
Abstract
HIV-1 mucosal transmission in genital epithelia occurs through infection of Langerhans cells and subsequent transinfection of CD4+ T cells. We previously reported that the vasodilator neuropeptide calcitonin gene-related peptide (CGRP), secreted upon activation of sensory peripheral neurons that innervate all mucosal epithelia, significantly inhibits transinfection. To investigate the association between CGRP and HIV-1 during infection, we evaluated circulating CGRP levels in HIV-1-infected patients. Plasma was obtained from combination antiretroviral therapy (cART)-naive or cART-treated patients with primary/acute (PHI) or chronic (CHI) HIV-1 infection, as well as from individuals who naturally control HIV-1 infection, namely exposed seronegatives (ESNs), elite controllers (ECs), and long-term nonprogressors (LTNPs). CGRP plasma levels were measured using an enzyme immunoassay. Compared with healthy HIV-1-negative controls, CGRP plasma levels significantly decreased in PHI patients and even further in CHI patients, but remained unchanged in ESNs, ECs, and LTNPs. Moreover, CGRP plasma levels were restored to baseline upon cART in both PHI and CHI. Finally, CGRP plasma levels directly correlated with CD4+ T cell counts and inversely with viral loads. Altogether, CGRP could serve as a novel diagnostic plasma biomarker for progression of HIV-1 infection. Moreover, administration of CGRP to cART-naive HIV-1-infected patients, to compensate for CGRP decline, could help controlling on-going HIV-1 infection.
Sexual transmission of HIV-1 occurs at mucosal epithelia, upon virus invasion of these protective barriers. One of the mechanisms mediating mucosal HIV-1 entry involves targeting of Langerhans cells (LCs), the resident antigen-presenting cells positioned within the epithelial compartment of genital stratified epithelia. 1 For instance, our own studies showed that inner foreskin LCs are permissive to HIV-1 infection, and, in turn, transinfect CD4+ T cells with HIV-1. 2,3 Characterization of the cellular mechanisms that control transinfection, which might provide new strategies to inhibit HIV-1 mucosal entry, remains ill defined.
We recently hypothesized that peripheral sensory pain neurons (i.e., nociceptors), innervating all mucosal epithelia, might play a role during HIV-1 transinfection. Nociceptor activation by noxious stimuli results not only in transmission of pain information to the central nervous system but also local secretion of different neuropeptides, which exert immunomodulatory function. Among these is the potent vasodilator calcitonin gene-related peptide (CGRP) 4 that mediates neurogenic inflammation and modulates immune functions within mucosal tissues. Based on the capacity of CGRP+ nociceptors to directly associate with LCs and of CGRP to modulate LCs function, 5 we showed that CGRP affects several mucosal interactions between LCs and HIV-1, resulting in significant inhibition of HIV-1 transinfection in vitro and ex vivo. 6 –8 These observations demonstrate that mucosal HIV-1 entry could be controlled by a neuroimmune sensory crosstalk. Therefore, we proposed that CGRP could be exploited topically as a microbicide for the clinical prophylactic prevention of mucosal HIV-1 infection.
Circulating CGRP levels vary in different pathological conditions, for example, increase in migraine and decrease in hypertension. 9 Whether CGRP levels are also altered during active HIV-1 infection is elusive.
Hence, to better characterize the association between HIV-1 and circulating CGRP during on-going infection, we measured comparatively the levels of CGRP in the plasma of different groups of HIV-1-infected patients, using the alpha-CGRP (human) Enzyme Immunoassay kit (Peninsula Laboratories, San Carlos, CA), according to the manufacturer's instructions. The studied populations were recruited at the Department of Infectious Diseases of the San Raffaele Scientific Institute, upon approval of the institutional review board and the local ethic committee (MUCIM protocol, no. 95/DG, January 2007), and all subjects provided informed consent for the study.
A total of 104 plasma samples were tested, as follows: (1) HIV-1 exposed seronegatives (ESNs 10 ; n = 9) that we previously described, 11 including individuals remaining seronegative despite a prolonged history of unprotected sexual intercourses with HIV-1 seropositive partners at least twice per week and for at least 2 years; (2) elite controllers (ECs 12 ; n = 7), defined as HIV-1-infected patients able to exert spontaneous control of viremia for at least 2 consecutive years in the absence of combination antiretroviral therapy (cART) and viral loads persistently <37 copies/mL with certified seroconversion for ≥7 years; (3) long-term nonprogressors (LTNPs; n = 7) that were part of the Evaluation of Long-term nonprogressors' Viroimmunology Study cohort and described in our previous study. 13 The inclusion criteria for the LTNPs cohort were (i) certified HIV-1 seroconversion for ≥7 years; (ii) CD4 T cell counts ≥500 cells/μl; (iii) cART-naive; (iv) asymptomatic HIV-1 infection; (v) good health condition; (4) HIV-1-infected patients with primary/acute infection (PHI), defined on the basis of the following criteria 14 : (i) presence of an acute retroviral syndrome; (ii) positive test for HIV-1 RNA in plasma; (iii) presence of less than three positive bands in a Western blot assay. For cART-naive PHI patients (n = 29), samples were withdrawn during the first month after the first HIV-1-positive test. A subset of PHI patients (n = 9) was offered cART (i.e., two or three nucleoside reverse transcriptase inhibitors and one or two protease inhibitors) at diagnosis, and samples were collected at disease onset and 2–8 months after cART initiation; (5) HIV-1-infected patients with chronic HIV-1 infection (CHI) for at least 8 months and cART-naive (n = 19). An additional group included CHI patients under cART (defined above) for at least 24 and not more than 30 months, with chronic and progressive infection but without previous AIDS-defining disease (n = 10). A similar length of time of suppressive therapy in cART-treated CHI patients was chosen to minimize possible differences in immune status. Age-matched healthy individuals not exposed to HIV-1 (n = 23) served as control. All plasma samples were heat inactivated before testing.
Compared with those in healthy HIV-1-negative controls, CGRP plasma levels were significantly decreased in PHI patients and even further in CHI patients (Fig. 1A; means [95% confidence interval (CIs)] of 43.2 [31.0–55.5], 28.5 [19.8–37.1], and 8.4 [2.9–14.0], for controls, PHI and CHI, respectively). In contrast, no significant differences were observed when comparing controls with either ESNs or ECs/LTNPs (Fig. 1A). These results show that CGRP plasma levels gradually decline as HIV-1 infection proceeds, but remain unchanged when infection is naturally suppressed.
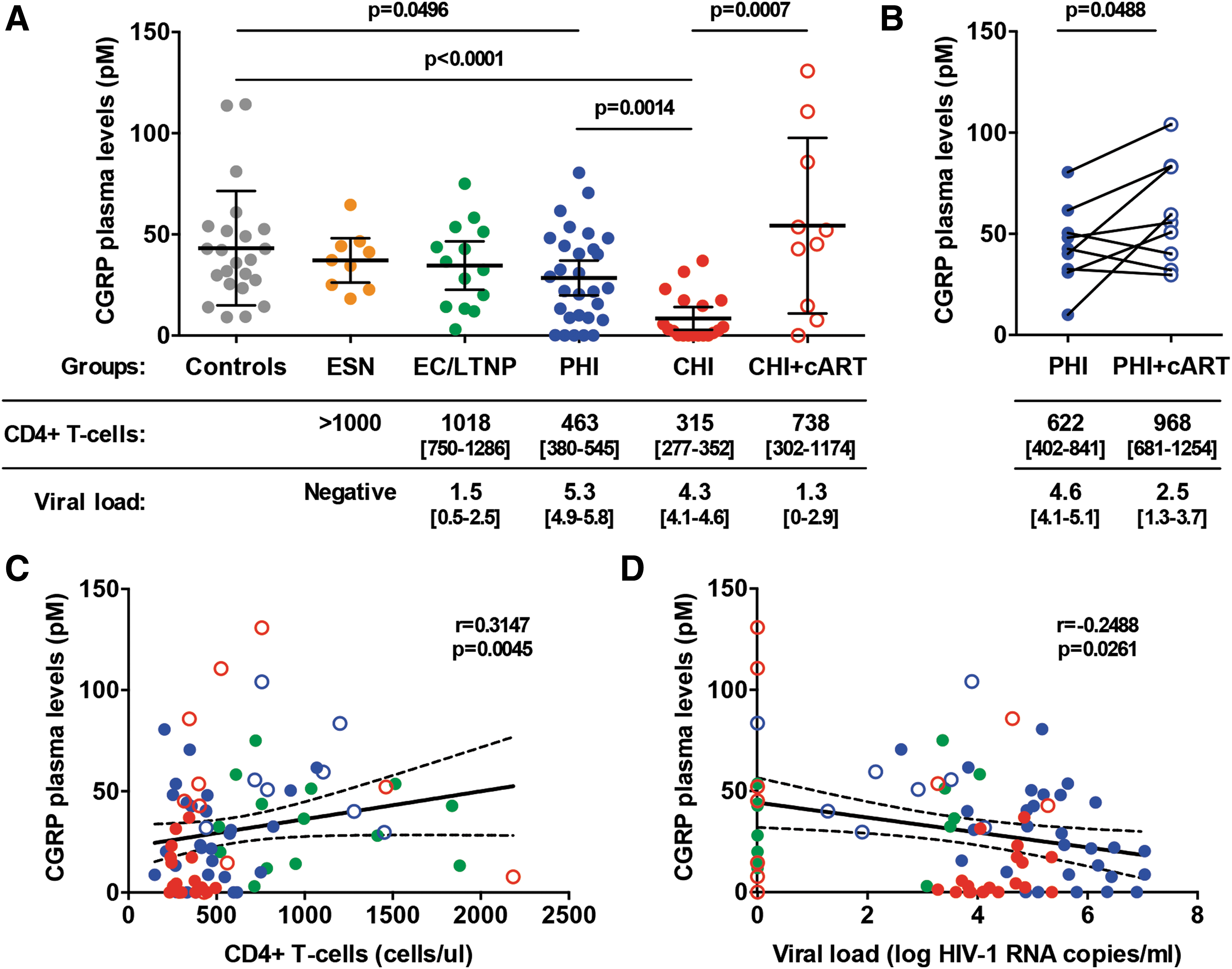
CGRP is a plasma biomarker for HIV-1 disease progression.
Next, we investigated whether cART affects CGRP, by comparing CGRP plasma levels in HIV-1-infected patients who were either cART-naive or cART-treated. Although highly variable among cART-treated CHI patients, the overall mean of CGRP plasma levels was significantly increased upon treatment, when compared with that in cART-naive CHI patients, and was normalized to that in the control group (Fig. 1A; mean [95% CIs] of 54.3 [23.3–85.4] for CHI+cART). For some PHI patients (n = 9), we could measure CGRP in plasma samples from the same patients, obtained before cART and at follow-up after cART initiation. This analysis revealed that CGRP plasma levels were markedly increased upon cART in six of nine patients (mean fold of 2.2) and slightly decreased in three of nine patients (mean fold of 0.8), with an overall significant increase in mean CGRP levels (Fig. 1B; means [95% CIs] of 44.1 [28.8–59.4] and 59.8 [40.1–79.5] before and after cART, respectively).
Of note, as cART duration was similar in either PHI or CHI patients, the lack of CGRP normalization upon cART in some patients cannot be attributed to differences in treatment duration. These results show that CGRP plasma levels are restored when HIV-1 replication is suppressed upon cART. Normalization of CGRP plasma levels probably occurs through a cART-indirect mechanism related to restoration of the immunological status in HIV-1-infected patients.
Finally, we determined whether CGRP plasma levels correlate with clinical parameters of HIV-1 disease progression evaluated in plasma, namely CD4+ T cell counts and viral loads. As expected, CD4+ T cell counts significantly and inversely correlated with viral loads (Spearman r = −0.5457, p < .0001), when considering the data from all ECs/LTNPs, PHI ± cART, and CHI ± cART patients. In all of these patients taken together, CGRP plasma levels directly correlated with CD4+ T cell counts (Fig. 1C) and inversely with viral loads (Fig. 1D). These correlations were significant but weak (Spearman r = 0.3147 and −0.2488, respectively), probably due to the low number of patients tested in each group. These results show that the decline in CGRP plasma levels is significantly associated with other clinical parameters of HIV-1 progression.
Our results show that CGRP plasma levels progressively decline in uncontrolled HIV-1 infection, but remain or normalize to baseline levels upon natural HIV-1 control or cART. We, therefore, suggest that decreased levels of CGRP, which directly correlate with CD4+ T cell counts and inversely with viral loads, could provide a novel diagnostic plasma biomarker for active HIV-1 disease progression. As CGRP has protective roles during cardiovascular pathophysiology, 15 –17 we suggest that its decline might contribute to the higher incidence of such complications in HIV-1-infected patients. 18,19 Moreover, we propose that supplementing CGRP in cART-naive HIV-1-infected patients, to counteract the overall CGRP decline, might prevent on-going HIV-1 transinfection, for example, once HIV-1-positive LCs have migrated to secondary lymphoid organs to disseminate the infection and facilitate disease progression.
Measuring CGRP tissue levels is challenging due to difficulties in obtaining mucosal tissues from HIV-1-infected patients. Therefore, as a surrogate of mucosal CGRP, we evaluated here the levels of circulating CGRP in blood. Plasma CGRP is generally attributed to spillover into blood from tissue sites of neuronal release, 9 with nociceptors being the primary source of secreted CGRP.
Reduced levels of CGRP could hence result from HIV-1-mediated increase in CGRP-degrading enzymes 9 and/or factors that inhibit CGRP secretion from nociceptors. 20 In parallel, as HIV-1 is neurotoxic to peripheral neurons and leads to neuronal apoptosis, 21 one could speculate that uncontrolled HIV-1 infection induces nociceptor loss and subsequently decreased CGRP secretion, which is reflected in plasma. Yet, HIV-1-induced neuronal damage persists under cART 22 and, therefore, cannot exclusively explain the normalization in CGRP plasma levels upon cART. Although non-neuronal cells also secret CGRP (i.e., endothelial cells, epithelial cells, and different immune cells 9 ), their contribution to CGRP release into blood remains unknown. Future studies are now needed to determine the cellular sources of CGRP in cART-treated patients, for instance, circulating T cells that secret CGRP, 9 whose counts were found herein to directly correlate with CGRP plasma levels.
Taken together, we propose that in addition to the potential topical use of CGRP to limit infection during sexual transmission, 8 supplementing CGRP clinically in cART-naive HIV-1-infected patients could also limit on-going HIV-1 infection.
Footnotes
Acknowledgments
This study was supported by grants to M.B. from the French National Agency for AIDS and Hepatitis research (Agence Nationale de Recherche sur le SIDA et les Hépatites, ANRS), and to Y.G. from SIDACTION.
Author Disclosure Statement
The authors declare no conflict of interest.