PCR (RT-PCR).
Activation of HERV-K in primary mammary epithelial cells
For activating HERVs in primary mammary epithelial cells, cells were treated with decitabine (10 μM; Sigma-Aldrich) or solvent for 2 days, with medium change every day and fresh decitabine added. When cells were 60% confluent, they were transfected with pcDNA_OCT4 (a gift from Derrick Rossi) using siTran 1.0 (Origene) and CombiMag (OZBiosciences). For hormone treatments, estradiol was added to 10 nM after transfection. After 24 h of estradiol treatment, cells were treated with a combination of estradiol (10 nM) and progesterone (100 nM) for 14–15 h.
Electrophoresis mobility shift assay
We performed electrophoresis mobility shift assays (EMSA) using the Odyssey Infrared EMSA Buffer Kit (LI-COR). Crude DNA-free nuclear extract from the T47D breast cancer cell line was used as a source of PR and OCT4. The cells were treated with estradiol (10 nM) for 39 h. Nuclear extract was prepared using the EpiQuik Nuclear Extraction Kit II (Epigentek).
A 27-mer oligonucleotide containing the PRE (TTAAGGGC
Binding reactions were assembled for a total reaction volume of 20 μL, containing 10 mM Tris, 100 mM KCl, 3.5 mM dithiothreitol, 1 μL of LightShift™ Poly (dI·dC), 500 nM progesterone, 10% glycerol, 50 fmole of each labeled probe, and 4 μg of nuclear extract (unless otherwise indicated). The reaction was incubated at room temperature for 20 min. For competition experiments, the unlabeled oligonucleotide was added before labeled probes and incubated at room temperature for 7–8 min.
For supershifting, 1 μL of undiluted mouse monoclonal anti-PR (MA1412; Invitrogen) or rabbit monoclonal anti-OCT4 (M00174; Boster) was added to the reaction and incubated for 10 min at room temperature followed by 1 h on ice. Normal mouse and rabbit sera served as controls. Concentration of dithiothreitol was reduced to 2 mM for anti-PR and mouse serum, and 1.5 mM for anti-OCT4 and rabbit serum.
Polyacrylamide gel electrophoresis was carried out using nondenaturing Mini-PROTEAN TBE gels (Bio-Rad). Oligonucleotide bands were visualized using an Odyssey CLx scanner.
Co-immunoprecipitation
Pierce Co-Immunoprecipitation Kit (Thermo Scientific) was used. Ten micrograms of rabbit anti-OCT4 (M00174; Boster) or rabbit anti-PR monoclonal antibodies (M00541-1; Boster) was linked onto AminoLink Plus Coupling Resin. Rabbit serum was used as negative control. Linked antibodies were allowed to react with nuclear extract from T47D cells stimulated sequentially with estradiol and progesterone. After washing resin with phosphate-buffered saline, proteins were eluted, separated with an Any kD™ Mini-PROTEAN® TGX™ precast protein gel, and transferred onto a nitrocellulose membrane using the Mini Trans-Blot® Cell (Bio-Rad). PR proteins were detected using rabbit anti-PR and the Enhanced Chemiluminescent Reagent Kit (Goat anti-rabbit IgG; Boster).
Statistical analysis
All quantitative data were analyzed using the Mann–Whitney U test. Statistical significance is defined at p < .05.
Results
Female sex hormones enhance HERV-K expression in T47D cells through nuclear receptors
According to a protocol established by Ono et al., 10 we treated T47D cells with estradiol (10 nM) for 24 h followed by a combination of estradiol (10 nM) and progesterone (1 μM) treatment for 13–15 h. Using quantitative reverse-transcription PCR, we analyzed the expression of various HERV elements. Consistent with previous findings, we observed hormonal activation of HERV-K env and pol genes in the cell line (Fig. 1A, B) and that the effect of progesterone is dose related (Fig. 1C). Fulvestrant, an estrogen antagonist, and mifepristone, a progesterone antagonist, blocked the effect of the hormones. Because these drugs bind nuclear receptors, it is evident that estradiol and progesterone activate HERV-K through nuclear receptors.

Effect of estradiol and progesterone on HERV-K and adjacent genes in T47D cells.
Testosterone or cortisol had no effect on the expression of HERV-K env (data not shown).
Expression of class I HERV elements, syncytin-1 and syncytin-2, did not respond to female sex hormones (not shown).
Female sex hormones regulate host gene expression through solo LTRs
Gogvadze et al. 15 reported that two solo LTRs, both belonging to the LTR5HS subgroup, drove expression of downstream host DNA sequences. Both reside in introns on the noncoding strand, initiating transcription of antisense RNA against the host gene. One of them is located between exons 5 and 6 of a gene named SLC4A8, and the other is located between exons 23 and 24 of a gene named IFT172. T47D cells provide a model to investigate the effect of female sex hormones on gene expression driven by these solo LTRs. Using primers designed by Gogvadze et al., we indeed found a stimulating effect of hormones on LTR-driven expression of downstream host sequences, that is, antisense RNA against exons of SLC4A8 and IFT172 (Fig. 1D). Mifepristone, a PR antagonist, blocked the effect of female sex hormones.
LTR5HS drives expression of reporter genes and responds to sex hormones in T47D cells
To investigate the molecular mechanisms of the action of steroid hormones, we cloned LTR5HS and LTR13A, both found in HERV-Ks, upstream of a luciferase reporter gene. The LTR5HS was amplified from the SLC4A8 intron at 12q13.13, while LTR13A was amplified from the first intron of the ITPR3 gene at 6p21.31. Upon transfection into T47D cells, we found that estradiol and progesterone upregulated the promoter/enhanced activity of LTR5HS, but not that of LTR13A (Fig. 2A). In the absence of progesterone, the effect of the LTRs on the expression of the reporter gene was comparable to that of the vector alone.
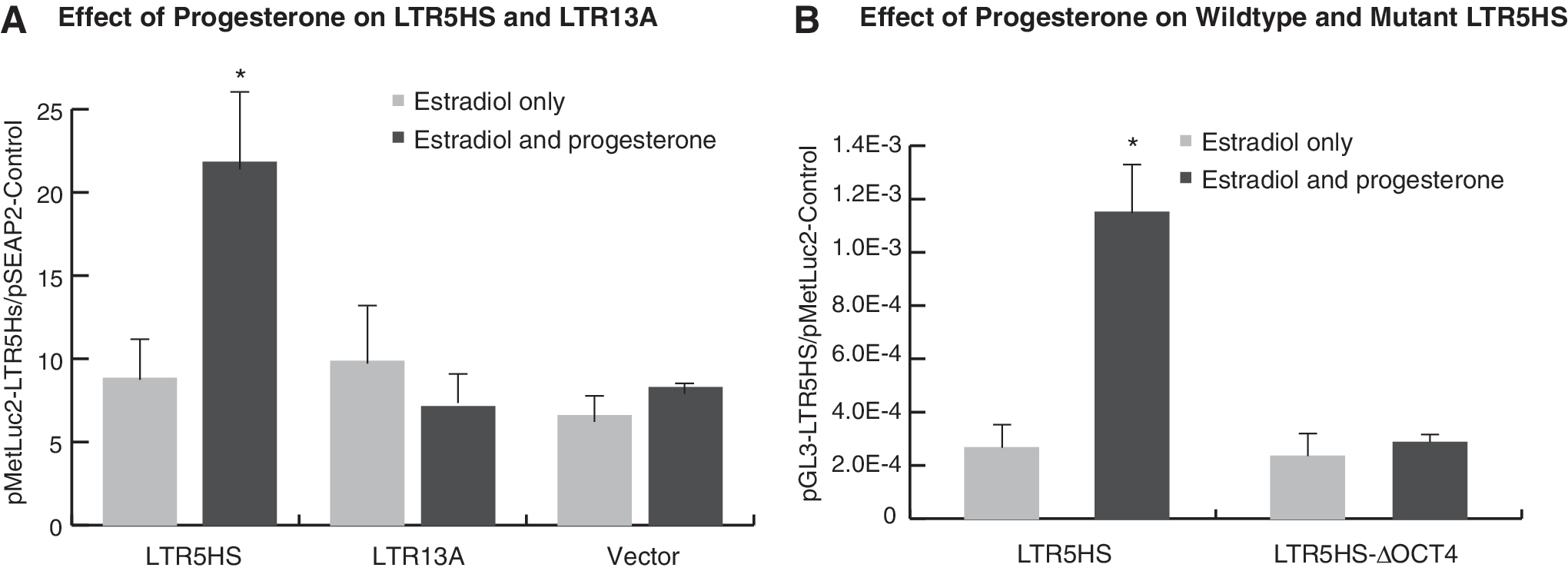
Effect of estradiol and progesterone on HERV-K LTRs in transiently transfected T47D cells.
Using LTR5HS from HERV-KCON, 23 Grow et al. 14 found that the promoter/enhancer activity of LTR5HS was dependent on an octamer motif, which is located about 500 bp downstream of the conserved androgen-response element/PRE/glucocorticoid response element. 24 With the plasmids constructed by Grow et al., we demonstrated that responsiveness to sex hormones is completely abolished if the octamer motif is deleted (Fig. 2B).
Knocking down OCT4 reduces expression of HERV-K env
To further investigate the role of OCT4 in activation of HERV-K elements, we used specific siRNA to knockdown OCT4 expression in T47D cells. Transfection with OCT4-specific siRNA significantly reduced the expression of HERV-K10 env compared to control samples transfected with random duplex RNA, although the latter also showed some suppressive effect, presumably due to nonspecific hybridization (Fig. 3A). 25 (Quantitative RT-PCR demonstrated that the knockdown efficiency of the siRNA was 70.7%–78.5% compared to random duplex RNA.) Similarly, expression of antisense RNA driven by the solo LTR in the SLC4A8 gene was inhibited by OCT4 knockdown (not shown).

OCT4 mediates the effect of progesterone in T47D cells.
DNA demethylation and overexpression of OCT4 activate HERV-K expression in primary mammary epithelial cells
It has been long recognized that sex hormones do not stimulate expression of HERV genes in normal mammary epithelium. 19 With quantitative PCR, we observed a slight stimulating effect of sex hormones on the expression of OCT4 in T47D cells (Fig. 3B). Therefore, we hypothesized that transfection of primary epithelial cells with an OCT4-expressing plasmid may achieve an effect similar to hormonal treatment of T47D cells. In addition to overexpression of OCT4, we also treated the primary cells with decitabine, an inhibitor of DNA methyltransferase, since DNA demethylation is known to activate HERV-K in HEK293 cells. 14
As expected, we found both OCT4 and decitabine enhanced expression of the HERV-K10 env gene in primary mammary epithelial cells. Moreover, we observed a synergistic effect of the two treatments. Hormonal treatment did not make a significant difference on HERV-K10 expression in primary cells (Fig. 3C).
Methylated PRE binds PR with higher affinity
The PRE in LTR5HS contains a CpG pair (nucleotide 193–194). To study the effect of cytosine methylation, double-stranded 27-mer oligonucleotide probes (nucleotide 186–212) were synthesized for EMSA. In one version, the cytosine residue of the CpG pair on both strands was methylated and the probe was labeled with IRDye800 at the 5′ ends. Another version of the probe, labeled with IRDye700, was not methylated. Because the fluorophores emit light at different wavelengths, binding of PR onto both probes can be carried out in the same tubes to compare the binding affinity of each probe.
As can be seen in Figure 4A–C, more of the methylated probe was shifted, while more of the unmethylated probe remained unbound, indicating that the methylated probe had a higher protein-binding affinity. Protein binding of both probes could be competitively inhibited with an unlabeled, unmethylated version of the PRE probe, with the unmethylated probe being more easily competed off.
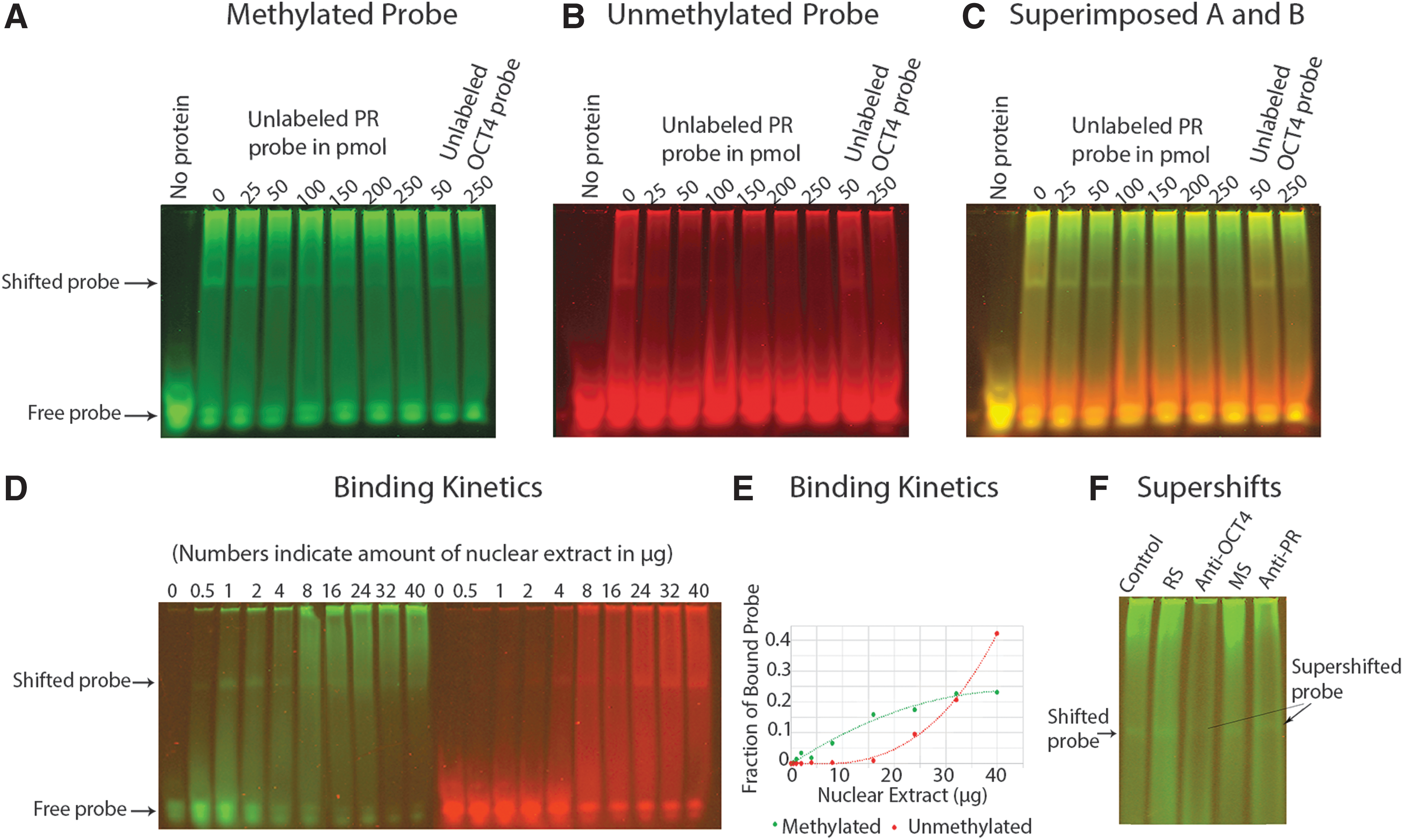
Binding of methylated and unmethylated PRE to PRs in nuclear extract of T47D cells.
Protein-binding kinetics demonstrated preferential binding of nuclear extract to methylated DNA (Fig. 4D, E). In this experiment, 50 fmoles of each probe was allowed to react with increasing concentrations of nuclear extract. As shown in Figure 4D, the methylated probe bound low concentrations of proteins more tightly, while the unmethylated probe only bound proteins when the concentrations of nuclear extract were high. A plot of bound fractions of the fluorophores against concentrations of nuclear extract revealed distinct binding kinetics of the two probes (Fig. 4E). The methylated probe approached a second-order polynomial kinetics and was saturated within the range of the experiment, while the unmethylated probe demonstrated a third-order polynomial kinetics and was not saturated within the range of the experiment.
To confirm that the shifted band contained a PR, a mouse monoclonal antibody that binds both PR-A and PR-B was used in supershifting experiments. The antibody consistently supershifted the DNA-protein complex, as seen in Figure 4F. Significantly, a rabbit monoclonal antibody against OCT4 also supershifted the complex, indicating that OCT4 was also in the complex.
Co-immunoprecipitation of PR and OCT4
To further investigate potential interactions between PR and the OCT4 transcription factor, rabbit anti-OCT4 was linked to resin and incubated with nuclear extract from T47D cells that had been treated sequentially with estradiol and progesterone. After washing with phosphate-buffered saline, bound proteins were eluted and subjected to Western blot analysis. As seen in Figure 5A, the rabbit monoclonal anti-OCT4 was able to pull down PR-B (119 kDa). The size of the PR-B band was consistent with the result of immunoblotting using T47D nuclear extract and the same primary antibody (Fig. 5B). The higher molecular weight protein (∼220 kDa) in Figure 5A, which was present in both the precipitates and the flow-through, but not in the serum control, was presumably a nonspecific signal from the nuclear extract.

Discussion
While there has been some research on the activation of HERVs in early embryos, 14 our project focuses on hormonal regulation of these elements. Our major findings are as follows: Class II HERVs are regulated by female sex hormones in T47D cells through nuclear receptors; sex hormones regulate cellular sequences through a class of solo LTRs; the effect of progesterone on HERV-K expression is at least partly mediated by OCT4; and DNA methylation has an overall effect of suppressing HERV-K expression, but enhances the binding of the PR on DNA.
Of particular interest is the requirement of two transcription factor-binding motifs in the LTR for progesterone to activate the expression of HERVs, the palindromic PRE (GGTGCAAGATGTTCT), and the octamer motif (ATGCAAAG). Only LTR5HS of the HML-2 subgroup of HERV-K contains both elements. 14 Therefore, it is probably the only category of LTRs in the human genome that is responsive to progesterone.
A chain of actions now emerges in the development of at least some breast cancer. Estradiol induces the expression of PR, which, when bound to progesterone, recruits OCT4—which exists in very low levels in tumor-initiating cells 26 —to LTR5HS. OCT4 then activates HERV-K, whose Env product promotes cancer development through the Ras-Raf-MEK-ERK pathway (Fig. 6). Drugs inhibiting multiple links in this chain may work synergistically in the treatment of breast cancer.

PR and OCT4 bind their corresponding motifs in the LTR to activate transcription of Env, which upregulates expression of Ras in several breast cancer cell lines. 8 Since Wnt-1, EGFR, and c-Src are all found to be required for progesterone-mediated ERK activation in T47D cells, 7 Env may also act upstream of Wnt-1. Once activated, ERK1/2 can induce expression of progrowth genes. ERK1/2 also phosphorylates PR, enhancing its ability to induce cell proliferation. 4,5,31 EGFR, epidermal growth factor receptor; ERK, extracellular signal-regulated kinase.
It is noteworthy that both OCT4 and HERVs are expressed in diverse cancer types. 26,27 Therefore the interaction between OCT4 and HERVs during tumorigenesis may be a widespread phenomenon, especially in reproductive tissues, where hormonal influence is prominent, or in germline tissues, where global DNA demethylation occurs. Both OCT4 and HERVs are active during embryogenesis. In addition, some HERVs are activated during the production of induced pluripotent stem cells (iPSCs) using OCT4. 28 Overexpression of HERV-H causes a differentiation defect in iPSCs. These findings in stem cells offer corroborative evidence of the conspiration between OCT4 and HERV in tumorigenesis.
If the PR prefers methylated DNA in vivo, as we have seen in vitro, or mediates different effects when bound to methylated and unmethylated DNA, it will affect expression of ERVs in germline cells and during early embryonic development, when DNA is demethylated and remethylated. This may have to do with the timing of LTR5HS activation during normal embryogenesis. 29 This finding also has implications in hormone physiology and hormone therapies, as DNA demethylation may affect the potency of PR modulators. In addition, oncologists will have to watch for consequences of endocrine dysregulation and HERV activation when demethylating agents are used in chemotherapy.
Female sex hormones affect the expression of at least two host genes, SLC4A8 and IFT172, through solo LTRs. The LTR in the SLC4A8 gene appears to be more responsive to female sex hormones than the LTR in IFT172. Since the two LTRs are 99% homologous and have identical PREs and octamer motifs, the different response to hormones is most likely due to epigenetic environments. This is evidenced by the different expression profiles of the antisense RNAs in various human tissues. 15 For example, the IFT172 antisense RNA is highly expressed in testicular and seminoma tissues, while the SLC4A8 antisense RNA is more abundant in the occipital cerebral cortex and the hypothalamus.
Remarkably, SLC4A8 is downregulated in basal breast cancers and upregulated in most luminal breast cancers. 30 This gene encodes a membrane protein that pumps the bicarbonate anion into the cell, neutralizing acids produced by glycolytic metabolism in hypoxic environments. 30 Therefore, hormonal regulation of SLC4A8 in cancer cells will likely affect the growth of solid tumors. Further research in this aspect of breast cancer development is warranted.
Footnotes
Acknowledgment
The authors thank Miss Olivia Mueller for creating the graphics.
Author Disclosure Statement
The authors declare that there is no conflict of interests regarding the publication of this article.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.